
Are Chilean Pasture Seed End-Users Adopting New Species? Trends and Joinpoint Regression Analysis of the Last 19 Years of Seed Imports


Abstract
1. Introduction
2. Materials and Methods
2.1. Data Source, Criteria Selection and Classification
2.2. Statistical Analysis
3. Results
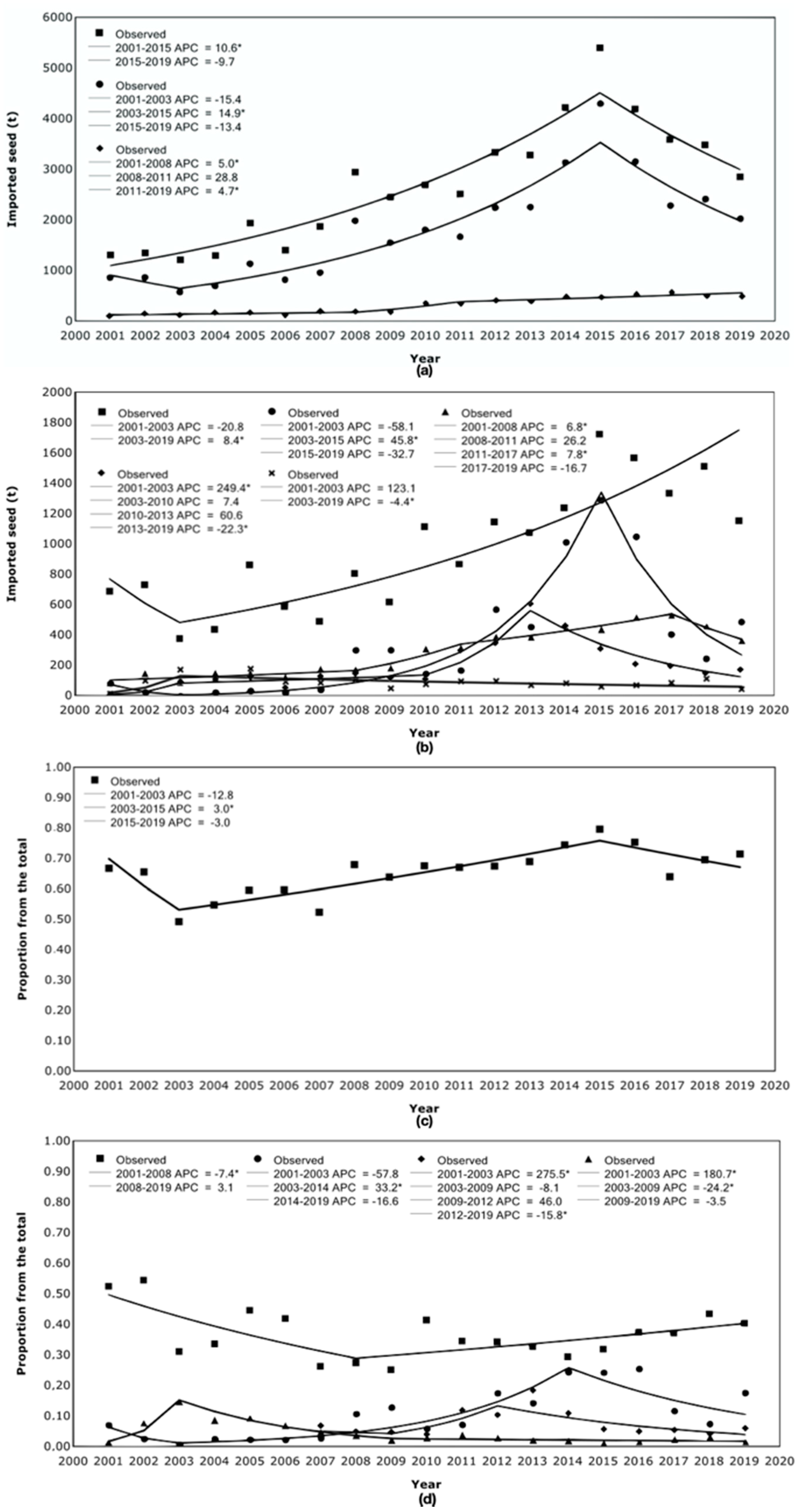
3.1. Total Imported Seed
3.2. Genus and Group Trends and Changes
3.3. Species Trends and Changes
4. Discussion
4.1. Overview of the Genus and Species Imported
4.2. Chances of Adopting New Species
4.3. Implications for the Agricultural Policies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oenema, O.; de Klein, C.; Alfaro, M. Intensification of grassland and forage use: Driving forces and constraints. Crop Pasture Sci. 2014, 65, 524–537. [Google Scholar] [CrossRef]
- Herrero, M.; Havlik, P.; Valin, H.; Notenbaert, A.; Rufino, M.C.; Thornton, P.K.; Blummel, M.; Weiss, F.; Grace, D.; Obersteiner, M. Biomass use, production, feed efficiencies, and greenhouse gas emissions from global livestock systems. Proc. Natl. Acad. Sci. USA 2013, 110, 20888–20893. [Google Scholar] [CrossRef]
- Wilkins, P.W.; Humphreys, M.O. Progress in breeding perennial forage grasses for temperate agriculture. J. Agric. Sci. 2003, 140, 129–150. [Google Scholar] [CrossRef]
- Duller, S.; Thorogood, D.; Bonos, S.A. Breeding Objectives in Amenity Grasses. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 137–160. [Google Scholar]
- Beard, J.B.; Green, R.L. The Role of Turfgrasses in Environmental Protection and Their Benefits to Humans. J. Environ. Qual. 1994, 23, 452–460. [Google Scholar] [CrossRef]
- Bondesen, O. Seed production and seed trade in a globalised world. Seed production in the northern light. In Proceedings of the 6th International Herbage Seed Conference, Gjennestad, Norway, 18–20 June 2007; pp. 9–12. [Google Scholar]
- ISF Forage & Turf Crop Seed Statistics. Available online: http://www.worldseed.org/cms/medias/file/ResourceCenter/SeedStatistics/ForageandTurfSeedMarket/Seed_Production_of_Selected_Species_2006.pdf (accessed on 15 June 2016).
- Huyghe, C.; De Vliegher, A.; Van Gils, B.; Peeters, A. Grasslands in the economic supply chain: Related industries. In Grasslands and Herbivore Production in Europe and Effects of Common Policies; Huyghe, C., De Vliegher, A., Van Gils, B., Peeters, A., Eds.; éditions Quae: Versailles, France, 2014; pp. 227–238. ISBN 9782759221578. [Google Scholar]
- Chapman, D.F.; Bryant, J.R.; Olayemi, M.E.; Edwards, G.R.; Thorrold, B.S.; McMillan, W.H.; Kerr, G.A.; Judson, G.; Cookson, T.; Moorhead, A.; et al. An economically based evaluation index for perennial and short-term ryegrasses in New Zealand dairy farm systems. Grass Forage Sci. 2017, 72, 1–21. [Google Scholar] [CrossRef]
- McDonagh, J.; O’Donovan, M.; McEvoy, M.; Gilliland, T.J. Genetic gain in perennial ryegrass (Lolium perenne) varieties 1973 to 2013. Euphytica 2016, 212, 187–199. [Google Scholar] [CrossRef]
- Rolston, M.P.; Chynoweth, R.J.; Vreugdenhil, S.R.; Gunnarsson, A.M. Ryegrass seed production in New Zealand: Achieving 3000+ kg/ha yields. Agron. N. Z. 2018, 48, 115–123. [Google Scholar]
- Grogan, D.; Gilliland, T.J. A review of perennial ryegrass variety evaluation in Ireland. Ir. J. Agric. Food Res. 2011, 50, 65–81. [Google Scholar]
- Gilliland, T.J.; Johnston, J.; Connolly, C. A review of forage grass and clover seed use in Northern Ireland, UK between 1980 and 2004. Grass Forage Sci. 2007, 62, 239–254. [Google Scholar] [CrossRef]
- Hannaway, D.B.; Daly, C.; Halbleib, M.D.; James, D.; West, C.P.; Volenec, J.F.; Chapman, D.; Li, X.; Cao, W.; Shen, J.; et al. Development of suitability maps with examples for the United States and China. In Tall Fescue for the 21st Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; ASA, CSSA, SSSA: Madison, WI, USA, 2009; pp. 33–47. [Google Scholar]
- Annicchiarico, P.; Barrett, B.; Brummer, E.C.; Julier, B.; Marshall, A.H. Achievements and Challenges in Improving Temperate Perennial Forage Legumes. Crit. Rev. Plant Sci. 2015, 34, 327–380. [Google Scholar] [CrossRef]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grassland-livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.; Inostroza, L.; Moscoso, C.; Parra, L.; Quiroz, A. Persistence and yield stability of temperate grassland legumes for sustainable animal production. In Improving Grassland and Pasture Management in Temperate Agriculture; Marshall, A.H., Collins, R., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018; pp. 219–236. ISBN 9781786762009. [Google Scholar]
- Boelt, B.; Julier, B.; Karagić, Đ.; Hampton, J. Legume Seed Production Meeting Market Requirements and Economic Impacts. Crit. Rev. Plant Sci. 2015, 34, 412–427. [Google Scholar] [CrossRef]
- Moscoso, C.J.; Urrutia, N.L. Overview of the Forage Land-Use in Southern Chile in a Thirty-Year Period. Crop. Forage Turfgrass Manag. 2017, 3, 1–4. [Google Scholar] [CrossRef]
- Demanet, R.; Mora, M.L.; Cussen, C.R.; Rowarth, J.S. Seed Production in Chile; International Herbage Seed Production Research Group: Auckland, New Zealand, 2000; pp. 3–6. [Google Scholar]
- Acuña, A.; Moscoso, C.J.; Danneberger, K.; Muñoz-Araya, M. Evolution of Chile’s Turfgrass Seed Market (2001–2015). Int. Turfgrass Soc. Res. J. 2017, 13, 477–479. [Google Scholar] [CrossRef]
- Stolpe, N.; Undurraga, P. Long term climatic trends in Chile and effects on soil moisture and temperature regimes. Chil. J. Agric. Res. 2016, 76, 487–496. [Google Scholar] [CrossRef]
- Araya-Osses, D.; Casanueva, A.; Román-Figueroa, C.; Uribe, J.M.; Paneque, M. Climate change projections of temperature and precipitation in Chile based on statistical downscaling. Clim. Dyn. 2020, 54, 4309–4330. [Google Scholar] [CrossRef]
- Pembleton, K.G.; Tozer, K.N.; Edwards, G.R.; Jacobs, J.L.; Turner, L.R. Simple versus diverse pastures: Opportunities and challenges in dairy systems. Anim. Prod. Sci. 2015, 55, 893–901. [Google Scholar] [CrossRef]
- Ergon, Å.; Seddaiu, G.; Korhonen, P.; Virkajärvi, P.; Bellocchi, G.; Jørgensen, M.; Østrem, L.; Reheul, D.; Volaire, F. How can forage production in Nordic and Mediterranean Europe adapt to the challenges and opportunities arising from climate change? Eur. J. Agron. 2018, 92, 97–106. [Google Scholar] [CrossRef]
- Gresta, F.; Cristaudo, A.; Onofri, A.; Restuccia, A.; Avola, G. Germination response of four pasture species to temperature, light, and post-harvest period. Plant Biosyst. 2010, 144, 849–856. [Google Scholar] [CrossRef]
- Moot, D.J.; Scott, W.R.; Roy, A.M.; Nicholls, A.C. Base temperature and thermal time requirements for germination and emergence of temperate pasture species. N. Z. J. Agric. Res. 2000, 43, 15–25. [Google Scholar] [CrossRef]
- Humphreys, M.; Feuerstein, U.; Vandewalle, M.; Baert, J. Ryegrasses. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 211–260. [Google Scholar]
- Rognli, O.A.; Saha, M.C.; Bhamidimarri, S.; van der Heijden, S. Fescues. In Fodder Crops and Amenity Grasses; Springer: New York, NY, USA, 2010; pp. 261–292. [Google Scholar]
- Hamed, K.H.; Rao, A.R. A modified Mann-Kendall trend test for autocorrelated data. J. Hydrol. 1998, 204, 182–196. [Google Scholar] [CrossRef]
- Kim, H.J.; Fay, M.P.; Feuer, E.J.; Midthune, D.N. Permutation tests for joinpoint regression with applications to cancer rates. Stat. Med. 2000, 19, 335–351. [Google Scholar] [CrossRef]
- Iarocci, G.; Cocchiara, R.A.; Sestili, C.; Del Cimmuto, A.; La Torre, G. Variation of atmospheric emissions within the road transport sector in Italy between 1990 and 2016. Sci. Total Environ. 2019, 692, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Agaku, I.T.; Alpert, H.R. Trends in annual sales and current use of cigarettes, cigars, roll-your-own tobacco, pipes, and smokeless tobacco among US adults, 2002–2012. Tob. Control 2016, 25, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Gillis, D.; Edwards, B.P.M. The utility of joinpoint regression for estimating population parameters given changes in population structure. Heliyon 2019, 5, e02515. [Google Scholar] [CrossRef] [PubMed]
- Cash-Gibson, L.; Rojas-Gualdrón, D.F.; Pericàs, J.M.; Benach, J. Inequalities in global health inequalities research: A 50-year bibliometric analysis (1966–2015). PLoS ONE 2018, 13, e0191901. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-S.; Mariotto, A.B.; Zhu, L.; Kim, H.-J.; Cho, H.; Feuer, E.J. Developments and challenges in statistical methods in cancer surveillance. Stat. Interface 2014, 7, 135–151. [Google Scholar] [CrossRef]
- NCI Joinpoint Regression Program, Version 4 8.0.1; National Cancer Institute: Bethesda, MD, USA, 2020.
- Xu, J.; Lin, Y.; Yang, M.; Zhang, L. Statistics and pitfalls of trend analysis in cancer research: A review focused on statistical packages. J. Cancer 2020, 11, 2957–2961. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- ODEPA. Estadisticas Económicas. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/estadisticas-economicas (accessed on 6 October 2020).
- del Pozo, A.; Ovalle, C.; Espinoza, S.; Barahona, V.; Gerding, M.; Humphries, A. Water relations and use-efficiency, plant survival and productivity of nine alfalfa (Medicago sativa L.) cultivars in dryland Mediterranean conditions. Eur. J. Agron. 2017, 84, 16–22. [Google Scholar] [CrossRef]
- Ovalle, C.; Espinoza, S.; Barahona, V.; Gerding, M.; Humphries, A.; del Pozo, A. Lucerne and other perennial legumes provide new options for rain fed livestock production in the Mediterranean-climate region of Chile. Cienc. Investig. Agrar. 2015, 42, 453–460. [Google Scholar] [CrossRef]
- Neal, J.S.; Fulkerson, W.J.; Lawrie, R.; Barchia, I.M. Difference in yield and persistence among perennial forages used by the dairy industry under optimum and deficit irrigation. Crop Pasture Sci. 2009, 60, 1071–1087. [Google Scholar] [CrossRef]
- Burnett, V.F.; Butler, K.L.; Hirth, J.R.; Mitchell, M.L.; Clark, S.G.; Nie, Z. Lucerne (Medicago sativa L.) Persistence Remains Unchanged under Variable Cutting Regimes. Agronomy 2020, 10, 844. [Google Scholar] [CrossRef]
- Ovalle, C.; del Pozo, A.; Avendaño, J.; Aravena, T.; Díaz, M.E. Cauquenes-INIA, Nuevo cultivar de Hualputra Chilena (Medicago polymorpha) para Areas de Secano Mediterraneo. Agric. Técnica 2001, 61, 89–92. [Google Scholar] [CrossRef]
- Peyraud, J.L.; Le Gall, A.; Lüscher, A. Potential food production from forage legume-based-systems in Europe: An overview. Ir. J. Agric. Food Res. 2009, 48, 115–135. [Google Scholar]
- Chapman, D.F.; Crush, J.R.; Lee, J.M.; Cosgrove, G.P.; Stevens, D.R.; Rossi, L.; Popay, A.J.; Edwards, G.R.; King, W.M. Implications of grass–clover interactions in dairy pastures for forage value indexing systems. 6. Cross-site analysis and general discussion. N. Z. J. Agric. Res. 2018, 61, 255–284. [Google Scholar] [CrossRef]
- Dineen, M.; Delaby, L.; Gilliland, T.; McCarthy, B. Meta-analysis of the effect of white clover inclusion in perennial ryegrass swards on milk production. J. Dairy Sci. 2018, 101, 1804–1816. [Google Scholar] [CrossRef] [PubMed]
- Weggler, K.; Thumm, U.; Elsaesser, M. Development of Legumes After Reseeding in Permanent Grassland, as Affected by Nitrogen Fertilizer Applications. Agriculture 2019, 9, 207. [Google Scholar] [CrossRef]
- Ovalle, C.; del Pozo, A.; Fernández, F.; Chavarría, J.; Arredondo, S. Arrowleaf Clover (Trifolium vesiculosum Savi): A New Species of Annual Legumes for High Rainfall Areas of the Mediterranean Climate Zone of Chile. Chil. J. Agric. Res. 2010, 70, 170–177. [Google Scholar] [CrossRef]
- Porqueddu, C.; Ates, S.; Louhaichi, M.; Kyriazopoulos, A.P.; Moreno, G.; del Pozo, A.; Ovalle, C.; Ewing, M.A.; Nichols, P.G.H. Grasslands in ‘Old World’ and ‘New World’ Mediterranean-climate zones: Past trends, current status and future research priorities. Grass Forage Sci. 2016, 71, 1–35. [Google Scholar] [CrossRef]
- Nichols, P.G.H.; Revell, C.K.; Humphries, A.W.; Howie, J.H.; Hall, E.J.; Sandral, G.A.; Ghamkhar, K.; Harris, C.A. Temperate pasture legumes in Australia—their history, current use, and future prospects. Crop Pasture Sci. 2012, 63, 691–725. [Google Scholar] [CrossRef]
- Enriquez-Hidalgo, D.; Cruz, T.; Teixeira, D.L.; Steinfort, U. Phenological stages of Mediterranean forage legumes, based on the BBCH scale. Ann. Appl. Biol. 2020, 176, 357–368. [Google Scholar] [CrossRef]
- Ortega, F.; Parra, L.; Quiroz, A. Breeding red clover for improved persistence in Chile: A review. Crop Pasture Sci. 2014, 65, 1138–1146. [Google Scholar] [CrossRef]
- Stewart, A.V.; Ellison, N.W. A molecular phylogenetic framework for cocksfoot (Dactylis glomerata L.) improvement. Crop Pasture Sci. 2014, 65, 780–786. [Google Scholar] [CrossRef]
- Ghesquière, M.; Humphreys, M.W.; Zwierzykowski, Z. Festulolium. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 293–316. [Google Scholar]
- Lee, J.M.; Hemmingson, N.R.; Minnee, E.M.K.; Clark, C.E.F. Management strategies for chicory (Cichorium intybus) and plantain (Plantago lanceolata): Impact on dry matter yield, nutritive characteristics and plant density. Crop Pasture Sci. 2015, 66, 168–183. [Google Scholar] [CrossRef]
- Langworthy, A.D.; Rawnsley, R.P.; Freeman, M.J.; Pembleton, K.G.; Corkrey, R.; Harrison, M.T.; Lane, P.A.; Henry, D.A. Potential of summer-active temperate (C3) perennial forages to mitigate the detrimental effects of supraoptimal temperatures on summer home-grown feed production in south-eastern Australian dairying regions. Crop Pasture Sci. 2018, 69, 808–820. [Google Scholar] [CrossRef]
- Tozer, K.; Barker, G.; Cameron, C.; Wilson, D.; Loick, N. Effects of including forage herbs in grass–legume mixtures on persistence of intensively managed pastures sampled across three age categories and five regions. N. Z. J. Agric. Res. 2016, 59, 250–268. [Google Scholar] [CrossRef]
- McCarthy, K.M.; McAloon, C.G.; Lynch, M.B.; Pierce, K.M.; Mulligan, F.J. Herb species inclusion in grazing swards for dairy cows—A systematic review and meta-analysis. J. Dairy Sci. 2020, 103, 1416–1430. [Google Scholar] [CrossRef]
- Perera, R.S.; Cullen, B.R.; Eckard, R.J. Growth and Physiological Responses of Temperate Pasture Species to Consecutive Heat and Drought Stresses. Plants 2019, 8, 227. [Google Scholar] [CrossRef]
- Gilliland, T.J. Control of Cultivar Release and Distribution. In Fodder Crops and Amenity Grasses; Boller, B., Posselt, U.K., Veronesi, F., Eds.; Springer: New York, NY, USA, 2010; pp. 175–199. [Google Scholar]
- Hampton, J.; Conner, A.; Boelt, B.; Chastain, T.; Rolston, P. Climate Change: Seed Production and Options for Adaptation. Agriculture 2016, 6, 33. [Google Scholar] [CrossRef]
- Ortega, F.; Moscoso, C.; Levío, J. Respuesta a la fertilización nitrogeneda de cultivares chilenos de bromo (Bromus valdivianus Phil) y de bromo Gala. In Proceedings of the del XXXVIII Congreso de la Sociedad Chilena de Producción Animal, Frutillar, Chile, 23–25 October 2013; Sociedad Chilena de Producción Animal: Frutillar, Chile, 2013; pp. 135–136. [Google Scholar]
- López-Olivari, R.; Ortega-Klose, F. Response of red clover to deficit irrigation: Dry matter yield, populations, and irrigation water use efficiency in southern Chile. Irrig. Sci. 2021, 39, 173–189. [Google Scholar] [CrossRef]
- Rogers, M.-J.; Lawson, A.; Kelly, K. Forage Options for Dairy Farms with Reduced Water Availability in the Southern Murray Darling Basin of Australia. Sustainability 2017, 9, 2369. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Metric tons (Mt) | |||||||||||||||||||
| Lolium spp. 1 | 872 | 879 | 594 | 709 | 1148 | 836 | 975 | 1994 | 1563 | 1816 | 1680 | 2249 | 2263 | 3135 | 4293 | 3152 | 2291 | 2417 | 2033 |
| Festuca spp. 1 | 109 | 157 | 128 | 179 | 178 | 128 | 203 | 197 | 191 | 360 | 356 | 417 | 397 | 489 | 480 | 540 | 577 | 504 | 496 |
| Medicago spp. | 170 | 51 | 133 | 162 | 234 | 220 | 334 | 388 | 468 | 153 | 118 | 300 | 365 | 253 | 221 | 159 | 275 | 261 | 145 |
| Trifolium spp. | 124 | 187 | 240 | 158 | 253 | 154 | 177 | 200 | 147 | 153 | 183 | 174 | 128 | 152 | 146 | 105 | 149 | 167 | 60 |
| Other pastures 2 | 33 | 67 | 111 | 73 | 104 | 64 | 153 | 156 | 81 | 178 | 153 | 177 | 112 | 185 | 194 | 167 | 293 | 108 | 114 |
| Not determined 3 | 1 | 1 | 16 | 14 | 22 | 3 | 30 | 18 | 20 | 20 | 59 | 63 | 21 | ||||||
| Total | 1308 | 1342 | 1207 | 1296 | 1930 | 1402 | 1865 | 2937 | 2450 | 2690 | 2507 | 3337 | 3284 | 4214 | 5393 | 4186 | 3585 | 3478 | 2848 |
| Metric tons (Mt) | Proportion of the Total (PT) | |||||||
|---|---|---|---|---|---|---|---|---|
| Mann–Kendall | AAPC 3 | Joinpoints | Mann–Kendall | AAPC 3 | Joinpoints | |||
| Group/Species | p-Value | Trend | % | n | p-Value | Trend | % | n |
| Total | <0.001 | Increasing 2 | 5.7 4 | 2 | ||||
| Lolium spp. | <0.001 | Increasing 2 | 4.3 | 3 | 0.001 | Increasing | −0.2 | 3 |
| L. perenne | <0.001 | Increasing 2 | 4.7 | 2 | 0.726 | No trend | −1.1 | 2 |
| L. multiflorum | <0.001 | Increasing 2 | 6.9 | 3 | <0.001 | Increasing | 2.9 | 3 |
| L. x hybridum | 0.001 | Increasing 2 | 17.2 | 3 | 0.363 | No trend | 11.8 | 3 |
| Not determined 1 | 0.007 | Increasing | 7.9 4 | 0 | 0.833 | No trend | 0.4 | 0 |
| Festuca spp. | <0.001 | Increasing 2 | 4.3 | 3 | 0.025 | Increasing | 2.4 4 | 0 |
| F. arundinacea | <0.001 | Increasing 2 | 7.1 | 3 | 0.003 | Increasing | 2.6 4 | 0 |
| Not determined 1 | 0.024 | Increasing | 8.7 4 | 0 | 0.994 | No trend | 1.3 | 0 |
| Medicago spp. | 0.441 | No trend | 2.6 | 0 | 0.074 | No trend | −4.4 4 | 0 |
| M. polymorpha | 0.242 | No trend | 0.431 | No trend | ||||
| M. sativa | 0.441 | No trend | 2.6 | 0 | 0.050 | Decreasing | −6.1 | 0 |
| Trifolium spp. | 0.021 | Decreasing | −2.9 4 | 0 | <0.001 | Decreasing | −9.6 4 | 0 |
| T. repens | 0.150 | No trend | 5.1 | 2 | 0.003 | Decreasing | −1.3 | 3 |
| T. subterraneum | 0.383 | No trend | 5.1 | 0 | 0.001 | Decreasing | −8.5 4 | 0 |
| T. alexandrinum | 0.061 | No trend | −15.6 4 | 0 | 0.001 | Decreasing | −22.5 4 | 0 |
| T. pratense | 0.456 | No trend | 7.4 | 0 | 0.901 | No trend | 0.1 | 0 |
| T. fragiferum | 0.029 | Decreasing | −9.6 | 0 | 0.001 | Decreasing | −16.6 | 0 |
| T. michelianum | 0.837 | No trend | −4.8 | 0 | 0.372 | No trend | −14.4 | 0 |
| T. vesiculosum | 0.360 | No trend | 0.592 | No trend | ||||
| Not determined 1 | 0.276 | No trend | 2.1 | 0 | 0.154 | No trend | ||
| Other pastures | 0.003 | Increasing | 6.1 4 | 0 | 0.233 | No trend | −1.2 | 0 |
| D. glomerata | 0.009 | Increasing | 6.6 4 | 0 | 0.441 | No trend | −1.3 | 0 |
| x Festulolium | 0.392 | No trend | 4.9 | 0 | 0.752 | No trend | −2.7 | 0 |
| Phalaris spp. | 0.011 | Decreasing | −12.9 4 | 0 | 0.001 | Decreasing | −21.8 4 | 0 |
| P. lanceolata | 0.096 | No trend | 17.4 4 | 0 | 0.372 | No trend | 7.2 | 0 |
| C. intybus | 0.064 | No trend | 63.2 4 | 0 | 0.046 | Increasing | 24.1 | 0 |
| Lotus spp. | 0.386 | No trend | 0.500 | No trend | ||||
| Bromus spp. | 0.080 | No trend | 0.386 | No trend | ||||
| Not determined 1 | 0.212 | No trend | 27.7 4 | 0 | 0.996 | No trend | −0.4 | 0 |
| Group/Species | 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | 2010 | 2011 | 2012 | 2013 | 2014 | 2015 | 2016 | 2017 | 2018 | 2019 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lolium spp. | |||||||||||||||||||
| L. perenne | 0.525 | 0.544 | 0.310 | 0.336 | 0.446 | 0.418 | 0.263 | 0.274 | 0.251 | 0.414 | 0.346 | 0.343 | 0.327 | 0.294 | 0.319 | 0.374 | 0.372 | 0.435 | 0.404 |
| L. multiflorum | 0.069 | 0.024 | 0.005 | 0.024 | 0.021 | 0.020 | 0.026 | 0.105 | 0.126 | 0.057 | 0.070 | 0.172 | 0.140 | 0.240 | 0.239 | 0.251 | 0.114 | 0.072 | 0.173 |
| L. x hybridum | 0.007 | 0.012 | 0.075 | 0.081 | 0.062 | 0.036 | 0.067 | 0.049 | 0.047 | 0.040 | 0.117 | 0.102 | 0.182 | 0.108 | 0.057 | 0.049 | 0.054 | 0.042 | 0.059 |
| L. rigidum | 0.006 | 0.008 | |||||||||||||||||
| Not determined 1 | 0.067 | 0.075 | 0.101 | 0.101 | 0.057 | 0.121 | 0.167 | 0.252 | 0.214 | 0.165 | 0.137 | 0.057 | 0.041 | 0.102 | 0.181 | 0.079 | 0.099 | 0.147 | 0.078 |
| Festuca spp. | |||||||||||||||||||
| F. arundinacea | 0.068 | 0.113 | 0.076 | 0.118 | 0.079 | 0.090 | 0.097 | 0.060 | 0.076 | 0.116 | 0.127 | 0.118 | 0.119 | 0.107 | 0.082 | 0.124 | 0.149 | 0.133 | 0.129 |
| Not determined 1 | 0.015 | 0.004 | 0.030 | 0.020 | 0.013 | 0.001 | 0.012 | 0.006 | 0.002 | 0.018 | 0.015 | 0.007 | 0.002 | 0.010 | 0.008 | 0.005 | 0.012 | 0.012 | 0.044 |
| Medicago spp. | |||||||||||||||||||
| M. polymorpha | 0.001 | 0.002 | 0.003 | <0.001 | 0.002 | 0.002 | 0.002 | 0.002 | <0.001 | 0.002 | |||||||||
| M. sativa | 0.130 | 0.038 | 0.110 | 0.125 | 0.121 | 0.156 | 0.177 | 0.130 | 0.191 | 0.056 | 0.045 | 0.088 | 0.109 | 0.058 | 0.041 | 0.038 | 0.076 | 0.072 | 0.051 |
| M. truncatula | <0.001 | <0.001 | 0.001 | ||||||||||||||||
| Trifolium spp. | |||||||||||||||||||
| T. repens | 0.016 | 0.079 | 0.148 | 0.087 | 0.096 | 0.071 | 0.050 | 0.038 | 0.023 | 0.031 | 0.041 | 0.031 | 0.024 | 0.021 | 0.012 | 0.018 | 0.026 | 0.034 | 0.018 |
| T. subterraneum | 0.037 | 0.016 | 0.025 | 0.026 | 0.018 | 0.028 | 0.010 | 0.009 | 0.012 | 0.004 | 0.015 | 0.010 | 0.008 | 0.001 | 0.003 | 0.014 | 0.009 | 0.003 | |
| T. alexandrinum | 0.022 | 0.011 | 0.033 | 0.018 | 0.007 | 0.008 | 0.006 | 0.001 | <0.001 | 0.002 | |||||||||
| T. pratense | <0.001 | 0.002 | 0.002 | 0.001 | <0.001 | <0.001 | 0.008 | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 0.013 | 0.002 | <0.001 | 0.001 | <0.001 | ||
| T. fragiferum | 0.004 | 0.003 | 0.003 | 0.003 | 0.001 | 0.001 | 0.009 | 0.001 | <0.001 | 0.002 | 0.001 | <0.001 | <0.001 | <0.001 | 0.001 | ||||
| T. michelianum | 0.001 | 0.001 | 0.003 | 0.002 | 0.004 | 0.003 | 0.001 | 0.001 | 0.001 | <0.001 | 0.001 | 0.001 | |||||||
| T. vesiculosum | <0.001 | 0.001 | <0.001 | 0.001 | 0.001 | <0.001 | |||||||||||||
| T. incarnatum | <0.001 | <0.001 | |||||||||||||||||
| T. ambiguum | <0.001 | <0.001 | |||||||||||||||||
| Not determined 1 | 0.054 | 0.001 | <0.001 | 0.007 | 0.002 | 0.013 | 0.007 | 0.003 | 0.004 | 0.028 | 0.003 | 0.002 | 0.004 | 0.001 | |||||
| Other pastures | |||||||||||||||||||
| D. glomerata | 0.013 | 0.027 | 0.067 | 0.033 | 0.049 | 0.035 | 0.043 | 0.023 | 0.018 | 0.032 | 0.037 | 0.035 | 0.023 | 0.027 | 0.019 | 0.018 | 0.060 | 0.025 | 0.027 |
| x Festulolium | 0.011 | 0.008 | 0.008 | 0.030 | 0.028 | 0.016 | 0.033 | 0.021 | 0.007 | 0.008 | 0.012 | 0.015 | 0.022 | 0.022 | 0.003 | 0.010 | |||
| Phalaris spp. | 0.008 | 0.019 | 0.012 | 0.009 | 0.005 | 0.001 | 0.005 | 0.002 | 0.007 | 0.002 | 0.002 | <0.001 | <0.001 | 0.001 | |||||
| P. lanceolata | <0.001 | 0.001 | <0.001 | 0.001 | 0.001 | <0.001 | <0.001 | 0.002 | 0.001 | <0.001 | 0.001 | 0.001 | |||||||
| C. intybus | <0.001 | <0.001 | <0.001 | <0.001 | 0.001 | 0.001 | <0.001 | 0.001 | <0.001 | <0.001 | 0.001 | 0.001 | |||||||
| Lotus spp. | 0.004 | 0.003 | 0.003 | 0.004 | 0.004 | 0.002 | |||||||||||||
| Bromus spp. | 0.001 | <0.001 | 0.001 | 0.001 | 0.001 | 0.001 | 0.001 | ||||||||||||
| P. pratense | <0.001 | ||||||||||||||||||
| Not determined 1 | 0.001 | 0.001 | 0.012 | 0.007 | 0.012 | 0.001 | 0.011 | 0.007 | 0.006 | 0.006 | 0.011 | 0.015 | 0.006 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moscoso, C.J.; Ortega-Klose, F.; Acuña, A. Are Chilean Pasture Seed End-Users Adopting New Species? Trends and Joinpoint Regression Analysis of the Last 19 Years of Seed Imports. Agriculture 2021, 11, 552. https://doi.org/10.3390/agriculture11060552
Moscoso CJ, Ortega-Klose F, Acuña A. Are Chilean Pasture Seed End-Users Adopting New Species? Trends and Joinpoint Regression Analysis of the Last 19 Years of Seed Imports. Agriculture. 2021; 11(6):552. https://doi.org/10.3390/agriculture11060552
Chicago/Turabian StyleMoscoso, Cristian J., Fernando Ortega-Klose, and Alejandra Acuña. 2021. "Are Chilean Pasture Seed End-Users Adopting New Species? Trends and Joinpoint Regression Analysis of the Last 19 Years of Seed Imports" Agriculture 11, no. 6: 552. https://doi.org/10.3390/agriculture11060552
APA StyleMoscoso, C. J., Ortega-Klose, F., & Acuña, A. (2021). Are Chilean Pasture Seed End-Users Adopting New Species? Trends and Joinpoint Regression Analysis of the Last 19 Years of Seed Imports. Agriculture, 11(6), 552. https://doi.org/10.3390/agriculture11060552