
Papaya (Carica papaya L.) Phenology under Different Agronomic Conditions in the Subtropics


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Meteorological Data
2.3. Morphological Features
2.4. Total Soluble Solids Content (TSS) Determination
2.5. Statistical Analysis
3. Results
3.1. Meteorological Conditions
3.2. Morphological Features
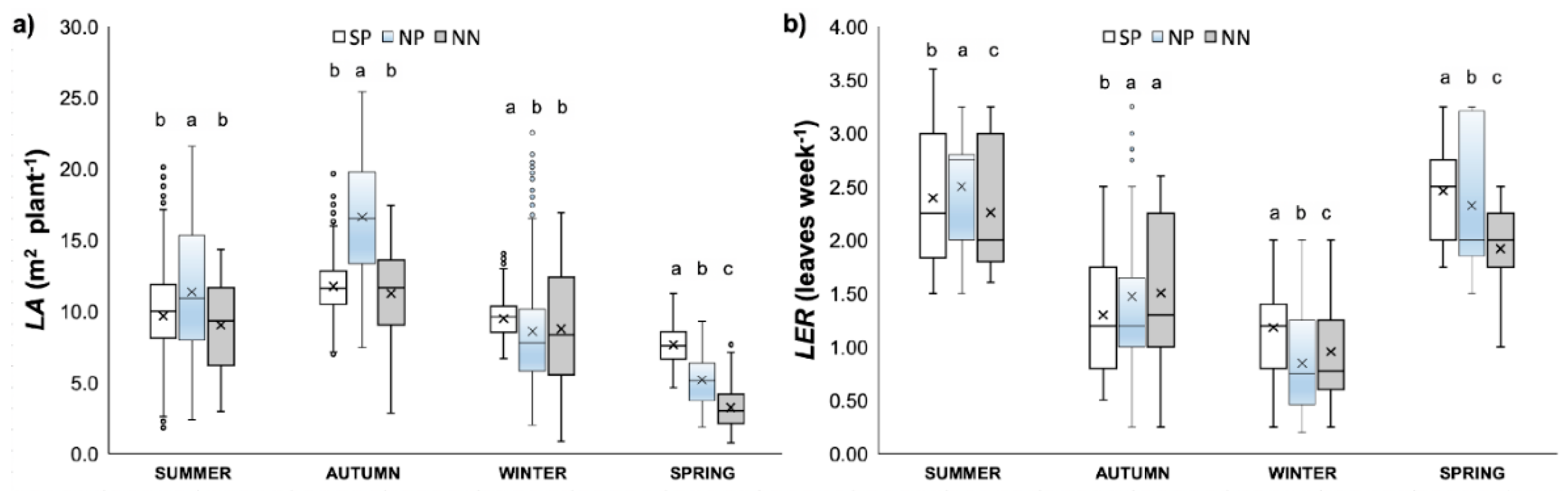
3.2.1. Phenology of the Canopy Development
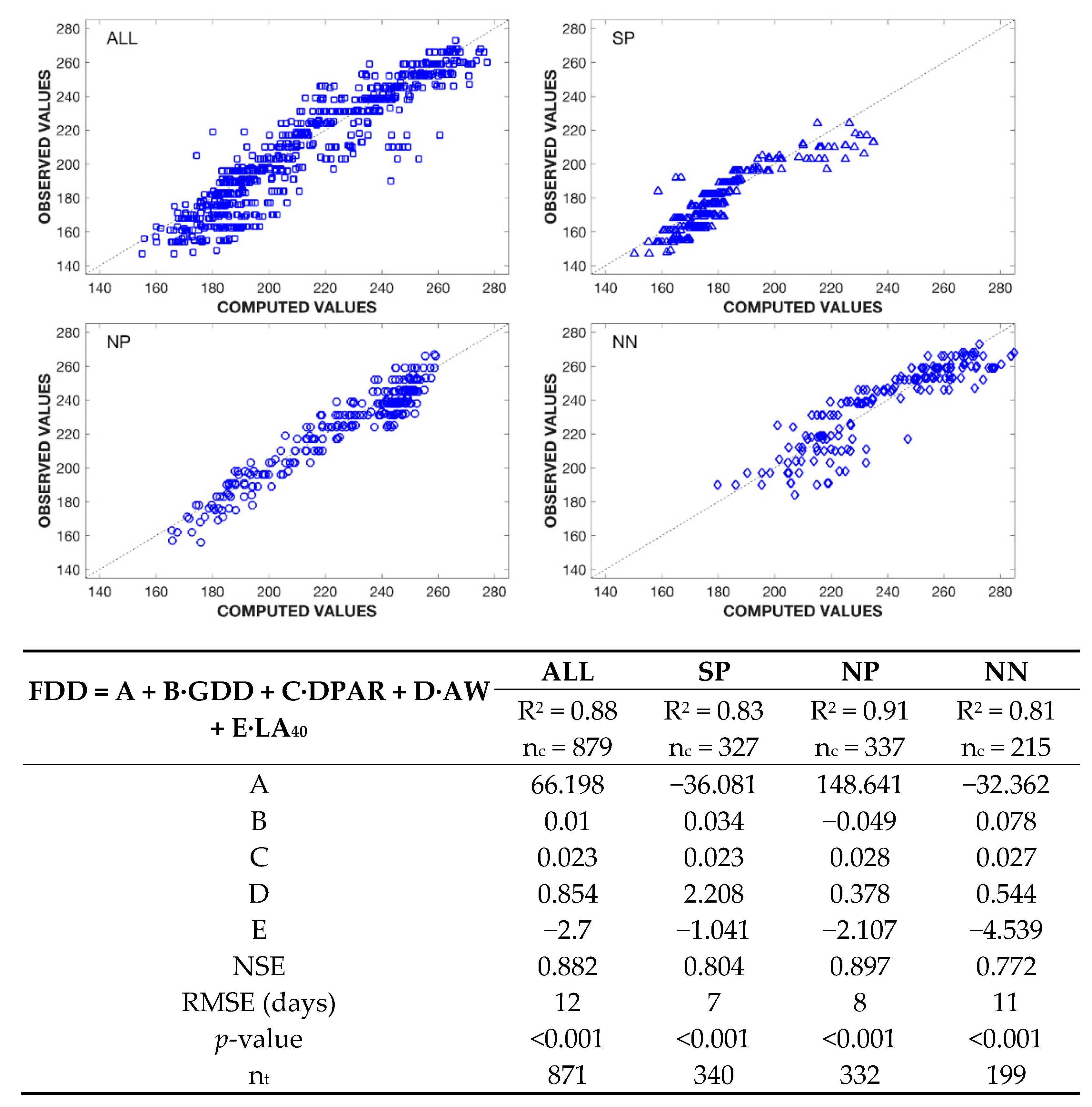
3.2.2. Phenology of the Pre-Anthesis Flower Development
3.2.3. Phenology of the Post-Anthesis Fruit Development
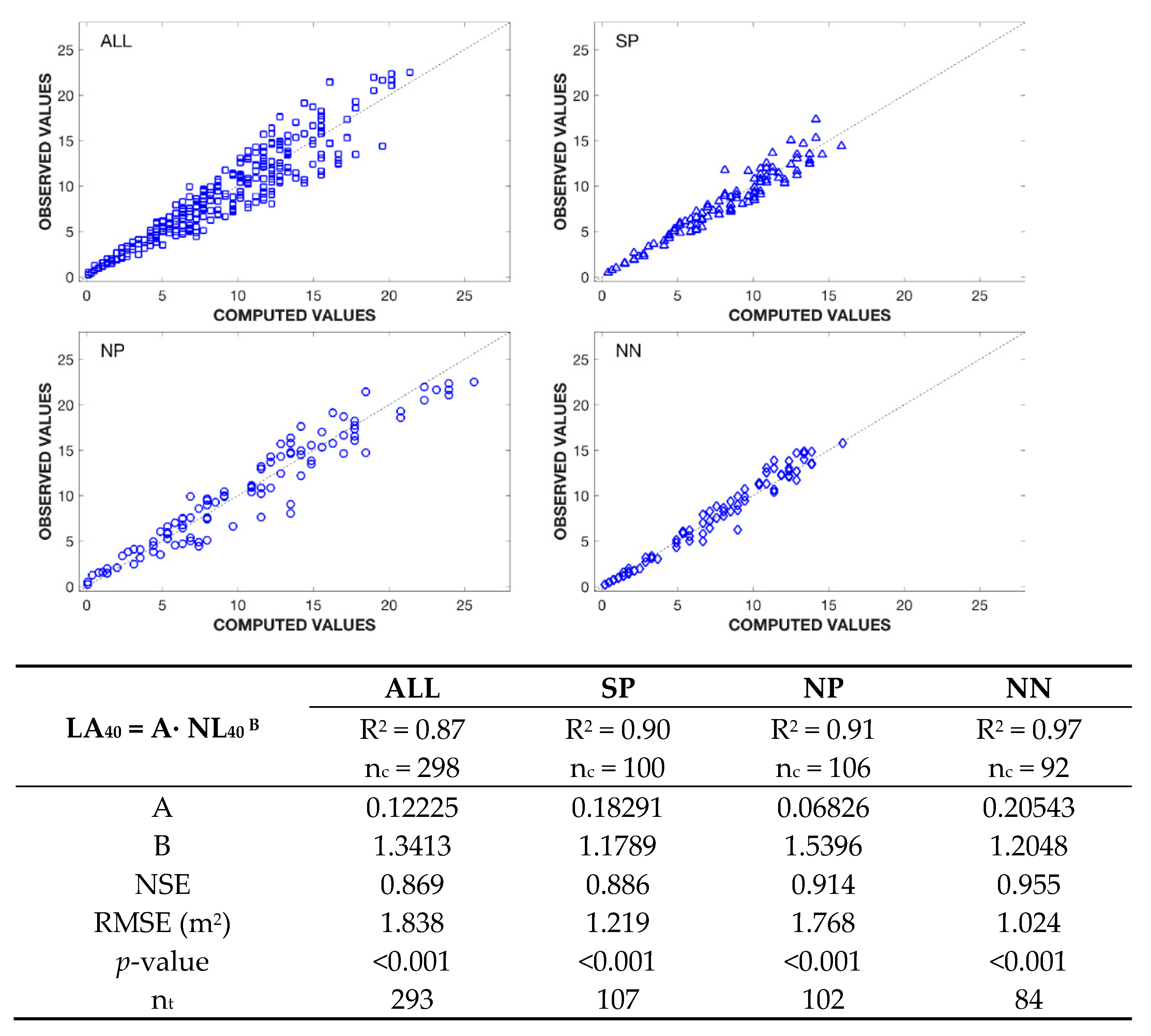
3.3. Number of Fruits—Leaf Area Ratio and Total Soluble Solids Content
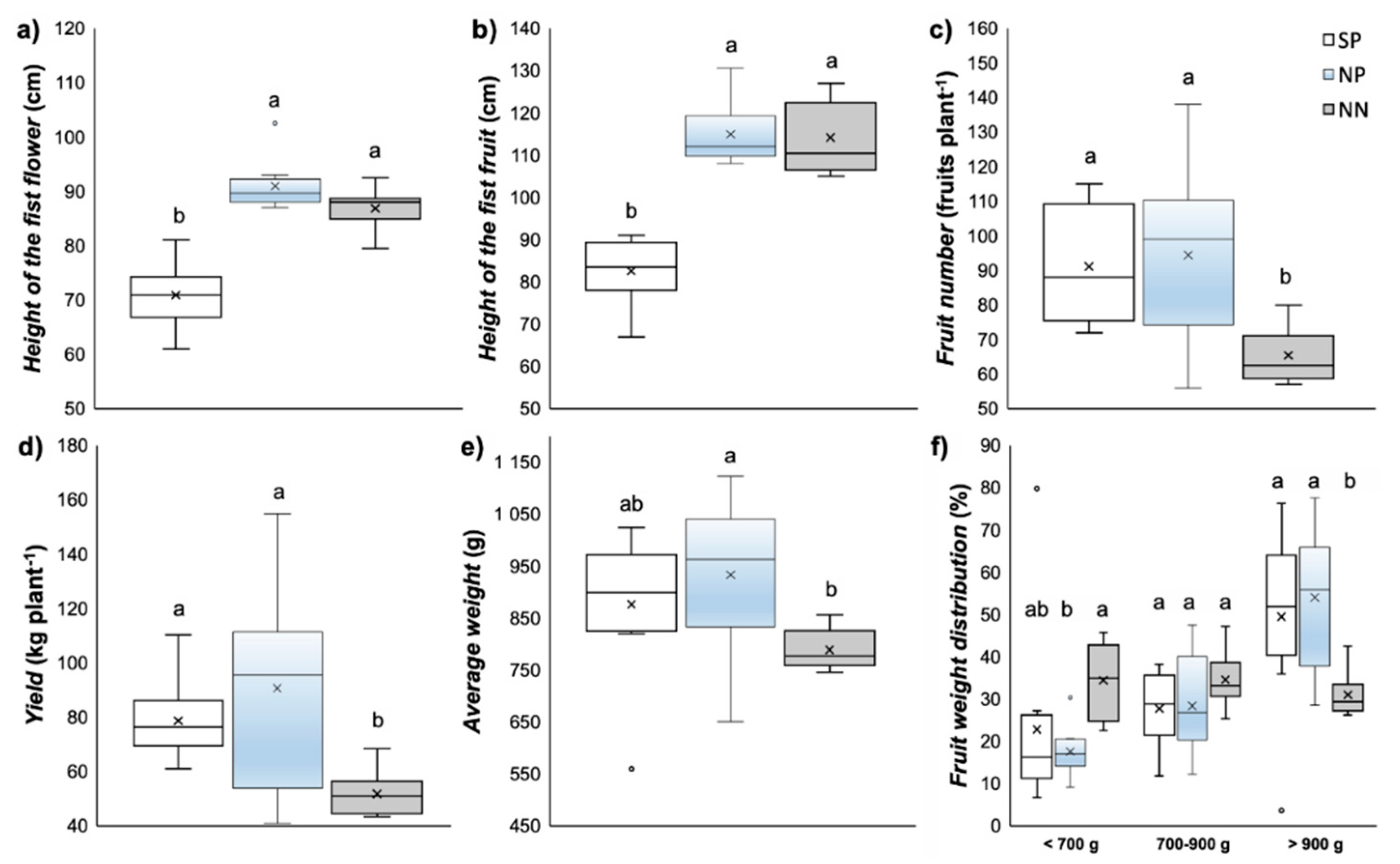
3.4. Production Parameters
4. Discussion
4.1. Leaf Area and Leaf Emission Rate
4.2. Floral Abnormalities
4.3. Fruit Set and Total Soluble Solids Content
4.4. Fruit Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teixeira, L.G.; Lages, P.C.; Jascolka, T.L.; Aguilar, E.C.; Soares, F.L.P.; Pereira, S.S.; Beltrão, N.R.M.; de Oliveira Matoso, R.; do Nascimento, A.M.; de Castilho, R.O.; et al. White tea (Camellia sinensis) extract reduces oxidative stress and triacylglycerols in obese mice. Food Sci. Technol. 2012, 32, 733–741. [Google Scholar] [CrossRef]
- Vij, T.; Prashar, Y. A review on medicinal properties of Carica papaya Linn. Asian Pac. J. Trop. Dis. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Galán Sauco, V.; Rodríguez Pastor, M.C. Greenhouse cultivation of papaya. Acta Hortic. 2007, 740, 191–195. [Google Scholar] [CrossRef]
- Campostrini, E.; Glenn, D.M. Ecophysiology of papaya: A review. Braz. J. Plant Physiol. 2007, 19, 413–424. [Google Scholar] [CrossRef]
- Martelleto, L.A.P.; Ribeiro, R.D.L.D.; Sudo-Martelleto, M.; Vasconcellos, M.A.D.S.; Pereira, M.B. Expression of female sterility and carpelloidy on papaya tree under different environments of protected cultivation. Rev. Bras. Frutic. 2011, 33, 1185–1193. [Google Scholar] [CrossRef]
- Nakasone, H.Y.; Paul, R.E. Tropical Fruits; CAB Int.: Wallingford, UK, 1998; 432p. [Google Scholar]
- Zhou, L. Source-sink Relationship during Papaya Fruit Development and Ripening. Ph.D. Thesis, University of Hawaii at Manoa, Honolulu, HI, USA, 1999. [Google Scholar]
- Stenzel, N.M.C.; Neves, C.S.V.J.; Marur, C.J.; Scholz, M.B.S.; Gomes, J.C. Maturation curves and degree-days accumulation for fruits of ‘Folha Murcha’ orange trees. Sci. Agric. 2006, 63, 219–225. [Google Scholar] [CrossRef]
- de Almeida, F.T.; Bernardo, S.; Sousa, E.F.; Marin, S.L.D.; Grippa, S. Growth and yield of papaya under irrigation. Sci. Agric. 2003, 60, 419–424. [Google Scholar] [CrossRef]
- Berilli, S.S.; Oliveira, J.G.; Marinho, A.B.; Lyra, G.B.; Sousa, E.F.; Viana, A.P.; Bernardo, S.; Pereira, M.G. Avaliação da taxa de crescimento de frutos de mamão (Carica papaya L.) em função das épocas do ano e graus-dias acumulados. Rev. Bras. Frutic. 2007, 29, 11–14. [Google Scholar] [CrossRef]
- Caron, V.C. Estudo da Relação entre o Amolecimento Precoce de Mamões ‘Golden’e Aspectos Climáticos, Nutricionais e Hormonais. Ph.D. Thesis, Escola Superior de Agricultura Luiz de Queiroz, University of São Paulo, São Paulo, Brazil, 2014. [Google Scholar]
- Silva, M.D.S. Desempenho de Variedades de Mamoeiro em Clima Subtropical do Estado de São Paulo. Ph.D. Thesis, Faculdade de Ciências Agronômicas Universidade Estadual Paulista “Júlio de Mesquita Filho”, São Paulo, Brazil, 2016. [Google Scholar]
- Ortega-Farías, S.O.; Lozano, P.; Moreno, Y.; León, L. Desarrollo de modelos predictivos de fenología y evolución de madurez en vid para vino cv. Cabernet Sauvignon y Chardonnay. Agric. Técnica 2002, 62, 27–37. [Google Scholar] [CrossRef]
- Pailly, O.; Habib, R.; Delecolle, R. Effect of soil and climate conditions on soluble solids evolution during maturation of kiwifruit. N. Z. J. Crop. Hortic. Sci. 1995, 23, 145–153. [Google Scholar] [CrossRef]
- Ometto, J.C. Bioclimatologia Vegetal; Editora Agronômica Ceres LTDA: São Paulo, Brazil, 1981; 400p. [Google Scholar]
- Nakasone, H.Y. Produção de Mamão nos Trópicos e Subtrópicos. In Mamão; Ruggiero, C., Ed.; FCAV-UNESP: Jaboticabal, Brazil, 1988; pp. 19–42. [Google Scholar]
- Hoffman, G.J. Humidity. In Controlled Environment Guidelines for Plant Research; Tibbits, T.W., Kozlowski, T.T., Eds.; Academic Press: London, UK, 1979; pp. 141–172. [Google Scholar] [CrossRef]
- Marler, T.E.; Mickelbart, M.V. Drought. leaf gas exchange. and chlorophyll fluorescence of field-grown papaya. J. Am. Soc. Hortic. Sci. 1998, 123, 714–718. [Google Scholar] [CrossRef]
- Machado Filho, J.A.; Campostrini, E.; Yamanishi, O.K.; Fagundes, G.R. Variação sazonal das trocas gasosas em folhas de mamoeiro cultivado em condições de campo. Bragantia 2006, 65, 185–196. [Google Scholar] [CrossRef]
- Ritter, A.; Muñoz-Carpena, R. Predictive ability of hydrological models: Objective assessment of goodness-of-fit with statistical significance. J. Hydrol. 2013, 480, 33–45. [Google Scholar] [CrossRef]
- Knight, R.J. Origin and world importance of tropical and subtropical fruit crops. In Tropical and Subtropical Fruits: Composition, Properties, and Uses; Nagy, S., Shaw, P.E., Eds.; AVI Publishing: Westport, CT, USA, 1980; pp. 1–120. [Google Scholar] [CrossRef]
- Manica, I. Fruticultura Tropical: Mamão; Editora Agronômica Ceres LTDA: São Paulo, Brazil, 1982; 276p. [Google Scholar]
- Allan, P. Carica papaya responses under cool subtropical growth conditions. Acta Hortic. 2002, 575, 757–763. [Google Scholar] [CrossRef]
- Allan, P. Phenology and production of Carica papaya ‘honey gold’ under cool subtropical conditions. Acta Hortic. 2007, 740, 217–223. [Google Scholar] [CrossRef]
- Cabrera, J.A. Bases Tecnológicas para una Producción Eficiente y Sostenible de la Papaya (Carica papaya L.) en Climas Subtropicales. Ph.D. Thesis, Universidad de La Laguna, Tenerife, Spain, 2019. [Google Scholar]
- Reis, F.O. Microaspersão Sobrecopa em Mamoeiro ‘Golden’: Um Estudo Relacionado às Trocas Gasosas. à Eficiência Fotoquímica e ao Fluxo de Seiva Xilemática. Ph.D. Thesis, Campos do Goytacazes. Universidade Estadual do Norte Fluminense, Campos dos Goytacazes, RJ, Brazil, 2007. [Google Scholar]
- Reis, F.O.; Campostrini, E. Trocas gasosas e eficiência fotoquímica potencial em mamoeiro do grupo ‘Formosa’ cultivado em condição de campo. Bragantia 2008, 67, 815–822. [Google Scholar] [CrossRef]
- El-Sharkawy, M.A.; Cock, M.J.H.; Hernandez, A.D.P. Stomatal response to air humidity and its relation to stomatal density in a wide range of warm climate species. Photosynth. Res. 1985, 7, 137–149. [Google Scholar] [CrossRef]
- Salazar, R. Determination of photosynthesis in commercial varieties of papaw (Carica papaya L.) and its possible relationship with the production and quality of the fruits. Rev. ICA 1978, 13, 291–295. [Google Scholar]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology; Academic Press: San Diego, CA, USA, 1991; 635p. [Google Scholar] [CrossRef]
- Marschner, H. Mineral Nutrition of Higher Plants; Academic Press: San Diego, CA, USA, 1995; 889p. [Google Scholar] [CrossRef]
- Ooka, J. Oidium Caricae. Crop Knowledge Master, University of Hawai’i at Mānoa, College of Tropical Agriculture and Human Resources, 1993. Available online: http://www.extento.hawaii.edu/Kbase/crop/Type/o_caric.htm (accessed on 21 December 2020).
- Douglas, S.M. Powdery Mildew in the Greenhouse. Department of Plant Pathology and Ecology. The Connecticut Agricultural Experiment Station: New Haven, CT, USA. Available online: https://portal.ct.gov/CAES/Fact-Sheets/Plant-Pathology/Powdery-Mildew (accessed on 21 December 2020).
- Campostrini, E.; Yamanishi, O.K. Influence of mechanical root restriction on gas-exchange of four papaya genotypes. Braz. J. Plant Physiol. 2001, 13, 129–138. [Google Scholar] [CrossRef]
- Jeyakumar, P.; Kavino, M.; Kumar, N. Physiological performance of papaya cultivars under abiotic stress conditions. Acta Hortic. 2007, 740, 209–214. [Google Scholar] [CrossRef]
- Deng, X.; Ye, W.; Yang, Q.; Cai, C.; Liu, S. Effects of network-shaded planting model on photosynthetic. J. Beijing For. Univ. 2003, 26, 57–61. [Google Scholar]
- Arkle Junior, T.D.; Nakasone, H.Y. Floral differentiation in the hermaphroditic papaya. HortScience 1984, 19, 832–834. [Google Scholar]
- Ramos, H.C.C.; Pereira, M.G.; Silva, F.F.; Viana, A.P.; Ferreguetti, G.A. Seasonal and genetic influences on sex expression in a backcrossed segregating papaya population. Crop Breed. Appl. Biotechnol. 2011, 11, 97–105. [Google Scholar] [CrossRef][Green Version]
- Tamaki, M.; Urasaki, N.; Sunakawa, Y.; Motomura, K.; Adaniya, S. Seasonal variations in pollen germination ability. reproductive function of pistils. and seeds and fruit yield in papaya (Carica papaya L.) in Okinawa. J. Jpn. Soc. Hortic. Sci. 2011, 80, 156–163. [Google Scholar] [CrossRef]
- Salinas, I.; Hueso, J.J.; Cuevas, J. Flower development in ‘BH-65’ papaya grown in plastic greenhouses in south-east Spain. Acta Hortic. 2019, 1250, 187–192. [Google Scholar] [CrossRef]
- Zhou, L.; Christopher, D.A.; Paull, R. Defoliation and fruit removal effects on papaya fruit production. sugar accumulation. and sucrose metabolism. J. Am. Soc. Hortic. Sci. 2000, 125, 644–652. [Google Scholar] [CrossRef]
- Cabrera, J.A.; Raya, V.; Lobo, M.G.; Ritter, A. Effect of climate conditions on growth and production of hydroponic papaya crops in the Canary Islands. Acta Hortic. 2020, 1268, 77–84. [Google Scholar] [CrossRef]
- Jung Chen, N.; Manenoi, A.; Paull, R.E. Papaya postharvest physiology and handling problems and solutions. Acta Hortic. 2007, 740, 285–293. [Google Scholar] [CrossRef]
- McFadyen, L.M.; Hutton, R.J.; Barlow, W.R. Effects of crop load on fruit water relations and fruit growth in peach. J. Hortic. Sci. 1996, 71, 469–480. [Google Scholar] [CrossRef]
- Leonardi, C.; Guichard, S.; Bertin, N. High vapour pressure deficit influences growth. transpiration and quality of tomato fruits. Sci. Hortic. 2000, 84, 285–296. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | |||
|---|---|---|---|
| Variable | SP | NP | NN |
| FDD (days) | 180.42 ± 17.71 a | 219.09 ± 27.76 b | 244.14 ± 20.09 c |
| TSS (%) | 13.29 ± 0.60 c | 11.92 ± 0.94 b | 10.74 ± 1.41 a |
| GDD | 1368.36 ± 140.78 c | 1286.01 ± 117.92 b | 1196.51 ± 106.05 a |
| DPAR (mol m−2) | 4420.86 ± 592.99 a | 5450.63 ± 989.36 b | 6230.40 ± 1307.78 c |
| NF/LA (fruits m−2) | 3.07 ± 0.98 a | 5.75 ± 3.38 b | 8.61 ± 6.75 c |
| Season | |||||
|---|---|---|---|---|---|
| Variable | Location | Winter | Spring | Summer | Autumn |
| TSS | SP | 13.27 ± 0.70 b | 13.18 ± 0.56 c | 13.44 ± 0.33 b | 13.36 ± 0.51 a |
| NP | 11.09 ± 0.63 a | 11.45 ± 0.54 b | 12.76 ± 0.55 b | 12.71 ± 0.58 a | |
| NN | 11.30 ± 0.71 a | 8.41 ± 0.56 a | 11.33 ± 0.72 a | 12.30 ± 0.04 b | |
| NF/LA | SP | 2.95 ± 1.20 a | 3.00 ± 0.80 a | 4.06 ± 0.15 ab | 3.19 ± 0.76 a |
| NP | 7.29 ± 1.77 b | 9.33 ± 1.79 b | 2.30 ± 0.81 a | 2.74 ± 0.27 a | |
| NN | 9.90 ± 2.83 c | 19.48 ± 5.03 c | 4.64 ± 2.30 b | 5.78 ± 0.92 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabrera, J.A.; Ritter, A.; Raya, V.; Pérez, E.; Lobo, M.G. Papaya (Carica papaya L.) Phenology under Different Agronomic Conditions in the Subtropics. Agriculture 2021, 11, 173. https://doi.org/10.3390/agriculture11020173
Cabrera JA, Ritter A, Raya V, Pérez E, Lobo MG. Papaya (Carica papaya L.) Phenology under Different Agronomic Conditions in the Subtropics. Agriculture. 2021; 11(2):173. https://doi.org/10.3390/agriculture11020173
Chicago/Turabian StyleCabrera, Juan Alberto, Axel Ritter, Vanesa Raya, Eudaldo Pérez, and María Gloria Lobo. 2021. "Papaya (Carica papaya L.) Phenology under Different Agronomic Conditions in the Subtropics" Agriculture 11, no. 2: 173. https://doi.org/10.3390/agriculture11020173
APA StyleCabrera, J. A., Ritter, A., Raya, V., Pérez, E., & Lobo, M. G. (2021). Papaya (Carica papaya L.) Phenology under Different Agronomic Conditions in the Subtropics. Agriculture, 11(2), 173. https://doi.org/10.3390/agriculture11020173