Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities †



Abstract
1. Introduction
2. Materials and Methods
2.1. Inoculum Preparation
2.2. Greenhouse Rotation Trials
2.3. Field Set-Up and Design
2.4. Biocontrol Treatment Preparation
2.5. Disease and Yield Assessments
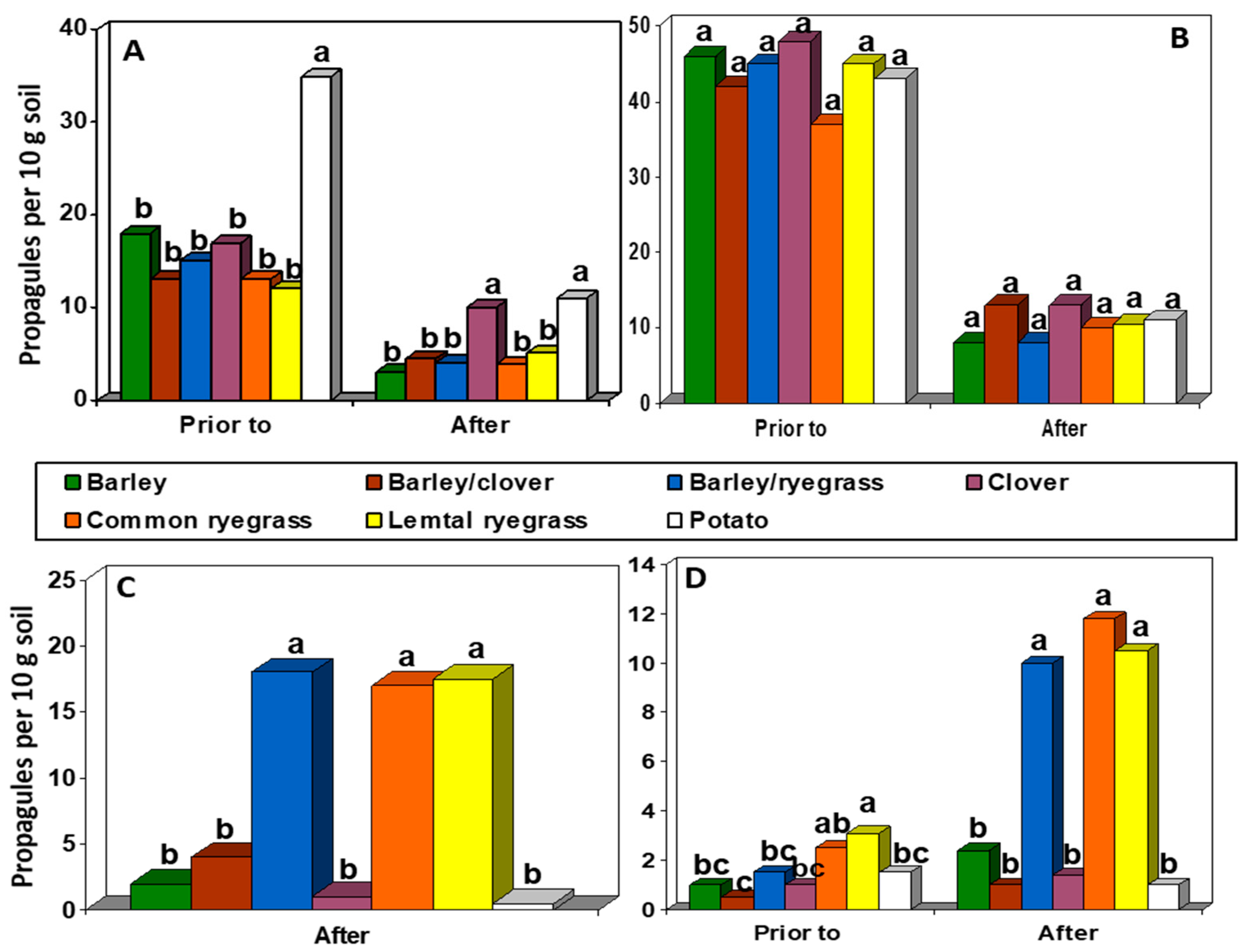
2.6. Soil Rhizoctonia Populations
2.7. Analysis of Microbial Communities Using FAME Profiles
2.8. Statistical Analysis
3. Results
3.1. Greenhouse Rotation Trials
3.2. Field Trial Emergence and Disease Ratings
3.3. Tuber Yield
3.4. Soil Rhizoctonia Populations
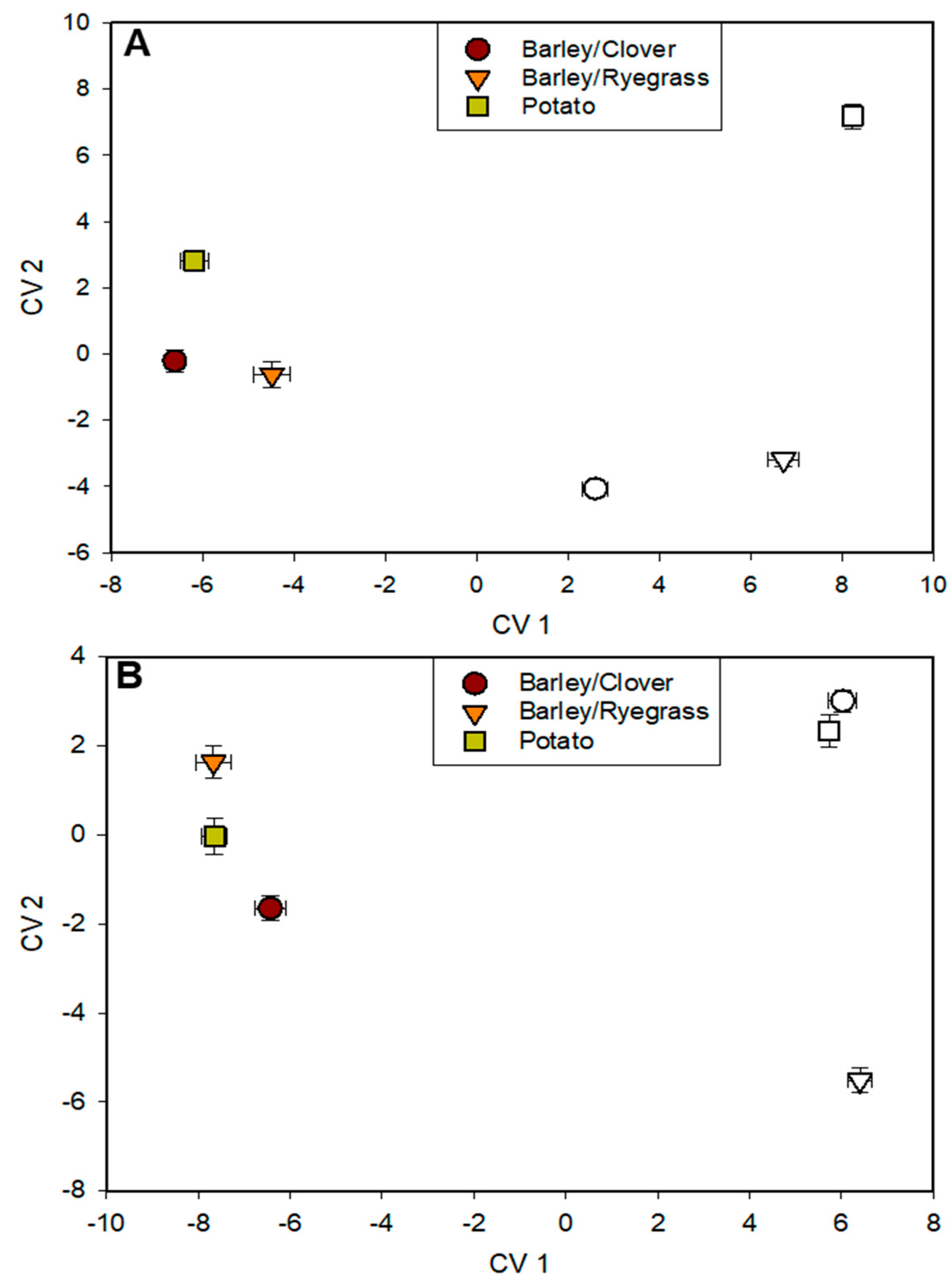
3.5. Analysis of Microbial Communities Using FAME Profiles
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsror, L. Biology, ecology and management of Rhizoctonia solani on potato. J. Phytopathol. 2010, 158, 649–658. [Google Scholar] [CrossRef]
- Powelson, M.L.; Rowe, R.C. Management of diseases caused by seedborne and soilborne fungi and fungus-like pathogens. In Potato Health Management, 2nd ed.; Johnson, D.A., Ed.; APS Press: St. Paul, MN, USA, 2008; pp. 149–158. [Google Scholar]
- Secor, G.A.; Gudmestad, N.C. Managing fungal diseases of potato. Can. J. Plant Pathol. 1999, 21, 213–221. [Google Scholar] [CrossRef]
- Stevenson, W.R.; Loria, R.; Franc, G.D.; Weingartner, D.P. (Eds.) Compendium of Potato Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 2001; 106p. [Google Scholar]
- Huber, D.M.; Sumner, D.R. Suppressive soil amendments for the control of Rhizoctonia species. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology, and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 433–443. [Google Scholar]
- Larkin, R.P.; Honeycutt, C.W. Effects of different 3-year cropping systems on soil microbial communities and soilborne disease of potato. Phytopathology 2006, 96, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.P. Soil health paradigms and implications for disease management. Ann. Rev. Phytopathol. 2015, 53, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Microbial diversity in soil: Selection of microbial populations by plant and soil type and implications for disease suppressiveness. Ann. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef]
- O’Donnell, A.G.; Seasman, M.; Macrae, A.; Waite, I.; Davies, J.T. Plants and fertilizers as drivers of change in microbial community structure. Plant Soil 2001, 232, 135–145. [Google Scholar] [CrossRef]
- Welbaum, G.E.; Sturz, A.V.; Dong, Z.; Nowak, J. Managing soil microorganisms to improve productivity of agro-ecosystems. Crit. Rev. Plant Sci. 2004, 23, 175–193. [Google Scholar] [CrossRef]
- Larkin, R.P.; Honeycutt, C.W.; Griffin, T.S.; Olanya, O.M.; Halloran, J.M.; He, Z. Effects of different potato cropping system approaches and water management on soilborne diseases and soil microbial communities. Phytopathology 2011, 101, 55–67. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S.; Honeycutt, C.W.; Olanya, O.M.; He, Z.; Halloran, J.M. Cumulative and residual effects of different potato cropping system management strategies on soilborne diseases and soil microbial communities over time. Plant Pathol. 2017, 66, 437–449. [Google Scholar] [CrossRef]
- Halloran, J.M.; Griffin, T.S.; Honeycutt, C.W. Economic analysis of potential rotation crops for Maine potato cropping systems. Am. J. Potato Res. 2005, 82, 155–162. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, G.S.; Honeycutt, C.W. Rotation and cover crop effects on soilborne potato diseases, tuber yield, and soil microbial communities. Plant Dis. 2010, 94, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Dabney, S.M.; Schreiber, J.D.; Rothrock, C.S.; Johnson, J.R. Cover crops affect sorghum seedling growth. Agron. J. 1996, 88, 961–970. [Google Scholar] [CrossRef]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani damping-off of tomato with Bacillus subtilus RB14. Appl. Environ. Microbiol. 1996, 62, 397–404. [Google Scholar] [CrossRef]
- Harris, A.R.; Adkins, P.G. Versatility of fungal and bacterial isolates for biological control of damping-off disease caused by Rhizoctonia solani and Pythium spp. Biol. Control 1999, 15, 10–18. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, N.; Yong, X.; Yang, X.; Shen, Q. Biocontrol of Rhizoctonia solani damping-off disease in cucumber with Bacillus pumilus SQR-N43. Microbiol. Res. 2012, 167, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Hijri, M. Screening, identification and evaluation of potential biocontrol fungal endophytes against Rhizoctonia solani AG3 on potato plants. FEMS Microbiol. Lett. 2010, 311, 152–159. [Google Scholar] [CrossRef]
- Murdoch, C.W.; Leach, S.S. Evaluation of Laetisaria arvalis as a biological control agent of Rhizoctonia solani on white potato. Am. Potato J. 1993, 70, 625–633. [Google Scholar] [CrossRef]
- Tsror, L.; Barak, R.; Sneh, B. Biological control of black scurf on potato under organic management. Crop Prot. 2001, 20, 145–150. [Google Scholar] [CrossRef]
- Brewer, M.T.; Larkin, R.P. Efficacy of several potential biocontrol organisms against Rhizoctonia solani on potato. Crop Prot. 2005, 24, 939–950. [Google Scholar] [CrossRef]
- Larkin, R.P.; Tavantzis, S. Use of biocontrol organisms and compost amendments for improved control of soilborne diseases and increased potato production. Am. J. Pot. Res. 2013, 90, 261–270. [Google Scholar] [CrossRef]
- Larkin, R.P. Impacts of biocontrol products on Rhizoctonia disease of potato and soil microbial communities, and their persistence in soil. Crop Prot. 2016, 90, 96–105. [Google Scholar] [CrossRef]
- Bernard, E.; Larkin, R.P.; Tavantzis, S.; Erich, M.S.; Alyokhin, A.; Gross, S. Rapeseed rotation, compost and biocontrol amendments reduce soilborne diseases and increase tuber yield in organic and conventional potato production systems. Plant Soil 2014, 374, 611–627. [Google Scholar] [CrossRef]
- Van Bruggen, A.H.C.; Grünwald, N.J.; Bolda, M. Cultural methods and soil nutrient status in low and high input agricultural systems, as they affect Rhizoctonia species. In Rhizoctonia Species: Taxonomy, Molecular Biology, Ecology, Pathology, and Disease Control; Sneh, B., Jabaji-Hare, S., Neate, S., Dijst, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1996; pp. 407–421. [Google Scholar]
- Larkin, R.P. Characterization of soil microbial communities under different potato cropping systems by microbial population dynamics, substrate utilization, and fatty acid profiles. Soil Biol. Biochem. 2003, 35, 1451–1466. [Google Scholar] [CrossRef]
- Larkin, R.P. Relative effects of biological amendments and crop rotations on soil microbial communities and soilborne diseases of potato. Soil Biol. Biochem. 2008, 40, 1341–1351. [Google Scholar] [CrossRef]
- Han, J.S.; Cheung, J.H.; Yoon, T.M.; Song, J.; Rajkarnakar, A.; Kim, W.G.; Yoo, I.D.; Yang, Y.Y.; Suh, J.W. Biological control agent of common scab disease by antagonistic strain Bacillus sp. sunhua. J. Appl. Microbiol. 2005, 99, 213–221. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Zhuang, L.; Yu, Y.; Liu, J.; Zhang, L.; Gao, Z.; Wu, Y.; Gao, W.; Ding, G.; et al. A rhizosphere-derived consortium of Bacillus subtilis and Trichoderma harzianum suppresses common scab of potato and increases yield. Comput. Struct. Biotechnol. J. 2019, 17, 645–653. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W.; Schliefer, K.H. Phylogenteic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar]
- Cavigelli, M.A.; Robertson, G.P.; Klug, M.J. Fatty acid methyl ester (FAME) profiles as measures of soil microbial community structure. Plant Soil 1995, 170, 99–113. [Google Scholar] [CrossRef]
- Ritchie, N.J.; Schutter, M.E.; Dick, R.P.; Myrold, D.D. Use and length of heterogeneity PCR and fatty acid methyl ester profiles to characterize microbial communities in soil. Appl. Environ. Microbiol. 2000, 66, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, S.D.; Germida, J.J. Biolog analysis and fatty acid methyl ester profiles indicate that Pseudomonad inoculants that promote phytorediation alter the root associated microbial community of Bromus biebersteinii. Soil Biol. Biochem. 1998, 30, 1717–1723. [Google Scholar] [CrossRef]
- Essah, S.Y.C.; Honeycutt, C.W. Tillage and seed-sprouting strategies to improve potato yield and quality in short season climates. Am. J. Potato Res. 2004, 81, 177–186. [Google Scholar] [CrossRef]
- Flowers, R.A. A selective media for isolation of Rhizoctonia from soil and plant tissue. Proc. Am. Phytopathol. Soc. 1976, 3, 219. [Google Scholar]
- Henis, Y.; Ghaffar, A.; Baker, R.; Gillespie, S.L. A new soil-sampler and its use for the study of population dynamics of Rhizoctonia solani in soil. Phytopathology 1978, 68, 371–376. [Google Scholar] [CrossRef]
- Sneh, B.; Katan, J.; Henis, Y.; Wahl, I. Methods for evaluating inoculum density of Rhizoctonia in naturally infested soil. Phytopathology 1966, 56, 74–78. [Google Scholar]
- Buyer, J.S.; Roberts, D.P.; Russek-Cohen, E. Microbial community structure and function in the spermosphere as affected by soil and seed type. Can. J. Microbiol. 1999, 45, 138–144. [Google Scholar] [CrossRef]
- Sneh, B.; Burpee, L.; Ogoshi, A. Identification of Rhizoctonia Species; APS Publishing: St. Paul, MN, USA, 1991. [Google Scholar]
- Read, P.J.; Hide, G.A.; Firmager, J.P.; Hall, S.M. Growth and yield of potatoes as affected by severity of stem canker (Rhizocrtonia solani). Potato Res. 1989, 32, 9–15. [Google Scholar] [CrossRef]
- Hubbard, R.K.; Strickland, T.C.; Phatak, S. Effects of cover crop systems on soil physical properties and carbon/nitrogen relationships in the coastal plain of southeastern USA. Soil Till. Res. 2013, 126, 276–283. [Google Scholar] [CrossRef]
- Schutter, M.E.; Dick, R.P. Microbial community profiles and activities among aggregates of winter fallow and cover-cropped soil. Soil Sci. Soc. Am. J. 2002, 66, 142–153. [Google Scholar] [CrossRef]
- Johnston, H.W.; Celetti, M.J.; Kimpinski, J.; Platt, H.W. Fungal pathogens and Pratylenchus penetrans associated with preceding crops of clover, winter wheat, and annual ryegrass and their influence on succeeding potato crops on Prince Edward Island. Am. Potato J. 1994, 71, 797–808. [Google Scholar] [CrossRef]
- Carter, M.R.; Kunelius, H.T.; Sanderson, J.B.; Kimpinski, J.; Platt, H.W.; Bolinder, M.A. Productivity parameters and soil health dynamics under long-term 2-year potato rotations in Atlantic Canada. Soil Till. Res. 2003, 72, 152–168. [Google Scholar] [CrossRef]
- Burpee, L.; Martin, B. Biology of Rhizoctonia species associated with turfgrasses. Plant Dis. 1992, 76, 112–117. [Google Scholar] [CrossRef]
- Martin, S.B.; Lucas, L.T. Pathogenicity of Rhizoctonia zeae on tall fescue and other turfgrasses. Plant Dis. 1983, 67, 676–678. [Google Scholar] [CrossRef]
- Carling, D.E.; Leiner, R.H. Effect of temperature on virulence of Rhizoctonia solani and other Rhizoctonia on potato. Phytopathology 1990, 80, 930–934. [Google Scholar] [CrossRef]
- Webb, K.M.; Harveson, R.M.; West, M.S. Evaluation of Rhizoctonia zeae as a potential biological control option for fungal root diseases of sugar beet. Ann. Appl. Biol. 2015, 167, 75–89. [Google Scholar] [CrossRef]
- Carter, M.R.; Sanderson, J.B. Influence of conservation tillage and rotation length on potato productivity, tuber disease and soil quality parameters on a fine sandy loam in eastern Canada. Soil Till. Res. 2001, 63, 1–13. [Google Scholar] [CrossRef]
- Larkin, R.P. Incorporating disease-suppressive rotation crops and organic amendments into improved potato cropping systems. Phytopathology 2017, 107. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142. [Google Scholar] [CrossRef]
- Zelles, L.; Bai, Q.Y.; Beck, T.; Beese, F. Signature fatty acids in phospholipids and lipopolysaccharides as indicators of microbial biomass and community structure in agricultural soils. Soil Biol. Biochem. 1992, 24, 317–323. [Google Scholar] [CrossRef]
- Bossio, D.A.; Scow, K.M.; Gunapala, N.; Graham, K.J. Determinants of soil microbial communities: Effects of agricultural management, season, and soil type on phospholipid fatty acid profiles. Microb. Ecol. 1998, 36, 1–12. [Google Scholar] [CrossRef]
- Zelles, L.; Bai, Q.Y.; Rackwitz, R.; Chadwick, D.; Beese, F. Determination of phospholipid- and lipopolysaccaride-derived fatty acids as an estimate of microbial biomass and community structures in soil. Biol. Fertil. Soils 1995, 19, 115–123. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Parke, J.L.; Clayton, M.K.; Handelsman, J. Effects of an introduced bacterium on bacterial communities on roots. Ecology 1993, 74, 840–854. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. with special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef] [PubMed]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rotation Crop | Discolored Stem Incidence w (%) | Stem Canker Incidence x (%) | Stem Canker Severity (0–5) w | Shoot Size y |
|---|---|---|---|---|
| Experiment 1 | ||||
| Barley | 68.8 ab z | 0.0 c | 0.69 ab | 2.31 abc |
| Barley/clover | 62.5 b | 6.3 ab | 0.69 ab | 2.56 a |
| Barley/ryegrass | 87.5 ab | 12.5 bc | 1.00 ab | 2.37 ab |
| Clover | 68.8 ab | 37.5 a | 1.31 a | 1.93 c |
| Common ryegrass | 56.3 b | 12.5 bc | 1.06 ab | 2.44 a |
| Lemtal ryegrass | 87.5 ab | 18.8 ab | 0.69 b | 2.44 a |
| Potato | 93.8 a | 25.0 ab | 1.25 a | 2.00 bc |
| Experiment 2 | ||||
| Barley | 93.8 ab | 39.5 ab | 1.50 ab | 2.33 ab |
| Barley/clover | 87.5 ab | 31.3 ab | 1.52 ab | 2.62 a |
| Barley/ryegrass | 93.8 ab | 18.8 b | 1.12 bc | 2.73 a |
| Clover | 91.8 ab | 43.8 a | 1.79 a | 2.15 b |
| Common ryegrass | 83.3 ab | 14.5 b | 1.00 c | 2.38 ab |
| Lemtal ryegrass | 68.8 b | 33.3 ab | 1.08 bc | 2.69 a |
| Potato | 100.0 a | 37.5 ab | 1.73 a | 2.71 a |
| Shoot Emergence w (%) | Stem Canker | ||
|---|---|---|---|
| Severity x | Incidence y (%) | ||
| Preceding Rotation Crop (Mainplots) | |||
| Barley/ryegrass | 75.4 a z | 1.33 b | 25.0 b |
| Barley/clover | 79.3 a | 1.45 ab | 39.1 b |
| Potato | 84.2 a | 1.70 a | 59.4 a |
| Biocontrol Treatment (Subplots) | |||
| Laetisaria arvalis | 83.9 a | 1.53 a | 45.8 a |
| Trichoderma virens | 76.7 b | 1.56 a | 45.8 a |
| Bacillus subtilis | 77.6 ab | 1.46 a | 37.5 a |
| Control | 80.4 ab | 1.52 a | 35.4 a |
| Rotation Crop | Severity x | Incidence (%) | Incidence ≥ 2 y (%) |
|---|---|---|---|
| Biocontrol Treatment | |||
| Barley/Ryegrass | |||
| Laetisaria arvalis | 1.68 c z | 64.9 b | 43.3 c |
| Trichoderma virens | 2.21 b | 87.3 a | 55.8 b |
| Bacillus subtilis | 1.96 bc | 84.3 a | 43.4 c |
| Control | 2.69 a | 85.8 a | 71.8 a |
| Barley/Clover | |||
| Laetisaria arvalis | 1.76 a | 74.1 a | 49.9 a |
| Trichoderma virens | 1.02 b | 52.4 b | 25.8 b |
| Bacillus subtilis | 1.83 a | 70.8 a | 45.8 a |
| Control | 1.74 a | 83.4 a | 49.1 a |
| Potato | |||
| Laetisaria arvalis | 1.81 a | 75.8 a | 60.1 a |
| Trichoderma virens | 1.77 a | 79.0 a | 56.6 a |
| Bacillus subtilis | 1.98 a | 75.1 a | 57.5 a |
| Control | 1.80 a | 78.3 a | 60.0 a |
| Yield (Mg/ha) | ||||||
|---|---|---|---|---|---|---|
| Small v | Medium w | Large x | Total | Marketable y | Misshapen | |
| Location 1 (Central Maine) | ||||||
| Preceding Rotation Crop (Mainplots) | ||||||
| Barley/ryegrass | 0.87 b z | 2.30 b | 14.1 a | 17.3 a | 16.4 a | 1.31 a |
| Barley/clover | 0.92 b | 2.03 b | 12.4 ab | 15.4 ab | 14.5 ab | 1.58 a |
| Potato | 1.54 a | 3.05 a | 10.1 b | 14.7 b | 13.2 b | 1.62 a |
| Biocontrol Treatment (Subplots) | ||||||
| Laetisaria arvalis | 1.05 a | 2.64 a | 12.2 a | 15.9 a | 14.9 a | 1.45 a |
| Trichoderma virens | 1.19 a | 2.17 a | 12.1 a | 15.4 a | 14.3 a | 1.31 a |
| Bacillus subtilis | 1.01 a | 2.60 a | 12.4 a | 16.0 a | 15.0 a | 1.52 a |
| Control | 1.19 a | 2.44 a | 12.2 a | 15.8 a | 14.6 a | 1.74 a |
| Location 2 (Northern Maine) | ||||||
| Preceding Rotation Crop (Mainplots) | ||||||
| Barley/ryegrass | 1.72 a | 4.32 a | 10.6 a | 16.68 a | 15.0 a | 0.75 a |
| Barley/clover | 1.87 a | 4.27 a | 12.4 a | 18.51 a | 16.6 a | 0.80 a |
| Potato | 1.74 a | 5.50 a | 12.3 a | 19.55 a | 17.8 a | 0.78 a |
| Biocontrol Treatment (Subplots) | ||||||
| Laetisaria arvalis | 1.39 b | 4.09 b | 12.9 a | 18.4 a | 17.0 a | 0.66 a |
| Trichoderma virens | 1.46 b | 3.90 b | 12.3 ab | 17.7 a | 16.2 a | 0.87 a |
| Bacillus subtilis | 1.92 ab | 4.98 ab | 12.3 ab | 19.2 a | 17.3 a | 0.90 a |
| Control | 2.34 a | 5.82 a | 9.5 b | 17.6 a | 15.3 a | 0.67 a |
| Fatty Acid Classes (%) x | Class Ratios y | |||||||
|---|---|---|---|---|---|---|---|---|
| Sat | Mono | Poly | Branch | Hydrox | Mono/Sat | Gram+/Gram− | Fungi/Bact | |
| Location 1 (Central Maine) | ||||||||
| After Growth | ||||||||
| Barley/ryegrass | 32.5 b z | 43.0 a | 10.6 a | 20.8 b | 11.0 b | 1.33 a | 1.94 a | 0.34 a |
| Barley/clover | 32.9 b | 41.4 a | 11.0 a | 20.0 b | 11.9 ab | 1.27 a | 1.71 a | 0.35 a |
| Potato | 38.0 a | 42.1 a | 8.5 b | 23.2 a | 13.3 a | 1.11 b | 1.76 a | 0.24 b |
| After Incorporation | ||||||||
| Barley/ryegrass | 30.7 ab | 41.9 a | 12.4 a | 18.5 ab | 9.4 a | 1.37 a | 1.99 a | 0.45 a |
| Barley/clover | 30.1 b | 39.2 b | 11.9 a | 18.2 b | 10.6 a | 1.31 a | 1.78 a | 0.42 a |
| Potato | 32.3 a | 38.5 b | 12.0 a | 19.6 a | 10.0 a | 1.20 b | 2.02 a | 0.41 a |
| Location 2 (Northern Maine) | ||||||||
| After Growth | ||||||||
| Barley/ryegrass | 20.0 a | 22.5 a | 6.5 ab | 8.8 b | 12.5 a | 1.14 a | 0.72 b | 0.31 a |
| Barley/clover | 20.0 a | 22.3 a | 7.3 a | 9.4 b | 12.3 a | 1.12 a | 0.84 b | 0.35 a |
| Potato | 20.5 a | 22.5 a | 5.1 b | 10.4 a | 10.6 a | 1.10 a | 1.04 a | 0.25 a |
| After Incorporation | ||||||||
| Barley/ryegrass | 18.6 a | 24.7 a | 7.5 a | 8.9 a | 12.8 a | 1.36 a | 0.83 a | 0.37 a |
| Barley/clover | 19.8 a | 23.1 ab | 7.1 a | 9.1 a | 11.1 a | 1.18 ab | 0.89 a | 0.36 ab |
| Potato | 19.8 a | 21.3 b | 6.0 b | 9.2 a | 12.3 a | 1.09 b | 0.81 a | 0.28 b |
| Treatment | Fatty Acid Classes (%) x | Class Ratios y | ||||||
|---|---|---|---|---|---|---|---|---|
| Sat | Mono | Poly | Branch | Hydrox | Mono/Sat | Gram+/Gram− | Fungi/Bact | |
| Location 1 (Central Maine) | ||||||||
| Barley/Ryegrass | ||||||||
| L. arvalis | 26.3 a z | 26.9 a | 7.6 a | 14.5 a | 6.3 a | 1.04 a | 2.39 a | 0.37 ab |
| T. virens | 24.6 a | 26.8 a | 8.8 a | 13.1 b | 5.9 a | 1.11 a | 2.38 a | 0.47 a |
| B. subtilis | 26.7 a | 26.2 a | 6.9 a | 13.0 b | 6.3 a | 1.00 a | 2.24 a | 0.36 ab |
| control | 26.8 a | 26.7 a | 7.2 a | 13.6 ab | 7.3 a | 1.01 a | 2.02 a | 0.35 b |
| Barley/Clover | ||||||||
| L arvalis | 26.3 ab | 26.6 ab | 7.7 a | 14.0 a | 6.7 b | 1.02 b | 2.14 a | 0.38 a |
| T. virens | 24.5 b | 27.8 a | 7.8 a | 14.1 a | 6.2 b | 1.13 a | 2.32 a | 0.39 a |
| B. subtilis | 27.4 a | 25.9 b | 6.4 b | 13.8 a | 7.8 a | 0.98 b | 1.84 b | 0.30 b |
| control | 26.4 ab | 27.2 ab | 7.9 a | 13.9 a | 6.1 b | 1.04 ab | 2.38 a | 0.40 a |
| Potato | ||||||||
| L. arvalis | 29.4 a | 24.9 a | 5.2 b | 14.5 ab | 7.8 a | 0.88 a | 1.96 a | 0.24 b |
| T.virens | 27.9 ab | 26.6 a | 6.2 ab | 14.2 b | 7.9 a | 0.98 a | 2.05 a | 0.29 ab |
| B.subtilis | 26.6 ab | 25.0 a | 6.9 a | 15.5 a | 6.8 a | 0.96 a | 2.38 a | 0.34 a |
| control | 25.6 b | 24.6 a | 6.7 ab | 14.6 ab | 7.4 a | 0.97 a | 2.09 a | 0.32 ab |
| Location 2 (Northern Maine) | ||||||||
| Preceding Rotation Crop | ||||||||
| Barley/ryegrass | 29.7 a | 29.6 a | 24.1 a | 15.7 a | 13.8 b | 1.00 a | 1.18 a | 0.82 a |
| Barley/clover | 30.2 a | 29.4 a | 23.2 a | 16.0 a | 16.1 a | 0.98 a | 1.04 a | 0.73 b |
| Potato | 30.8 a | 28.7 a | 22.8 a | 15.5 a | 15.7 a | 0.93 b | 1.18 a | 0.74 b |
| Biocontrol Treatment | ||||||||
| L. arvalis | 30.7 a | 29.2 a | 23.5 ab | 15.5 b | 16.2 a | 0.96 a | 1.01 b | 0.75 a |
| T.virens | 30.1 a | 29.4 a | 23.9 a | 15.5 b | 15.3 ab | 0.98 a | 1.08 ab | 0.78 a |
| B.subtilis | 30.3 a | 29.2 a | 23.4 ab | 15.6 ab | 14.5 b | 0.97 a | 1.14 ab | 0.78 a |
| control | 30.0 a | 29.1 a | 22.9 b | 16.2 a | 14.8 ab | 0.98 a | 1.16 a | 0.74 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larkin, R.P.; Brewer, M.T. Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities. Agriculture 2020, 10, 128. https://doi.org/10.3390/agriculture10040128
Larkin RP, Brewer MT. Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities. Agriculture. 2020; 10(4):128. https://doi.org/10.3390/agriculture10040128
Chicago/Turabian StyleLarkin, Robert P., and Marin T. Brewer. 2020. "Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities" Agriculture 10, no. 4: 128. https://doi.org/10.3390/agriculture10040128
APA StyleLarkin, R. P., & Brewer, M. T. (2020). Effects of Crop Rotation and Biocontrol Amendments on Rhizoctonia Disease of Potato and Soil Microbial Communities. Agriculture, 10(4), 128. https://doi.org/10.3390/agriculture10040128