
Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region


Abstract

1. Introduction
2. Materials and Methods
2.1. Sites and Samples
2.2. Soil Physicochemical Analysis
2.3. Extraction of AM Spores from Field Soils
2.4. AM Fungal Colonization of Rubber Roots
2.5. Molecular Analysis of Rubber Roots
2.6. Bioinformatics
2.7. Statistical Analysis
3. Results
3.1. Soil Conditions of Rubber Plantations
3.2. Abundance of Arbuscular Mycorrhizal Spores in Soils
3.3. Arbuscular Mycorrhizal Fungal Colonization of Rubber Roots
3.4. Arbuscular Mycorrhizal Fungal Community in Rubber Roots
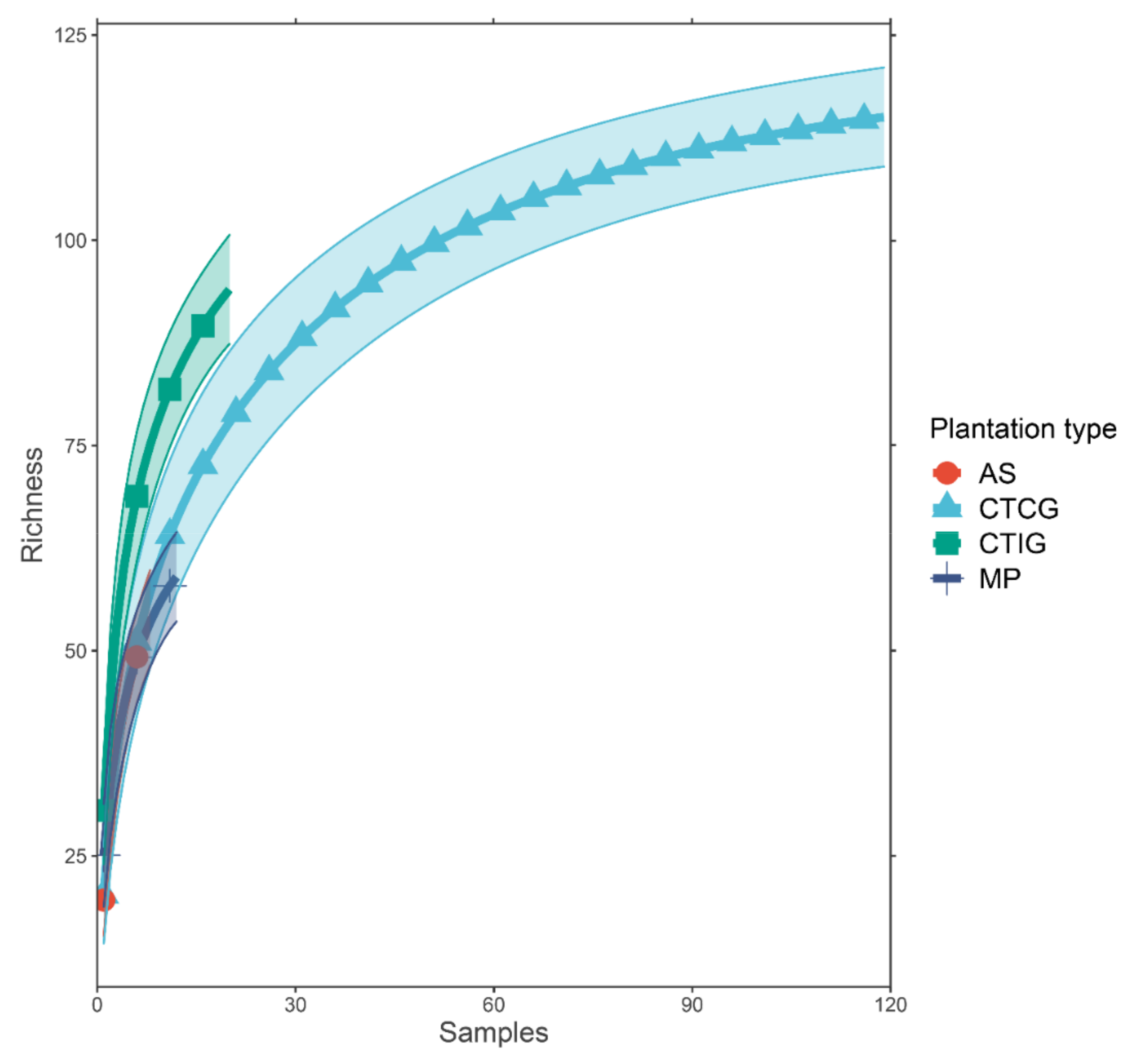
3.5. Richness and Diversity of Arbuscular Mycorrhizal Fungal in Rubber Roots
3.6. Arbuscular Mycorrhizal Fungi and Soil Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Verheye, W. Growth and production of rubber. In Land Use, Land Cover and Soil Sciences. Encyclopedia of Life Support Systems (EQLSS); Verheye, W., Ed.; UNESCO-EOLSS Publishers: Oxford, UK, 2010; p. 16. [Google Scholar]
- International Rubber Study Group. Rubber Statistical Bulletin; International Rubber Study Group: Singapore, 2019. [Google Scholar]
- Chen, C.; Liu, W.; Jiang, X.; Wu, J. Effects of rubber-based agroforestry systems on soil aggregation and associated soil organic carbon: Implications for land use. Geoderma 2017, 299, 13–24. [Google Scholar] [CrossRef]
- Sterling, A.; Martínez-Vuiche, E.J.; Suárez-Córdoba, Y.D.; Agudelo-Sánchez, A.A.; Fonseca-Restrepo, J.A.; Andrade-Ramírez, T.K.; Virguez-Díaz, Y.R. Assessing growth, early yielding and resistance in rubber tree clones under low South American Leaf Blight pressure in the Amazon region, Colombia. Ind. Crops Prod. 2020, 158, 112958. [Google Scholar] [CrossRef]
- Ramírez, U.; Charry, A.; Jäger, M.; Huertado, J.; Sterling, A.; Romero, M.; Sierra, L.; Quintero, M. Estrategia Sectorial de la Cadena de Caucho en Caquetá, con Enfoque Agroambiental y cero Deforestación; CIAT: Palmira, Colombia, 2018. [Google Scholar]
- Castellanos, O.; Fonseca, S.; Barón, M. Agenda prospectiva de investigación y desarrollo tecnológico para la cadena productiva de caucho natural y su industria en Colombia. Ing. Investig. 2009, 29, 63–65. [Google Scholar]
- Quesada, C.A.; Lloyd, J.; Schwarz, M.; Patiño, S.; Baker, T.R.; Czimczik, C.; Fyllas, M.N.; Martinelli, L.; Nardoto, G.B.; Schmerler, J.; et al. Variations in chemical and physical properties of Amazon forest soils in relation to their genesis. Biogeosciencies 2010, 7, 1515–1541. [Google Scholar] [CrossRef]
- He, P.; Qin, H.; Wu, M.; Wu, B.; Wei, J.; Wang, D. Identification of genes differentially expressed in the roots of rubber tree (Hevea brasiliensis Muell. Arg.) in response to phosphorus deficiency. Mol. Biol. Reprod. 2013, 40, 1397–1405. [Google Scholar] [CrossRef]
- Herrmann, L.; Lesueur, D.; Bräu, L.; Davison, J.; Jairus, T.; Robain, H.; Robin, A.; Vasar, M.; Wiriyakitnateekul, W.; Öpik, M. Diversity of root-associated arbuscular mycorrhizal fungal communities in a rubber tree plantation chronosequence in Northeast Thailand. Mycorrhiza 2016, 26, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.M.R.; da Silva, D.K.A.; Ferreira, A.C.A.; Goto, B.T.; Maia, L.C. Diversity of arbuscular mycorrhizal fungi in Atlantic forest areas under different land uses. Agric. Ecosyst. Environ. 2014, 185, 245–252. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A new fungal phylum, the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef]
- Cardoso, I.M.; Kuyper, T.W. Mycorrhizas and tropical soil fertility. Agric. Ecosyst. Environ. 2006, 116, 72–84. [Google Scholar] [CrossRef]
- Baum, C.; El-Tohamy, W.; Gruda, N. Increasing the productivity and product quality of vegetable crops using arbuscular mycorrhizal fungi: A review. Sci. Hortic. 2015, 187, 131–141. [Google Scholar] [CrossRef]
- Goldschmidt, E.E. Plant grafting: New mechanisms, evolutionary implications. Front. Plant Sci. 2014, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Aloni, B.; Cohen, R.; Karni, L.; Aktas, H.; Edelstein, M. Hormonal signaling in rootstock-scion interactions. Sci. Hortic. 2010, 127, 119–126. [Google Scholar] [CrossRef]
- Walder, F.; Niemann, H.; Natarajan, M.; Lehmann, M.F.; Boller, T.; Wiemken, A. Mycorrhizal Networks: Common Goods of Plants Shared under Unequal Terms of Trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef]
- Soka, G.; Ritchie, M. Arbuscular mycorrhizal symbiosis and ecosystem processes: Prospects for future research in tropical soils. Open J. Ecol. 2014, 4, 11–22. [Google Scholar] [CrossRef]
- Feldmann, F.; da Silva, J.J.P.; Idczak, E.; Lieberei, R. AMF spore community composition of natural and agricultural sites in Central Amazonia—A long term study. In German-Brazilian Workshop on Neotropical Ecosystems—Achievements and Prospects of Cooperative Research; FRG and EMBRAPA: Hamburg, Germany, 2000; pp. 1–31. [Google Scholar]
- Sterling, A.; Rodrìguez, C.H. Valoración de Nuevos Clones de Hevea Brasiliendis con Proyección Para la Amazonia Colombiana: Fases pre y Post-Sangría Temprana en el Caquetá; Imagen Editorial SAS: Bogotá, Colombia, 2020; p. 326. [Google Scholar]
- IGAC. Estudio General de Suelos y Zonificación de Tierras Departamento de Caquetá Escala 1:100.000; Instituto Geográfico Agustin Codazzi—IGAC: Bogotá, Colombia, 2014.
- Sterling, A.; Rodríguez, C.H. Agroforestería en el Caquetá: Clones Promisorios de Caucho en Asocio con Copoazú y Plátano Hartón con Potencial para la Amazonia Colombiana; Instituto Amazonico de Investigaciones Científicas Sinchi: Bogotá, Colombia, 2014. [Google Scholar]
- Sterling, A.; Rodríguez, C.H. Valoración y Análisisde la Biodiversidad y Servicios Ecosistémicos Asociados a Campos Clonales de Caucho en Caquetá, Amazonia Colombiana; Instituto Amazonico de Investigaciones Científicas Sinchi: Bogotá, Colombia, 2019. [Google Scholar]
- Sieverding, E. Vesicular-arbuscular mycorrhiza management in tropical ecosystems. In Deutche Gesellschaft für Technische Zusammenarbeit 224; Verlag, H.B., Ed.; Technical Cooperation Federal Republic of Germany: Eschbom, Germany, 1991. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for cleaning roots and staining parasitic and vesicular-arbuscular fungi for rapid assessment of infection. Transcr. Br. Soc. Mycol. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Ashton, P.D.; Aziz, N.; Feng, G.; Nelson, M.; Dytham, C.; Fitter, A.H.; Helgason, T. Distinct seasonal assemblages of arbuscular mycorrhizal fungi revealed by massively parallel pyrosequencing. New Phytol. 2011, 190, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, S.; Young, J.P.W. Improved PCR primers for the detection and identi¢cation of arbuscular mycorrhizal fungi. FEMS Microbiol. Ecol. 2008, 65, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Moora, M.; Semchenko, M.; Adenan, S.B.; Ahmed, T.; Akhmetzhanova, A.A.; Alatalo, J.M.; Al-Quraishy, S.; Adriyanova, E.; Anslan, S.; et al. Temperature and pH define the realized niche space of arbuscular mycorrhizal fungi. New Phytol. 2021. [Google Scholar] [CrossRef]
- Vasar, M.; Andreson, R.; Davison, J.; Jairus, T.; Moora, M.; Remm, M.; Young, J.P.W.; Zobel, M.; Öpik, M. Increased sequencing depth does not increase captured diversity of arbuscular mycorrhizal fungi. Mycorrhiza 2017, 27, 761–773. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Milne, I.; Lindner, D.; Bayer, M.; Husmeier, D.; McGuire, G.; Marshall, D.F.; Wright, F. TOPALi v2: A rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 2009, 25, 126–127. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Kindt, R. Package ‘BiodiversityR’: Community Ecology and Suitability Analysis; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2020. [Google Scholar]
- Dunnington, D.; Wickham, H.; Chang, W.; Henry, L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H. Package ‘ggplot2’: Create Elegant Data Visualisations Unisg the Grammar of Graphics. R Package Version 2020. Available online: https://ggplot2.tidyverse.org/reference/ggplot2-package.html (accessed on 9 January 2021).
- Yan, L. Package ‘Ggvenn’. 2021. Available online: https://cren.r-project.org/web/packages/ggvenn/ggvenn.pdf (accessed on 11 January 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’: Community Ecology Package. 2018. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 13 January 2021).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. R-Core. Nlme: Linear and Nonlinear Mixed Effects Models. 2013. Available online: https://cran.r-project.org/web/packages/nlme/index.html (accessed on 12 January 2021).
- Robert, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology, in R Package. 2012. Available online: https://cran.r-project.org/web/packages/labdsv/index.html (accessed on 13 January 2021).
- R Core Team. Core Team R: A Language and Environment for Statistical Computing. 2020. Available online: http://www.r-project.org/index.html (accessed on 12 January 2021).
- Di Rienzo, J.A.; Casanoves, F.; Pla, L.; Vilchez, S.; Di Renzo, M.J. InfoStat. 2020. Available online: http://www.infostat.com/ (accessed on 15 January 2021).
- Di Rienzo, J.A.; Casanoves, F.; Pla, L.; Vilchez, S.; Di Renzo, M.J. Qeco. 2018. Available online: https://sites.google.com/site/qecosite/home (accessed on 15 January 2021).
- R Studio. 2020. Available online: https://www.rstudio.com/products/rstudio/older-versions/ (accessed on 15 January 2021).
- Aguilar-Trigueros, C.A.; Hempel, S.; Powell, J.R.; Cornwell, W.K.; Rilling, M.C. Bridging reproductive and microbial ecology: A case study in arbuscular mycorrhizal fungi. ISME J. 2019, 13, 873–884. [Google Scholar] [CrossRef]
- Sanders, I.R. Plant and arbuscular mycorrhizal fungal diversity: Are we looking at the relevant levels of diversity and are we using the right techniques? New Phytol. 2004, 164, 415–418. [Google Scholar] [CrossRef]
- Leal, P.L.; Siqueira, J.O.; Stürmer, S.L. Switch of tropical Amazon forest to pasture affects taxonomic composition but not species abundance and diversity of arbuscular mycorrhizal fungal community. Appl. Soil Ecol. 2013, 71, 72–80. [Google Scholar] [CrossRef]
- Reyes, H.A.; Ferreira, P.F.A.; Silva, L.C.; da Costa, M.G.; Nobre, C.P.; Gehring, C. Arbuscular mycorrhizal fungi along secondary forest succession at the eastern periphery of Amazonia: Seasonal variability and impacts of soil fertility. Appl. Soil Ecol. 2019, 136, 1–10. [Google Scholar] [CrossRef]
- Stürmer, S.L.; Siqueira, J.O. Species richness and spore abundance of arbuscular mycorrhizal fungi across distinct land uses in Western Brazilian Amazon. Mycorrhiza 2011, 21, 255–267. [Google Scholar] [CrossRef]
- Pfeiffer, C.M.; Bloss, H.E. Growth and nutrition of guayule (Parthenium argentatum) in a saline soil as influenced by vesicular-arbuscular mycorrhiza and phosphorus fertilization. New Phytol. 1988, 108, 315–321. [Google Scholar] [CrossRef]
- Tawaraya, K.; Tacaya, Y.; Turjaman, M.; Tuah, S.J.; Limin, S.H.; Tamai, Y.; Cha, J.Y.; Wagatsuma, T.; Osaki, M. Arbuscular mycorrhizal colonization of tree species grown in peat swamp forest of Central Kalimantan, Indonesia. Forest Ecol. Manag. 2003, 182, 381–386. [Google Scholar] [CrossRef]
- Dumbrell, A.J.; Nelson, M.; Helgason, T.; Dytham, C.; Fitter, A.H. Idiosyncrasy and overdominance in the structure of natural communities of arbuscular mycorrhizal fungi: Is there a role for stochastic processes? J. Ecol. 2010, 98, 419–428. [Google Scholar] [CrossRef]
- Entry, J.A.; Rygiewicz, P.T.; Watrud, L.S.; Donelly, P.K. Influence of adverse soil conditions on the formation and function of Arbuscular mycorrhizas. Adv. Environ. Res. 2002, 7, 123–138. [Google Scholar] [CrossRef]
- Moora, M.; Davison, J.; Öpik, M.; Metsis, M.; Saks, Ü.; Jairus, T.; Vasar, M.; Zobel, M. Anthropogenic land use shapes the composition and phylogenetic structure of soil arbuscular mycorrhizal fungal communities. FEMS Microbiol. Ecol. 2014, 90, 609–621. [Google Scholar] [CrossRef]
- García-Montiel, D.C.; Neill, C.; Melillo, J.; Thomas, S.; Steudler, P.A.; Cerri, C.C. Soil phosphorus transformations following forest clearing for pasture in the Brazilian Amazon. Soil Sci. Soc. Am. J. 2000, 64, 1792–1804. [Google Scholar] [CrossRef]
- Clark, R.B. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and host plant growth and mineral adquisition at low pH. Plant Soil 1997, 192, 15–22. [Google Scholar] [CrossRef]
- Cuenca, G.; Cáceres, A.; González, M.G. AM inoculation in tropical agroculture: Field results. In Mycorrhiza; Varma, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 403–417. [Google Scholar]
- Izaguirre-Mayoral, M.L.; Flores, S.; Pieters, A.; Olivares, E.; Cuenca, C. Rhizophagus manihotis promotes the growth of rhizobia-nodulated Vigna luteola L. in phosphorus deficient acid montane soils devoid of ground cover vegetation. Symbiosis 2011, 55, 1–9. [Google Scholar] [CrossRef]
- Peña-Venegas, C.P.; Castro, D.; Fausto, M.; Silva, A.; Rodríguez, C.H. Comportamiento de Variables del Suelo en la Sucesión de Bosques Secundarios, in Sucesión Ecológica y Restauración en Paisajes Fragmentados de la Amazonia Colombiana; Rodríguez, C.H., Sterling, A., Eds.; Instituto Amazónico de Investigaciones Científicas Sinchi: Bogotá, Colombia, 2020; pp. 141–170. [Google Scholar]
- Sepp, S.-K.; Davison, J.; Jairus, T.; Vasar, M.; Moora, M.; Zobel, M.; Öpik, M. Non-random association patterns in a plant-mycorrhizal fungal network reveal host-symbiont specificity. Mol. Ecol. 2019, 28, 365–378. [Google Scholar] [CrossRef]
- Peña-Venegas, C.P.; Kuyper, T.W.; Davison, J.; Jairus, T.; Vasar, M.; Stomph, T.J.; Struik, P.C.; Öpik, M. Distinct arbuscular mycorrhizal fungal communities associate with different manioc landraces and Amazonian soils. Mycorrhiza 2019, 29, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Faggioli, V.S.; Cabello, M.N.; Grilli, G.; Vasar, M.; Covacevich, F.; Öpik, M. Root colonizing and soil borne communities of arbuscular mycorrhizal fungi differ among soybean fields with contrasting historical land use. Agric. Ecosyst. Environ. 2019, 269, 174–182. [Google Scholar] [CrossRef]
- Beukema, H.; Danielsen, F.; Vincent, G.; Hardiwinoto, S.; van Andel, J. Plant and bird diversity in rubber agroforests in the lowlands of Sumatra, Indonesia. Agrofor. Syst. 2007, 70, 217–242. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plantation Type | SOIL Physicochemical Variables | |||||
|---|---|---|---|---|---|---|
| pH | OC (%) | CEC (meq/100g) | Loam (%) | Clay (%) | Sand (%) | |
| CTCG | 4.62 ± 0.04a | 1.37 ± 0.24a | 10.46 ± 1.62a | 14.14 ± 0.46a | 47.77 ± 3.55a | 38.71 ± 4.03a |
| CTIG | 4.62 ± 0.05a | 0.85 ± 0.29a | 7.05 ± 1.67a | 8.14 ± 0.66b | 43.33 ± 4.38a | 48.33 ± 4.86a |
| AS | 4.55 ± 0.08a | 1.17 ± 0.29a | 5.94 ± 1.65a | 13.98 ± 0.35a | 49.00 ± 5.36a | 40.71 ± 4.99a |
| MP | 4.55 ± 0.05a | 1.15 ± 0.24a | 7.13 ± 1.54a | 8.00 ± 0.00b | 49.33 ± 4.37a | 42.67 ± 3.94a |
| K (mg/kg) | Ca (mg/kg) | Mg (mg/kg) | Na (mg/kg) | P (mg/kg) | ||
| CTCG | 67.48 ± 8.37a | 330.45 ± 30.47a | 58.90 ± 6.58a | 32.79 ± 3.26b | 1.31 ± 0.46a | |
| CTIG | 40.13 ± 10.41a | 212.29 ± 34.99a | 35.99 ± 7.72a | 29.64 ± 4.03b | 0.59 ± 0.58a | |
| AS | 38.60 ± 9.78a | 255.18 ± 40.88a | 46.00 ± 7.55a | 46.03 ± 4.00a | 1.09 ± 0.55a | |
| MP | 55.51 ± 9.13a | 238.00 ± 27.78a | 32.67 ± 6.16a | 49.32 ± 3.96a | 0.73 ± 0.45a | |
| Plantation Type | Rubber Tree Genotypes | Plantation Identity | Age (Years) | Geographic Location | Number Samples a | Total Number of AM Fungal Reads | Number of Vitual Taxa |
|---|---|---|---|---|---|---|---|
| CTCG | Promising Colombian genotypes: ECC 25, ECC 29, ECC 35, ECC 60, ECC 64, ECC 66, ECC 73, ECC 83, ECC 90 and IAN 873 (traditional cultivar) | CTCG 1 | 1 | El Paujil | 10 | 37,631 | 53 |
| CTCG 2 | 1 | San Vicente del Caguán | 10 | 32,814 | 80 | ||
| Promising Colombian genotypes (in a continuous sequence): ECC 101 to ECC 199, and IAN 873 (traditional cultivar). The genotype ECC 169 had not samples for molecular analysis | CTCG 3 | 1 | El Paujil | 99 | 140,362 | 99 | |
| CTIG | Promising introduced genotypes: CDC 312, CDC 56, FDR 4575, FDR 5597, FDR 5788, FX 3899 P1, FX 4098, GU 198 and MDF 180, and IAN 873 (traditional cultivar) | CTIG 1 | 9 | Florencia | 10 | 40,249 | 78 |
| CTIG 2 | 9 | San Vicente del Caguán | 10 | 26,622 | 78 | ||
| AS | Promising introduced genotypes: FX 4098 and FDR 5788, and IAN 873 (traditional cultivar) | AS 1 | 6 | Albania | 4 | 30,670 | 44 |
| AS 2 | 6 | San Vicente del Caguán | 4 | 4784 | 35 | ||
| MP | Introduced genotypes planted as traditional cultivars: FX 3864, FX 25 or IAN 873 | MP 1 | 30 | San Vicente del Caguán | 4 | 36,188 | 32 |
| MP 2 | 50 | Belén de los Andaquíes | 4 | 25,226 | 44 | ||
| MP 3 | 50 | Florencia | 4 | 44,543 | 50 |
| Factor | Level | Richness (S) | Exponential Shannon (expH′) | Inverse Simpson 1/D | Evenness (Piélou) |
|---|---|---|---|---|---|
| Genotype type | Colombian | 20.75 ± 2.78a | 5.26 ± 0.73a | 3.68 ± 0.40a | 0.53 ± 0.03a |
| Introduced | 25.08 ± 1.88a | 6.85 ± 0.55a | 4.67 ± 0.35a | 0.58 ± 0.02a | |
| Plantation type | CTCG | 20.80 ± 2.04b | 5.42 ± 0.44b | 3.82 ± 0.22a | 0.53 ± 0.02a |
| CTIG | 30.55 ± 2.63a | 8.13 ± 0.66a | 5.33 ± 0.43a | 0.61 ± 0.04a | |
| AS | 19.63 ± 3.13b | 4.90 ± 0.94b | 3.33 ± 0.66a | 0.51 ± 0.05a | |
| MP | 25.08 ± 2.55ab | 7.31 ± 0.77ab | 5.00 ± 0.54a | 0.60 ± 0.04a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peña-Venegas, C.P.; Sterling, A.; Andrade-Ramírez, T.K. Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region. Agriculture 2021, 11, 361. https://doi.org/10.3390/agriculture11040361
Peña-Venegas CP, Sterling A, Andrade-Ramírez TK. Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region. Agriculture. 2021; 11(4):361. https://doi.org/10.3390/agriculture11040361
Chicago/Turabian StylePeña-Venegas, Clara P., Armando Sterling, and Tatiana K. Andrade-Ramírez. 2021. "Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region" Agriculture 11, no. 4: 361. https://doi.org/10.3390/agriculture11040361
APA StylePeña-Venegas, C. P., Sterling, A., & Andrade-Ramírez, T. K. (2021). Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region. Agriculture, 11(4), 361. https://doi.org/10.3390/agriculture11040361