Abstract
After nearly 40 years of DNA molecular marker development in plant breeding, the wheat research community has amassed an extensive collection of molecular markers which have been widely and successfully used for selection of agronomic, physiological and disease resistance traits in wheat breeding programs. Tan spot is a major fungal disease of wheat and a significant global economic challenge and is caused by the necrotrophic fungal pathogen Pyrenophora tritici-repentis (Ptr). Here, the potential for using a PCR-based marker (Ta1AS3422) present on the short arm of wheat chromosome 1A, was evaluated for effectiveness in distinguishing tan spot disease susceptibility. The marker was initially screened against 40 commercial Australian hexaploid wheat varieties, and those that amplified the marker had an overall lower disease score (2.8 ± 0.7 for seedlings and 2.4 ± 0.4 for plants at the tillering stage), compared to those lacking the marker which exhibited a higher disease score (3.6 ± 0.8 for both growth stages). The potential of Ta1AS3422 as a marker for the tan spot disease response was further assessed against a panel of 100 commercial Australian hexaploid wheat varieties. A significant association was observed between marker absence/presence and tan spot disease rating (Pearson’s chi-squared test, χ2 (6) = 20.53, p = 0.002), with absence of Ta1AS3422 associated with susceptibility. This simple and cost-effective PCR-based marker may be useful for varietal improvement against tan spot, although further work is required to validate its effectiveness.
1. Introduction
Tan spot, also known as yellow (leaf) spot, is a major disease of wheat globally and poses a significant economic challenge. The disease is caused by the necrotrophic fungal pathogen Pyrenophora tritici-repentis (Died.) (abbreviated to Ptr) and is easily recognisable by its symptomatic tan-coloured foliar lesions, often surrounded by chlorotic halos. The formulation of disease symptoms is a complex interaction between the host and pathogen, leading to a varying degree of necrosis and/or chlorosis depending on the genetic background of the host and virulence of the pathogen.
Ptr is known to secrete necrotrophic effectors (NE), also called host-selective toxins, which facilitate disease development in wheat lines that harbour the corresponding NE sensitivity genes [1,2]. In the tan spot pathosystem, three Ptr NEs have been described (ToxA, ToxB and ToxC), of which ToxA is the most well studied and has been shown to be a potent effector that causes distinct necrotic symptoms on wheat varieties possessing the ToxA sensitivity gene, Tsn1 [1]. However, mutant Ptr strains lacking the ToxA gene (Δtoxa) are still pathogenic and able to induce disease responses [3,4], with prominent chlorosis in specific host backgrounds likely the result of other NEs. In Australia, all Ptr isolates examined to date possess the ToxA gene and lack ToxB, while the prevalence of ToxC in Australian Ptr isolates, while present, has not yet been fully determined [3,5]. Although the genetics of tan spot disease resistance is inherently complex, molecular marker development and genetic analysis has enabled the identification of gene loci associated with resistance/susceptibility, and in more advanced cases the location of the NE sensitivity gene. In Australia, the implementation of ToxA-assisted screening by the wheat breeding industry to actively select against varieties possessing the Tsn1 gene, has seen the area sown to ToxA sensitive wheat varieties reduced by approximately 1.4 million hectares from 37.5% in the 2009/10 season to 8.3% by the 2015/16 season [3,6] with a concurrent shift away from varieties that are very susceptible to tan spot towards more moderate resistance. Currently in Australia, all commercially grown wheat varieties rated as moderately resistant to moderately susceptible (MRMS) or higher to tan spot are ToxA insensitive [7].
With trait improvement the cornerstone of plant breeding, a myriad of molecular markers linked to traits of interest have been developed to assist with germplasm selection and accelerate variety improvement. DNA markers allow diverse germplasm to be screened efficiently without the need for phenotyping and enable selection of elite lines to advance further in breeding programs. Over the years, the development of genetic DNA molecular markers has progressed from DNA fingerprinting techniques based on amplified or restriction length polymorphism, to nucleotide level polymorphism by way of microsatellite or simple sequence repeats (SSR), single nucleotide polymorphisms (SNP) and genotyping-by-sequencing (GBS) using genotyping arrays such as the Affymetrix GeneChip system or Diversity Array Technology sequencing (DArTseq) [8,9,10,11].
SSRs and SNPs are both robust molecular markers for assessing genetic diversity and population structure. The affordability of next-generation sequencing technologies has seen a rise in the discovery of SNP markers in various crop species [12,13,14]. However, only a handful of these SNP loci have been validated for their utility for the trait of interest, and then converted into robust and deployable markers. Kompetitive allele-specific PCR (KASP) has emerged as an attractive technology for SNP genotyping in the selection of marker haplotypes, and more recently in the detection of wild relative species through introgression by means of chromosome-specific codominant KASP markers based on single-copy regions [15]. Comparably, SSRs are widely distributed in plant genomes with the presence of SSR markers in both coding and non-coding regions [16,17,18]. Although fewer in number than SNP markers, SSR markers are widely used and SSR databases are available publicly for various crops including wheat [19,20,21]. Moreover, studies have claimed SSR markers to be a more efficient choice in diversity analysis than SNPs due to their faster rate of mutation [22,23,24].
Over 4000 SSR markers have been developed and utilised in wheat genetic studies, and are available for numerous abiotic and biotic traits including drought [25,26], heat [27,28], fusarium head blight (Fusarium graminearum) [29,30], powdery mildew (Blumeria graminis f. sp. tritici) [31], septoria nodorum blotch (Parastagonospora nodorum) [32] and spot blotch (Bipolaris sorokiniana) [33,34]. Wheat SSR markers range from mono-nucleotide to hexa-nucleotide tandemly repeated sequences, with the density of repeats highest on chromosome 2D and lowest on chromosome 3A [20,35]. Through the ongoing improvement of the wheat genome assembly, the predicted average density of SSRs in the wheat genome has increased from 36.68 to 95.01 SSR per Mb [20,35], and given the polyploidy nature of wheat and repetitive sequences accounting for 80–90% of the genome, the predicted SSR density of the present wheat genome assembly may increase further still. SSR markers developed for use in PCR assays can be characterised as either monomorphic or polymorphic based on the number of corresponding loci. Monomorphic SSR markers are amplified from one locus only and have been shown to be useful to analyse genetic relationships and evolutionary studies in plant species [35,36]. In this study, a monomorphic microsatellite (SSR) marker on chromosome 1AS, derived from a genome-wide analysis of the wheat variety Chinese Spring [35] and designated as Ta1AS3422, was evaluated for its potential as a tool for molecular marker-assisted breeding for tan spot disease resistance.
2. Materials and Methods
2.1. Plant Materials
A total of 100 commercially available Australian hexaploid wheat varieties (Triticum aestivum) and 12 tetraploid wheat varieties (Triticum turgidum subsp. durum) were used in this study. Seeds were sown in seedling trays containing vermiculite, fertilized with Thrive (Yates) and grown in a controlled growth chamber with a 12 h photoperiod at 22 °C. Two-week-old plant leaves (first and second leaves) were then harvested for genomic DNA extraction. For each wheat line, leaf samples were collected from two independent seedlings.
2.2. Marker Selection
The SSR marker Ta1AS3422 was selected based on an initial assessment of SSR markers against a small set of tan spot differential wheat lines. Owing to the propensity of Ta1AS3422 to distinguish tan spot susceptible wheat lines from resistant lines, this study was conducted to further investigate the potential of this PCR-based marker as a useful marker for the tan spot disease response.
2.3. DNA Extraction and Genotypic Analysis
Genomic DNA was extracted from wheat leaves using the Biosprint 15 DNA extraction kit Qiagen (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The primer pair 1AS_3252820_4724_213180_2F (5′ GTGATCTGTCGTTCGCCT 3′) and 1AS_3252820_4724_213180_2R (5′ GATGTATGCCGCAAGTTCTA 3′) were used to genotype the wheat varieties, and were derived from a monomorphic microsatellite marker previously identified from the hexaploid wheat variety Chinese Spring, and validated by reverse electronic PCR (re-PCR) [35]. The marker name is abbreviated to Ta1AS3422 in this study for simplicity. A standard PCR assay was performed to detect presence/absence of Ta1AS3422, with an expected product size of 217 bp. The PCR reaction consisted of 1× MyTaq Reaction Buffer, 0.4 U of MyTaq polymerase, 0.25 µM of each primer and 50 ng of DNA. Thermal cycling conditions consisted of an initial incubation at 94 °C for 3 min, followed by 35 cycles of 94 °C for 30 s, 57 °C for 30 s and 72 °C for 30 s. PCR products were analysed on 1.8% agarose gels. In the incidence of heterogeneous amplification, genomic DNA was extracted from two additional seedlings to confirm the presence or absence of the PCR amplification product.
2.4. Sequence Analysis
The wheat 1A chromosome reference sequence was retrieved from the IWGSC Seq Repository database (https://urgi.versailles.inra.fr/download/iwgsc/IWGSC_RefSeq_Annotations/v1.0/), and the general feature format associated with 1A was retrieved from ftp://ftp.ensemblgenomes.org/pub/plants/release-43/gff3/triticum_aestivum. The assembled wheat gene annotations adjacent to the Ta1AS3422 marker were visualised using the Diversity Among Wheat geNomes (DAWN) integrative genomics viewer (http://crobiad.agwine.adelaide.edu.au/dawn/). The TraesCS1A02G114400.1 protein sequence was retrieved from https://plants.ensembl.org/Triticum_aestivum/Info/Index and used for BLASTp homology searches against the T. aestivum genome sequence (Chinese Spring) in the Ensembl plants database. Primer and probe sequences for genetic markers were retrieved from the GrainGenes marker database (https://wheat.pw.usda.gov/cgi-bin/GG3/), the DArT sequence database (https://www.diversityarrays.com/technology-and-resources/sequences/) and from Manifesto et al. [37].
2.5. Phenotypic and Statistical Analysis
Tan spot disease resistance ratings of the 100 Australian commercial wheat varieties were obtained from the Australian crop variety sowing guides (https://grdc.com.au/resources-and-publications/all-publications/crop-variety-guides). Disease ratings consisted of the following nine categories, R = resistant, RMR = resistant to moderately resistant, MR = moderately resistant, MRMS = moderately resistant to moderately susceptible, MS = moderately susceptible, MSS = moderately susceptible to susceptible, S = susceptible, SVS = susceptible to very susceptible, and VS = very susceptible.
The disease scores of wheat varieties following infection with a ToxA gene deletion mutant (Δtoxa) of a Ptr race 1 isolate (M4) were obtained from our previous study [3]. In this earlier study 40 varieties at the seedling stage (GS13) and 15 at the tillering stage (GS25) were inoculated with 2500 conidia mL−1 and disease reactions were evaluated after 7 days [3], using the 1–5 tan spot scoring scale, where 1 = presence of resistant specks; 2 = lesions with little necrosis and chlorosis; 3 = lesions with distinct necrosis and chlorosis; 4 = coalescing type 3 lesions; 5 = extensive necrosis and chlorosis in the absence of well-defined borders between lesions [38]. Plants were grown in a controlled growth chamber (Conviron) under a 12 h photoperiod and maintained at 22 °C with a relative humidity of 40% and a light intensity of 300 μmol m−2 s−1. All infections were independently repeated using three biological replicates per variety. All statistical analyses were performed using JMP (v. 11.0.0) software. Pearson’s chi-squared test was used to assess the association between Ta1AS3422 presence/absence and tan spot disease rating, and Student’s t-test was used to compare marker presence/absence with the mean disease scores following Δtoxa mutant infection.
3. Results
Due to the potent necrosis induced by the prevalent Ptr effector ToxA, ToxA insensitive varieties (Tsn1) are favoured in Australia [3]. Therefore, the potential of Ta1AS3422 to distinguish tan spot disease susceptibility was evaluated in the absence of the ToxA–Tsn1 interaction, using a ToxA-deleted Ptr strain (Δtoxa) [3,39]. The disease scores of 40 hexaploid wheat varieties at the seedling stage and 15 varieties at the tillering stage previously infected with the Δtoxa mutant strain were used to assess the effectiveness of the marker in differentiating susceptible and resistant responses. As shown in Table 1, the mean disease scores between varieties that amplified the marker and those that did not were significantly different (Student’s t-test, p < 0.05). A lower mean disease score was observed for those varieties in which Ta1AS3422 was detected (2.8 ± 0.7 for seedlings and 2.4 ± 0.4 for plants at the tillering stage), whilst those that lacked the marker had a higher disease score (3.6 ± 0.8 for both growth stages). This latter group included SVS and S varieties such as Yitpi, Phantom, Harper and Scout. The distribution of the data is shown in Figure S1.

Table 1.
Comparison of mean disease scores of hexaploid wheat varieties following infection with a Δtoxa Ptr mutant at seedling and tillering growth stages [3].
To further verify this observation, a total of 100 Australian bread wheat (hexaploid) varieties and 12 durum (tetraploid) varieties with tan spot ratings ranging from MR to SVS, were tested for the presence or absence of Ta1AS3422 via PCR (Figure 1). Of the hexaploid wheats, 83 amplified the Ta1AS3422 marker, while it was not detected in 17 varieties (Table 2). No amplification of the marker was detected for any of the 12 durum varieties (Table 2). A significant association (Pearson’s chi-squared test, χ2 (6) = 20.53, p = 0.002) between Ta1AS3422 absence/presence and tan spot disease rating was observed. Of the 17 hexaploid varieties where the marker was not detected, 9 (52.9%) had a susceptible rating (S to SVS), while only 15 (18%) of the 83 varieties containing the marker were categorised as S to SVS (Figure 2). The majority of the resistant varieties contained the marker, with 95.7% as MRMS (22 out of 23) and 71.4% as MR (5 out of 7).

Figure 1.
PCR-based detection of the Ta1AS3422 marker. Example of presence or absence of Ta1AS3422 marker in various commercial bread wheat (hexaploid) and durum (tetraploid) varieties analysed via agarose gel electrophoresis. For each variety, the PCR assay was performed on two DNA samples extracted from two independent seedlings (1 and 2).
Table 2.
Presence/absence of Ta1AS3422 in Australian bread (hexaploid) wheat and durum (tetraploid) varieties.
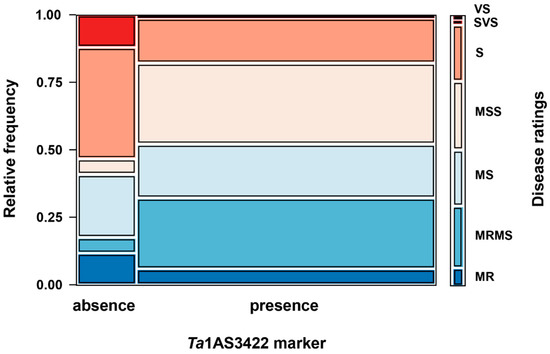
Figure 2.
Mosaic plot displaying the distribution of Ta1AS3422 presence/absence and tan spot disease ratings of 100 Australian commercial bread (hexaploid) wheat varieties. The colour refers to the tan spot disease rating (indicated on the far right) and shows the overall proportion of the observed disease that could be attributed to a specific rating, with the Y-axis showing the probability. The X-axis denotes the proportion of varieties amplifying (presence) and not amplifying (absence) the Ta1AS342 marker.
The Ta1AS3422 marker is located in the distal region of the short arm of chromosome 1A (1AS), at 118,557,370–118,557,586 bp in the genome sequenced bread wheat variety Chinese Spring. This locus is located approximately 100 Mb away from eight QTL markers on 1AS which are reported to have a significant association with tan spot disease (Table S1 and Figure S2). No QTLs associated with tan spot disease have been reported at the Ta1AS3422 region.
A 915 bp length gene (TraesCS1A02G114400.1) encoding a CCCH zinc finger-like domain, is annotated as adjacent to the Ta1AS3422 marker, with a 4 bp sequence overlap between the Ta1AS3422 forward primer and the 3′ end of the gene (Figure S3), based on the IWGSC RefSeq v1.0 (high confidence version) wheat genome assembly. A BLASTp search against the Chinese Spring genome showed that TraesCS1A02G114400.1 had no other high homology hits (>75% identity). No other gene is annotated adjacent to Ta1AS3422, with the next closest gene TraesCS1A02G114300.2 located approximately 48 Mb away (Figure S3).
4. Discussion
The Ta1AS3422 marker appears to show promise in differentiating wheat genotypes for tan spot disease severity, with varieties in which the marker amplified, displaying significantly lower mean disease scores following Δtoxa mutant infection, compared to those where the marker was absent. Furthermore, a significant association between Ta1AS3422 absence/presence and disease rating was observed across 100 hexaploid varieties, with absence associated with susceptibility. These results lend support to a potential role of Ta1AS3422 as a PCR-based DNA marker for the tan spot disease response. Of the 30 MR and MRMS hexaploid varieties examined, while the marker amplified in the majority, it appeared absent in two MR varieties (Magenta and Tennant), and the MRMS variety Sunvex, as might be expected for a non-perfect marker in different genetic backgrounds.
Traditionally, monomorphic SSR loci have not been considered to be useful for population or diversity studies due to their lack of polymorphism. Monomorphic markers can be used as a starting point for genetic studies within a particular region of interest, if markers distal to the gene of interest lack polymorphism. For example, in the case of the wheat stem rust resistance gene Sr13, no closely-linked markers associated with Sr13 were found to be polymorphic in the mapping populations examined [40]. However, sequence-tagged site (STS) markers that were monomorphic were used in combination with polymorphic markers to facilitate marker-assisted selection of Sr13 in a segregating population [41].
The utility and application of marker-assisted selection (MAS) in conventional breeding is typically based on the availability of tightly linked DNA markers to the traits, mainly derived from QTL analysis. Although the Ta1AS3422 marker was not derived from a mapping population segregating for tan spot disease response, the marker haplotypes of tightly linked markers identified from QTL analyses of mapping populations do not always have a significant association with traits of interest. For example, a lack of association was observed between the haplotypes of five closely linked markers for Sr13 and wheat stem rust resistance [40].
Monomorphic SSR markers present in one locus enable the identification of the chromosomal location of the markers and thus identification of genomic regions of interest. The physical locations of over 124,000 genes are now available through the genome assembly of Chinese Spring [42], and the Ta1AS3422 marker was found targeting a single locus on chromosome 1AS at the physical location adjacent to a CCCH motif containing zinc finger protein (TraesCS1A02G114400.1), which belongs to a diverse group of regulatory proteins characterised by tandem repeats of a combination of cysteine and histidine residues in a zinc-binding domain. Given the proximity of this gene, it may be worth investigating gene expression during tan spot infection.
Although the outcome of NE–host sensitivity gene interactions vary depending on the host genotype, the consensus is that the removal of effector sensitivity genes from the host through breeding efforts contributes to the improvement of tan spot resistance [3,43,44,45], coupled with the introgression of resistance genes into breeding material to improve germplasm [46]. The potential utility of the novel SSR marker molecular presented herein makes it a promising candidate to explore further. Additionally, the convenience, simplicity of use, ease of scoring, wide genomic distribution and low cost of SSRs makes them attractive markers for use in plant breeding programs. However, this study represents a preliminary investigation, and further work is required to validate the effectiveness of the marker for predicting disease susceptibility, for example by evaluation and allele assessment of a wheat panel comprised of broad genetic backgrounds with contrasting tan spot phenotypes through a genome wide association study (GWAS), to further determine the extent of the marker association with the tan spot disease response, and thus the utility of Ta1AS3422.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/agriculture11060513/s1, Table S1: Published significant QTL markers associated with tan spot resistance on wheat chromosome 1AS, Figure S1: Distribution of mean disease scores of hexaploid wheat varieties following infection with the Δtoxa Ptr mutant at seedling and tillering growth stages. Figure S2: Genomic location on wheat chromosome 1A of previously reported QTL markers related to tan spot disease resistance, Figure S3: Location of Ta1AS3422 on the Chinese Springe 1A chromosome.
Author Contributions
P.T.S. designed the experiments. P.T.S. carried out the experiments and wrote the paper. C.S.M. revised the manuscript. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Grains Research and Development Corporation (GRDC) and Curtin University (project code CUR00023).
Acknowledgments
The authors thank the National variety trials (NVT) for the provision of seeds.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Oliver, R.; Tan, K.-C.; Moffat, C. Necrotrophic pathogens of wheat. In Encyclopedia of Food Grains, 2nd ed.; Colin, W., Harold, C., Koushik, S., Jon, F., Eds.; Academic Press: Amsterdam, The Netherlands, 2016; pp. 273–278. ISBN 9780123947864. [Google Scholar] [CrossRef]
- Tan, K.-C.; Oliver, R.P.; Solomon, P.S.; Moffat, C.S. Proteinaceous necrotrophic effectors in fungal virulence. Funct. Plant. Biol. 2010, 37, 907–912. [Google Scholar] [CrossRef]
- See, P.T.; Marathamuthu, K.A.; Iagallo, E.M.; Oliver, R.P.; Moffat, C.S. Evaluating the importance of the tan spot ToxA–Tsn1 interaction in Australian wheat varieties. Plant. Pathol. 2018, 67, 1066–1075. [Google Scholar] [CrossRef]
- Manning, V.A.; Ciuffetti, L.M. Necrotrophic Effector Epistasis in the Pyrenophora tritici-repentis-Wheat Interaction. PLoS ONE 2015, 10, e0123548. [Google Scholar] [CrossRef]
- See, P.T.; Chen, K.; Marathamuthu, K.A.; Wood, B.; Schultz, N.; Shankar, M.; Moffat, C.S. Virulence assessment of an isolate collection of the tan spot pathogen Pyrenophora tritici-repentis in Australian wheat cultivars. Plant. Pathol. under review.
- Moffat, C.S.; Santana, F.M. Diseases Affecting Wheat: Tan Spot. In Integrated Disease Management of Wheat and Barley; Oliver, R., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2018. [Google Scholar]
- See, P.T.; Iagllo, E.M.; Oliver, R.P.; Moffat, C.S. Heterologous expression of the Pyrenophora tritici-repentis effector prpteins ToxA and ToxB, and the prevalence of effector sensitivity in Australian cereal crops. Front. Microbiol. 2019, 10, 182. [Google Scholar] [CrossRef]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant. Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Cruz, V.M.V.; Kilian, A.; Dierig, D.A. Development of DArT Marker Platforms and Genetic Diversity Assessment of the U.S. Collection of the New Oilseed Crop Lesquerella and Related Species. PLoS ONE 2013, 8, e64062. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Bernardo, A.N.; Bradbury, P.J.; Ma, H.; Hu, S.; Bowden, R.L.; Buckler, E.S.; Bai, G. Discovery and mapping of single feature polymorphisms in wheat using Affymetrix arrays. BMC Genom. 2009, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Scheben, A.; Verpaalen, B.; Lawley, C.T.; Chan, C.-K.K.; Bayer, P.E.; Batley, J.; Edwards, D. CropSNPdb: A database of SNP array data for Brassica crops and hexaploid bread wheat. Plant. J. 2019, 98, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Andorf, C.M.; Cannon, E.K.; Portwood, J.L., 2nd; Gardiner, J.M.; Harper, L.C.; Schaeffer, M.L.; Braun, B.L.; Campbell, D.A.; Vinnakota, A.G.; Sribalusu, V.V.; et al. MaizeGDB update: New tools, data and interface for the maize model organism database. Nucleic Acids Res. 2016, 44, D1195–D1201. [Google Scholar] [CrossRef] [PubMed]
- Ganal, M.W.; Altmann, T.; Röder, M.S. SNP identification in crop plants. Curr. Opin. Plant. Biol. 2009, 12, 211–217. [Google Scholar] [CrossRef]
- Grewal, S.; Hubbart-Edwards, S.; Yang, C.; Devi, U.; Baker, L.; Heath, J.; Ashling, S.; Scholefield, D.; Howells, C.; Yarde, J.; et al. Rapid identification of homozygosity and site of wild relative introgressions in wheat through chromosome-specific KASP genotyping assays. Plant. Biotechnol. J. 2020, 18, 743–755. [Google Scholar] [CrossRef]
- Dossa, K.; Yu, J.; Liao, B.; Cisse, N.; Zhang, X. Development of Highly Informative Genome-Wide Single Sequence Repeat Markers for Breeding Applications in Sesame and Construction of a Web Resource: SisatBase. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef]
- Deng, P.; Wang, M.; Feng, K.; Cui, L.; Tong, W.; Song, W.; Nie, X. Genome-wide characterization of microsatellites in Triticeae species: Abundance, distribution and evolution. Sci. Rep. 2016, 6, 32224. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, L.; Xu, Y.; Chen, C.; Rong, T.; Ali, F.; Zhou, S.; Wu, F.; Liu, Y.; Wang, J.; et al. Development and characterization of simple sequence repeat markers providing genome-wide coverage and high resolution in maize. DNA Res. 2013, 20, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Blake, V.C.; Woodhouse, M.R.; Lazo, G.R.; Odell, S.G.; Wight, C.P.; Tinker, N.A.; Wang, Y.; Gu, Y.Q.; Birkett, C.L.; Jannink, J.-L.; et al. GrainGenes: Centralized small grain resources and digital platform for geneticists and breeders. Database 2019, 2019. [Google Scholar] [CrossRef]
- Jaiswal, S.; Sheoran, S.; Arora, V.; Angadi, U.B.; Iquebal, M.A.; Raghav, N.; Aneja, B.; Kumar, D.; Singh, R.; Sharma, P.; et al. Putative Microsatellite DNA Marker-Based Wheat Genomic Resource for Varietal Improvement and Management. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef]
- Song, Q.J.; Shi, J.R.; Singh, S.; Fickus, E.W.; Costa, J.M.; Lewis, J.; Gill, B.S.; Ward, R.; Cregan, P.B. Development and mapping of microsatellite (SSR) markers in wheat. Theor. Appl. Genet. 2005, 110, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Choudhury, D.R.; Singh, A.K.; Kumar, S.; Srinivasan, K.; Tyagi, R.K.; Singh, N.K.; Singh, R. Comparison of SSR and SNP markers in estimation of genetic diversity and population structure of Indian rice varieties. PLoS ONE 2013, 8, e84136. [Google Scholar] [CrossRef]
- Chao, S.; Zhang, W.; Akhunov, E.; Sherman, J.; Ma, Y.; Luo, M.-C.; Dubcovsky, J. Analysis of gene-derived SNP marker polymorphism in US wheat (Triticum aestivum L.) cultivars. Mol. Breed. 2009, 23, 23–33. [Google Scholar] [CrossRef]
- Russell, J.; Booth, A.; Fuller, J.; Harrower, B.; Hedley, P.; Machray, G.; Powell, W. A comparison of sequence-based polymorphism and haplotype content in transcribed and anonymous regions of the barley genome. Genome 2004, 47, 389–398. [Google Scholar] [CrossRef]
- Peleg, Z.; Fahima, T.; Krugman, T.; Abbo, S.; Yakir, D.; Korol, A.B.; Saranga, Y. Genomic dissection of drought resistance in durum wheat x wild emmer wheat recombinant inbreed line population. Plant. Cell Environ. 2009, 32, 758–779. [Google Scholar] [CrossRef]
- Pinto, R.S.; Reynolds, M.P.; Mathews, K.L.; McIntyre, C.L.; Olivares-Villegas, J.J.; Chapman, S.C. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Appl. Genet. 2010, 121, 1001–1021. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Mason, R.E.; Huggins, T.; Hays, D.B. QTL on wheat (Triticum aestivum L.) chromosomes 1B, 3D and 5A are associated with constitutive production of leaf cuticular wax and may contribute to lower leaf temperatures under heat stress. Euphytica 2015, 201, 123–130. [Google Scholar] [CrossRef]
- Awlachew, Z.T.; Singh, R.; Kaur, S.; Bains, N.S.; Chhuneja, P. Transfer and mapping of the heat tolerance component traits of Aegilops speltoides in tetraploid wheat Triticum durum. Mol. Breed. 2016, 36, 78. [Google Scholar] [CrossRef]
- Wiśniewska, H.; Surma, M.; Krystkowiak, K.; Adamski, T.; Kuczyńska, A.; Ogrodowicz, P.; Mikołajczak, K.; Belter, J.; Majka, M.; Kaczmarek, Z.; et al. Simultaneous selection for yield-related traits and susceptibility to Fusarium head blight in spring wheat RIL population. Breed. Sci 2016, 66, 281–292. [Google Scholar] [CrossRef][Green Version]
- Zhu, Z.; Bonnett, D.; Ellis, M.; He, X.; Heslot, N.; Dreisigacker, S.; Gao, C.; Singh, P. Characterization of Fusarium head blight resistance in a CIMMYT synthetic-derived bread wheat line. Euphytica 2016, 208, 367–375. [Google Scholar] [CrossRef]
- Miranda, L.M.; Murphy, J.P.; Marshall, D.; Leath, S. Pm34: A new powdery mildew resistance gene transferred from Aegilops tauschii Coss. to common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Friesen, T.L.; Simons, K.J.; Xu, S.S.; Faris, J.D. Development, identification, and validation of markers for marker-assisted selection against the Stagonospora nodorum toxin sensitivity genes Tsn1 and Snn2 in wheat. Mol. Breed. 2009, 23, 35–49. [Google Scholar] [CrossRef]
- Kumar, S.; Röder, M.S.; Tripathi, S.B.; Kumar, S.; Chand, R.; Joshi, A.K.; Kumar, U. Mendelization and fine mapping of a bread wheat spot blotch disease resistance QTL. Mol. Breed. 2015, 35, 218. [Google Scholar] [CrossRef]
- Singh, V.; Singh, G.; Chaudhury, A.; Ojha, A.; Tyagi, B.S.; Chowdhary, A.K.; Sheoran, S. Phenotyping at hot spots and tagging of QTLs conferring spot blotch resistance in bread wheat. Mol. Biol Rep. 2016, 43, 1293–1303. [Google Scholar] [CrossRef]
- Han, B.; Wang, C.; Tang, Z.; Ren, Y.; Li, Y.; Zhang, D.; Dong, Y.; Zhao, X. Genome-Wide Analysis of Microsatellite Markers Based on Sequenced Database in Chinese Spring Wheat (Triticum aestivum L.). PLoS ONE 2015, 10, e0141540. [Google Scholar] [CrossRef]
- Nazareno, A.G.; dos Reis, M.S. The same but different: Monomorphic microsatellite markers as a new tool for genetic analysis. Am. J. Bot 2011, 98, e265-267. [Google Scholar] [CrossRef] [PubMed]
- Manifesto, M.; Schlatter, R.A.; Hopp, H.; Suárez, Y.E.; Dubcovsky, J. Quantitative Evaluation of Genetic Diversity in Wheat Germplasm Using Molecular Markers. Crop Sci. 2001, 41, 682–690. [Google Scholar] [CrossRef]
- Lamari, L.; Bernier, C.C. Evaluation of wheat lines and varieties to tan spot Pyrenophora triciti-repentis based on lesion type. Can. J. Plant. Pathol. 1989, 32, 4–10. [Google Scholar] [CrossRef]
- Moffat, C.S.; See, P.T.; Oliver, R.P. Generation of a ToxA knockout strain of the wheat tan spot pathogen Pyrenophora tritici-repentis. Mol. Plant. Pathol. 2014, 15, 918–926. [Google Scholar] [CrossRef]
- Simons, K.; Abate, Z.; Chao, S.; Zhang, W.; Rouse, M.; Jin, Y.; Elias, E.; Dubcovsky, J. Genetic mapping of stem rust resistance gene Sr13 in tetraploid wheat (Triticum turgidum ssp. durum L.). Theor. Appl. Genet. 2011, 122, 649–658. [Google Scholar] [CrossRef]
- Periyannan, S.K.; Qamar, Z.U.; Bansal, U.K.; Bariana, H.S. Development and validation of molecular markers linked with stem rust resistance gene Sr13 in durum wheat. Crop. Pasture Sci. 2014, 65, 74–79, 76. [Google Scholar] [CrossRef]
- A chromosome-based draft sequence of the hexaploid bread wheat (Triticum aestivum) genome. Science 2014, 345, 1251788. [CrossRef]
- Friesen, T.L.; Ali, S.; Kianian, S.; Francl, L.J.; Rasmussen, J.B. Role of host sensitivity to ptr ToxA in development of tan spot of wheat. Phytopathology 2003, 93, 397–401. [Google Scholar] [CrossRef]
- Chu, C.G.; Xu, S.S.; Faris, J.D.; Nevo, E.; Friesen, T.L. Seedling Resistance to Tan Spot and Stagonospora nodorum Leaf Blotch in Wild Emmer Wheat (Triticum dicoccoides). Plant. Dis. 2008, 92, 1229–1236. [Google Scholar] [CrossRef]
- Faris, J.D.; Abeysekara, N.S.; McClean, P.E.; Xu, S.S.; Friesen, T.L. Tan spot susceptibility governed by the Tsn1 locus and race-nonspecific resistance quantitative trait loci in a population derived from the wheat lines Salamouni and Katepwa. Mol. Breed. 2012, 30, 1669–1678. [Google Scholar] [CrossRef]
- Shankar, M.; Mather, D.; Jorgensen, D.; Golzar, H.; Chalmers, K.; Hollaway, G.; McLean, M.; Neate, S.; Loughman, R. Germplasm Enhancement for Resistance to Pyrenophora Tritici-Repentis in Wheat. In Advances in Wheat Genetics: From Genome to Field, Proceedings of the 12th International Wheat Genetics Symposium; Ogihara, Y., Takumi, S., Handa, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 193–199. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).