1. Introduction
The kiwifruit industry in Italy currently represents one of the most dynamic and profitable investments within the Italian agricultural sector. Kiwifruit production and plantation area have been increasing steadily since the 1970s, when the first orchards were planted. By now, the production of Italian kiwifruit is concentrated in the following districts: the province of Latina, the Romagna region, and the provinces of Cuneo and Verona [
1]. However, the kiwifruit industry has experienced serious problems in recent times: since the 2007–2008 seasons,
Pseudomonas syringae pv.
actinidiae epidemics have caused major economic losses [
2], while more recently, the introduction of the marmorated stinkbug (
Halyomorpha halys) has seriously compromised fruit quality and marketable yields [
3]. Finally, from 2012–2013, kiwifruit orchards in northern Italy showed a rapid vine decline especially in those areas, such as Verona province, where the common irrigation practice consists of temporary flooding [
4,
5].
Initially, symptoms were reported on a surface of about 600 ha [
6] and since then, kiwifruit vine decline syndrome (KVDS) has spread to the most important Italian regions for kiwifruit production (e.g., Piedmont, Friuli-Venezia Giulia, Lazio). Nowadays, KVDS affects almost 3000 ha of kiwifruit orchards, equal to 10% of the total area cultivated to kiwifruit in Italy [
7]. Symptoms similar to KVDS have been observed in different environments and were often associated either to waterlogging, root rotting or root asphyxia [
8,
9,
10,
11,
12]. In addition, KVDS development is usually associated with excessive precipitation/irrigation both in spring and in summer [
5]. In fact, the peak of symptomatology generally occurs in mid-late summer, in concomitance with high temperatures, when transpiration of the vines is highest and shoots support the highly-demanding fruit growth [
8]. Among the earliest symptoms, a progressive desiccation of leaves moving from leaf margin to inner tissues is observed. Leaf margin deformation and sprout desiccation are other common symptoms especially in 1- to 2-year old vines. With the progression of symptoms, the majority of the canopy shows leaf margin desiccation and, at this stage, extensive phylloptosis occurs, triggering the plant’s collapse. In symptomatic vines, a diffuse fruit drop may take place, while remaining berries are impaired in their development, showing a stunted growth often associated to skin wrinkling at harvest [
6]. Generally, once affected, vines are expected to die within 2 years [
6].
The symptomatology on the canopy reflects the massive damages to the root system. In comparison to healthy plants, the root apparatus in suffering vines is characterised by the scarcity or absence of feeder roots, while the structural roots show widespread brown, soft-rotting areas. With the progression of KVDS, the root cortical layers may show hypertrophy, followed by phloem detachment from the central cylinder, thus causing the breakdown of the cortex. The development of symptoms on the canopy may remain latent, even though the root system is already compromised.
Several abiotic [
6,
7,
13,
14] and biotic [
5,
6] factors have been suggested to trigger the onset of KVDS. Water pollution and salinity, accumulation of heavy metals, xenobiotic molecules or other contaminants in the soil, presence of nematodes and soil structure anomalies (i.e., compaction or clay aggregation) have been suggested to promote KVDS symptoms and diffusion. However, none of these factors was sufficient, per se, to reproduce KVDS symptoms [
5,
6]. Moreover, even though these factors enhance KVDS severity, no statistical correlation was observed among these factors and KVDS incidence [
5,
6].
Among the abiotic factors, waterlogging caused by either excessive irrigation or precipitation was shown to be the main driver of KVDS [
7,
13,
14]. Kiwifruit plants are highly sensitive to waterlogging conditions [
9,
10,
11]. For example, massive plant death was reported in the late 1980s in New Zealand after the passing of the cyclones Delilah and Bola, when the strong rainfalls caused soil water saturation for some days [
11,
12]. The strong sensitivity of kiwifruit plants to waterlogging is due to the high consumption of oxygen by the root system. In fact, an adult kiwifruit plant consumes up to 4.6 × 10
−6 mol/m
3 of oxygen, and thus it is able to deplete the oxygen reserves of a well-aerated soil in just 5 h [
15]. Furthermore, the root asphyxia symptoms can be worsened by the limited gas circulation within the root, due to the small intercellular space between contiguous cells [
15,
16].
After a few hours of induced asphyxia, vines experience a strong reduction in root activity, as well as in stomata conductance. Indeed, the lack of oxygen at root level induces a reduction in the stomata conductance, starting when just 25% of the roots are affected for at least 48 h [
15]. In these conditions, root-to-canopy water flow is suppressed, and leaves and shoots may suffer from water scarcity although the soil water content is high. Additionally, leaves cannot enact thermoregulation mechanisms against sun burn damages [
16], which cause symptoms resembling the initial steps of KVDS. Excessive irrigation induces also soil compaction and reduction in soil porosity, which is further hampered in soils with a scarce organic matter fraction content.
Field observations suggest that pathogenic microorganisms may also be involved in KVDS. In fact, affected plants are initially located in a limited area of the orchard, and symptoms seemingly spread from affected vines to the surrounding ones [
5]. Moreover, preliminary trials performed by transplanting young vines in pots filled with soil collected in the proximity of affected plants, allowed to reproduce KVDS symptoms which were observed after reiterate cycles of excessive root watering (simulating flooding conditions). Finally, soil sterilisation abolished symptom development, indicating the potential involvement of biotic factors [
7].
Different species of pathogenic fungi have been suggested to be involved, at least indirectly, in the development of the decline syndrome [
17,
18]. In fact, even after soil conditions were restored to optimal level, by increasing aeration and drainage, plants were still suffering from the decline syndrome.
Phytophthora megasperma was isolated from these plants and, in experimental conditions, this pathogen was able to induce root necrosis even under mildly unfavourable conditions [
17]. Species belonging to
Phytophthora-related genera (
Pythium,
Phytopythium) and
Armillaria spp. were reported to affect kiwifruit plants [
15,
19,
20,
21,
22,
23]. Nonetheless, their involvement in KVDS occurrence has not been demonstrated.
In this context, we investigated the potential involvement of the root-associated microbial community in the onset of KVDS. For this purpose, the bacterial and fungal community of roots of healthy and KVDS-affected kiwifruit vines was characterised with a classical microbiological approach. Finally, inoculation trials were performed in controlled conditions on potted plants subjected to different irrigation regimes to verify whether specific fungal isolates could reproduce KVDS symptomatology.
2. Materials and Methods
2.1. Microbial Isolation
Root samples of kiwifruit vines [
A. chinensis var.
deliciosa (A.Chev.) A.Chev. cv. ‘Hayward’] were collected from three mature, commercial orchards located in the Verona province (North Italy) characterised by different KVDS incidence. Orchard 1 (O1) had less than 5% of the plants suffering KVDS symptoms, orchard 2 (O2) was heavily affected by the syndrome, with most of the plants (approx. 60%) affected by serious canopy wilting; while orchard 3 (O3) did not show any visible symptoms of KVDS. A plant was considered symptomatic when at least 25% of the shoots showed symptoms of wilting or desiccations, or alternatively when the percentage of chlorotic/necrotic leaves were over 30% of the foliage (
Figure S1).
Roots (4 vines per orchard with 5 replicates per vine) were sampled, washed in a 0.05% Tween 80 solution then rinsed with deionised water. The root surface was disinfested for 1 min in a 1.5% sodium hypochlorite (NaOCl) solution and then rinsed twice with deionised water. After surface sterilisation, samples were individually divided into: (1) radicle, (2) secondary root cortex and (3) stele.
Concerning fungal isolation, small pieces of root tissue (15 mm) were placed on potato dextrose agar (PDA) amended with 0.2 g L−1 rifampicin and 0.5 g L−1 tetracycline to prevent bacterial growth. Plates were incubated for 5–10 days at 24 ± 1 °C in the dark. A pure culture of each isolate was obtained prior to morphological and molecular identification. For long-term storage, cultures were kept on PDA slants or dishes at 4 °C. The morphology of each isolate was determined on PDA medium. Mycelial samples (5 mm diameter) were taken from the edge of 5-day-old colonies and inoculated in the centre of a new PDA plate. Colony characteristics (colour, aerial mycelium, margin shape, presence and morphology of reproductive structures) of isolates were determined after 7–10 days of incubation in 12 h light/12 h dark cycle at 24 ± 1 °C.
For bacterial isolation, small pieces of the root (15 mm) were collected and grinded in a sterile 10 mM MgSO4 solution. The obtained suspension was filtered through a sterile gauze (100 µm mesh) to discard plant debris. Samples were serially diluted and plated on lysogenic broth (LB), nutrient agar (NA) and King’s B medium (KB), and incubated under microaerophilic conditions at 30 °C for 48 h. The different growing media were amended with cycloheximide (10 mg L−1) to prevent fungal contamination. All colonies with a distinct phenotype (colour, form, margin and elevation) were enumerated and collected from the plates at the highest countable dilution. The colony selection from the highest dilution plates allowed us to collect the most frequent and predominant species in each sample. Isolates were stored at −80 °C in LB broth supplemented with glycerol (20% v/v) (Sigma-Aldrich Srl., Milan, Italy) or on LB agar at 4 °C for short-term storage.
2.2. DNA Extraction and Molecular Identification
Fungal mycelium was collected from pure cultures grown on PDA after 7–10 days of incubation and suspended in 1 mL sterile water. The suspension was centrifuged at 10,000 rpm for 5 min. Pellets were re-suspended in 200 μL of InstaGeneTM Matrix (BioRad, Milan, Italy) and sonicated for 15 min in an ultrasonic bath. Subsequently, the samples were incubated, first at 56 °C for 30 min and then at 100 °C for 10 min. At the end of the incubation steps, samples were centrifuged at 10,000 rpm for 10 min and the supernatants stored at −20 °C.
Fungal isolates were subjected to the amplification of the ITS1-5.8S-ITS2 (Internal Transcribed Spacer) region using the following primer pair: ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′); ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). Amplification was performed in 25 µL total volume, with 0.5 µM of each primer, in agreement with Esteve-Zarzoso et al. [
24]. The amplification programme included an initial denaturation step at 94 °C for 5 min, 30 cycles of 94 °C for 30 s, 55 °C for 30 s, 72 °C for 1 min, and a final step of 72 °C for 10 min.
For bacterial isolates, DNA was extracted using Bacterial DNA isolation kit (Sigma-Aldrich, Milan, Italy) according to manufacturer’s instructions. 16S rRNA gene amplification was performed as previously described [
25], using Lac16S-for (5′-AATGAGAGTTTGATCCTGGCT-3′) and Lac16Srev (5′-GAGGTGATCCAGCCGCAGGTT-3′) primers.
The amplified fragments were then purified using GFX PCR DNA and Gel Band Purification Kit (Amersham Biosciences, Uppsala, Sweden), according to the manufacturer’s instructions, dried, and sequenced by the BMR Genomics Labs (Padova, Italy). Sequence chromatograms were obtained through Chromas software (Technelysium Pty Ltd, Brisbane, Australia) and can be accessed at bmr.cribi.unipd.it. Sequences were compared to those available in the GenBank database (
http://www.ncbi.nml.nih.gov/BLAST) and in the Ribosomal Database Project (
http://rdp.cme.msu.edu/index.jsp) to determine the closest known relative species.
2.3. Inoculation Trials in Greenhouse Conditions
Inoculation experiments were performed in controlled conditions in the greenhouse of the University of Bologna on potted kiwifruit vines to verify the ability of different isolates to trigger the typical symptoms of the syndrome once inoculated on healthy vines. The plants were cultivated in 10 litre plastic pots, each filled with an equal quantity of a potting mix composed of 40% brown peat, 10% pine bark and 50% disinfected soil.
Since no bacterial pathogens were isolated from analysed samples, only fungal species were investigated for the inoculation trials. Pathogenicity assay was performed on fifty-six 2-year-old A. macrosperma (cv. Bounty 71) potted vines, commonly adopted as a rootstock for both A. chinensis var. chinensis and A. chinensis var. deliciosa commercial cultivars. Bounty 71 was chosen, as it is commercialised as a tolerant genotype to root asphyxia. Vines were grown in standard greenhouse conditions under natural light (relative humidity: 60%, temperature: 20 to 24 °C) and maintained with standard NPK fertilisation supplied as in fertigation.
Since waterlogging seems to be one of the drivers of KVDS onset, half of the plants were subjected to irrigation schedule exceeding the soil field capacity (FC), whereas the remaining plants received irrigation volumes accounting to approximately 70% of the estimated FC. The higher water irrigation volumes were supplied to mimic the flooding system adopted in most affected orchards. In a preliminary test, soil FC was determined for each pot by drying the soil at 110 °C until a constant weight was obtained. Subsequently, water was applied to the dried-up soil, until percolation occurred. FC was calculated as the volume of water at which percolation started and expressed as a percentage relative to the weight of each pot. During the experimental period, pots were periodically weighed and the difference in weight on subsequent days was corrected by adding water to maintain the required FC. The two irrigation strategies were maintained from March to the first week of September, when inoculation was performed. During the experiment, to avoid the increase in weight due to the growth of the foliage, the plants were periodically pruned and the material weighed.
Four vines in both irrigation conditions were inoculated with one of the fungal pathogens isolated from affected vines (
Phytophthora megasperma,
Phytophthora infestans,
Phytopythium spp. or
Desarmillaria tabescens).
Phytopythium helicoides CBS 286.31 and
P. fagopyri CBS 293 were included as reference strains. The remaining 8 plants were used as uninoculated controls.
D. tabescens isolates were routinely cultured on potato dextrose agar (PDA) at 25 °C in the dark. Woody inoculum was prepared according to Mansilla et al. [
26], with slight modifications. To this end, fresh twig fragments (5–6 cm × 1.5–2 cm diameter, bark included) of
Actinidia chinensis var.
deliciosa were placed overnight in water. Thereafter, water was removed and boxes containing woody materials were autoclaved. After cooling, 250 mL of sterile PDA medium were added, and each box was inoculated by placing active mycelial culture fragments onto the stem segments. Boxes were incubated in the dark at 25 °C for 2 weeks. For inoculation in each pot, two inoculated shoots were buried in the soil adjacent to the main root. Control plants were mock inoculated with sterile twig segments.
Pure cultures of Phytophthora, Phytopythium and Pythium isolates were grown on potato dextrose agar medium (PDA) for 1 week at 25 °C in the dark. To induce sporangium production, active mycelial culture fragments were transferred to soil extract medium. Air-dried garden soil (with high content of organic matter) was diluted (1%, w/v) in distilled water and sterilised for 1 h at 121 °C. After a preliminary sedimentation at room temperature, the supernatant was centrifuged, the pH of the clear solution was adjusted to 6.8–7.0 and sterilised. The fungal-soil suspension was then placed in the dark at 15–18 °C for 48 h. Resulting sporangia were counted using a hemocytometer, and 50 mL of a suspension of sporangia (105 CFU mL−1) were used to inoculate each plant. Plants inoculated with sterile medium served as a control. Zoospores were released by placing the suspension at 4 °C for 1 h followed by another hour at 25 °C.
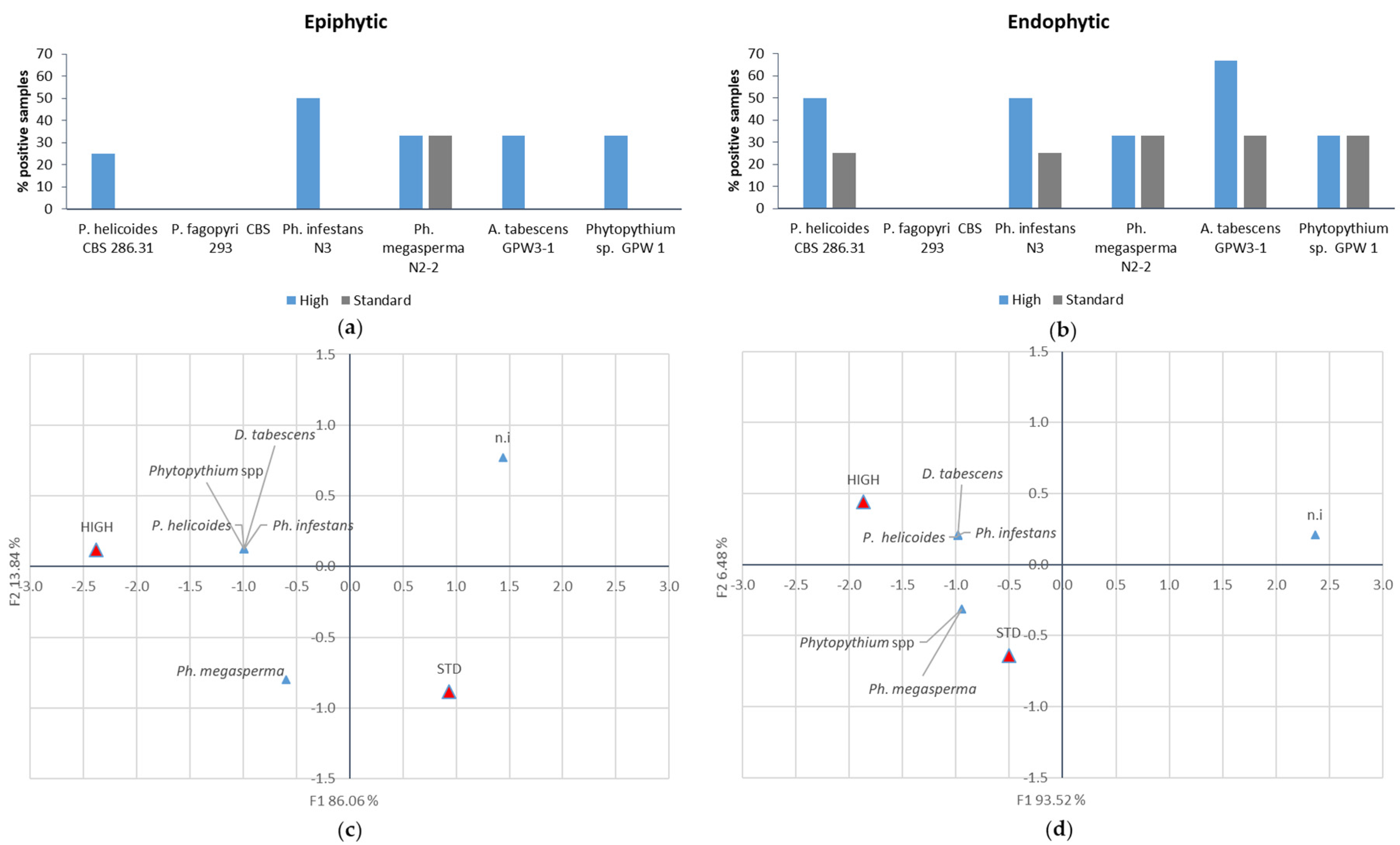
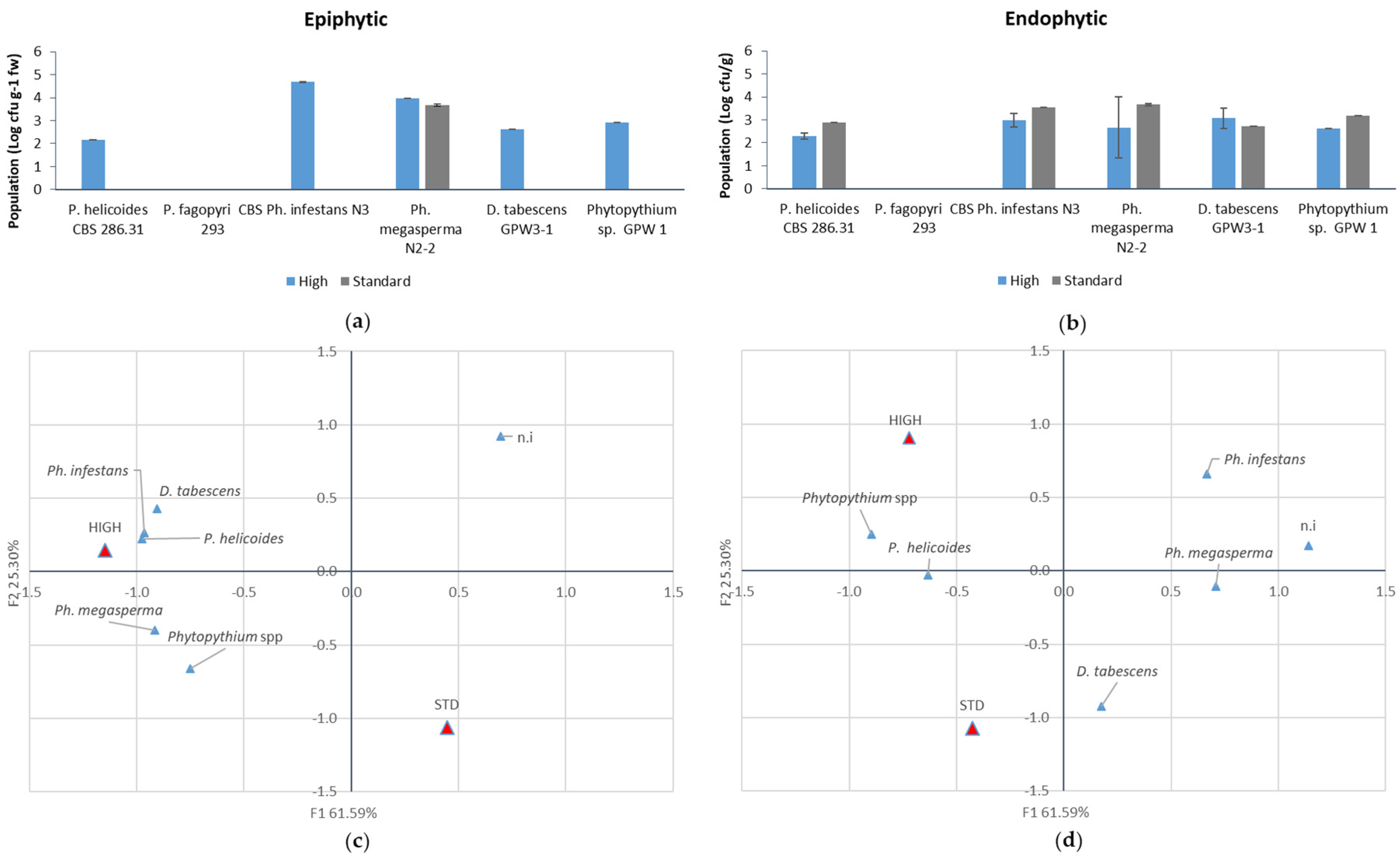
Plants were maintained at 22 ± 3 °C and the symptom evolution (chlorotic leaves and mortality incidence) was monitored weekly. Fungal populations were determined six weeks after inoculation. Firstly, to assess epiphytic populations, root samples were vigorously washed for 5 min in 15 mL sterile distilled water containing 0.05% (v/v) Tween 80. The concentration and identity of fungal suspensions were evaluated by plating 100-µL aliquots of serial 1:10 dilutions on PDA amended with rifampicin (20 mg L−1), streptomycin (100 mg L−1) and tetracycline (50 mg L−1). The growth pattern, mycelium structure and sporangial morphology of the individual isolates were examined under microscope for identification.
To assess the endophytic populations, the same tissue samples were surface sterilised by washing in 70% ethanol (2 min) and in 0.1 % NaOCl (2 min) solution, followed by rinses with sterile water. Successively, they were homogenised in 15 mM sterile distilled water containing 0.05% (v/v) Tween 80 and population was assessed as previously described.
2.4. Statistical Analysis
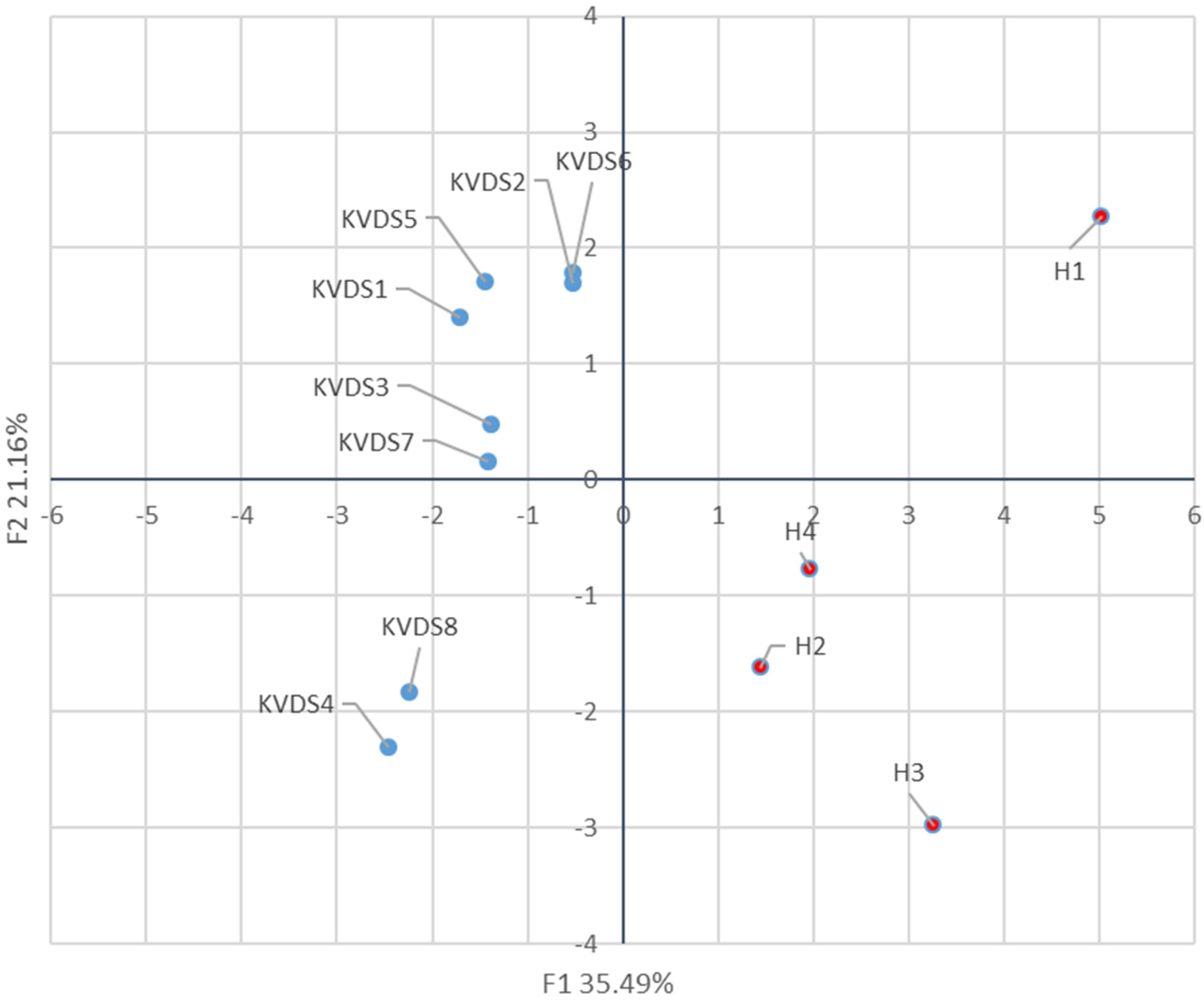
Principal component analysis (PCA) was performed on bacterial relative abundance, or on the fungal populations and percentage of isolation in test plants with Statistica 7.0 (StatSoft Inc., Tulsa, OK, USA). Incidence in greenhouse inoculation trials was analysed by the Marascuilo procedure (χ2 = 0.05). The effect of the irrigation strategy on the percentage of epiphytically and endophytically colonised roots was analysed by Z-test (p = 0.05). Population data were expressed as mean ± standard error (S.E.).
4. Discussion
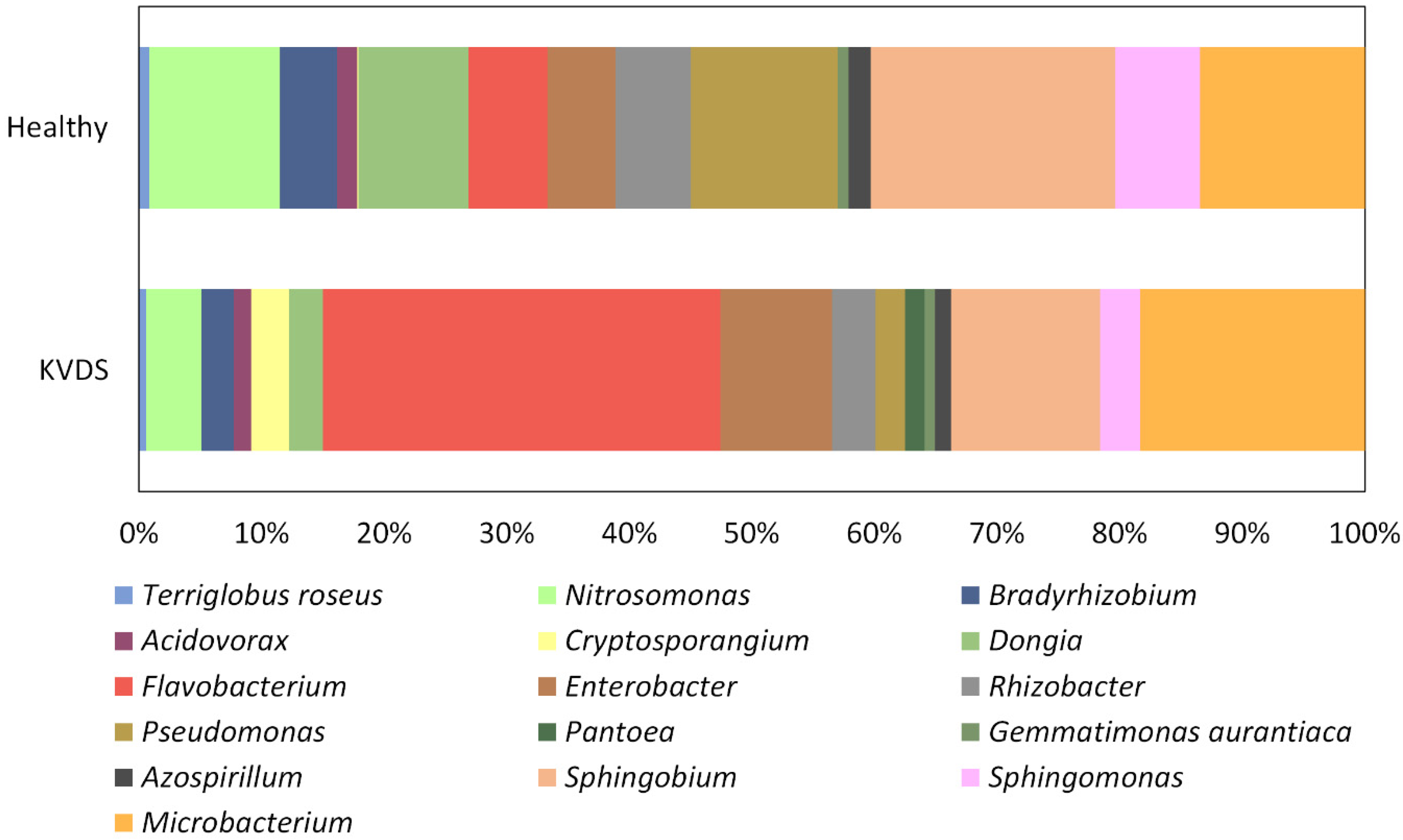
The extent of fruit and forest tree decline has constantly increased in recent years, urging the identification of the underlying mechanisms. Since different biotic and abiotic factors contribute to woody plant declines and symptoms are often nonspecific and chronical, rather than acute, these syndromes could be considered multifactorial diseases [
28,
29]. In this view, pathogenic agents may act either as primary or secondary factors when plants have already been weakened by climatic, environmental or agricultural stresses [
29]. This research aimed at verifying potential implications of the root microbial community on the onset and development of the KVDS epidemic outbreak in Italy. Asymptomatic and symptomatic roots could be discriminated based on their bacterial community. Nonetheless, no pathogenic bacteria were isolated from kiwifruit root system, suggesting that the bacterial community does not play a significant role in KVDS onset. The differences between the bacterial community of healthy and KVDS roots could relate to different biotic and abiotic conditions the roots are exposed to, or alternatively, to differences in the physiological status of the roots, which may in turn influence the resident microbial communities [
30].
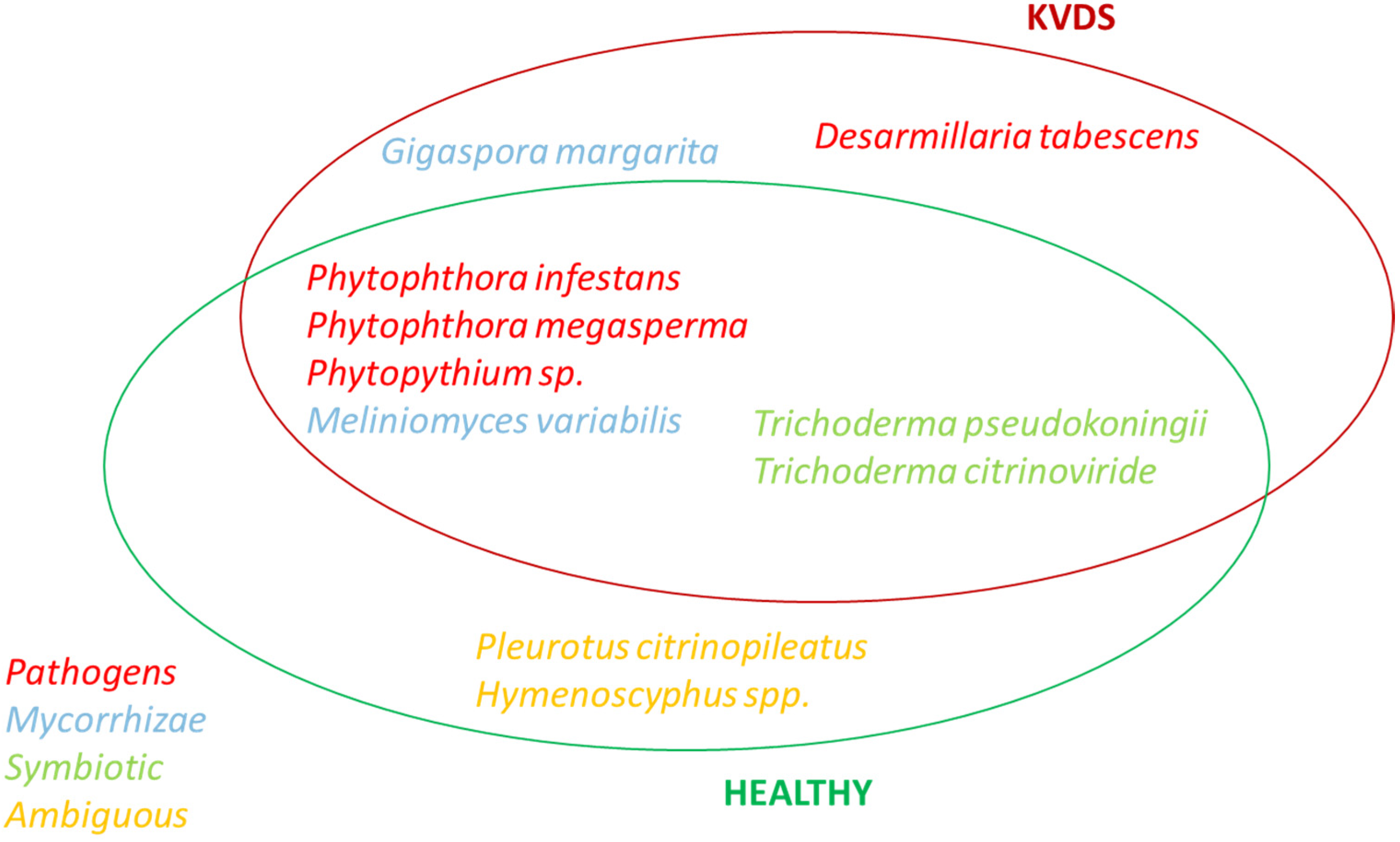
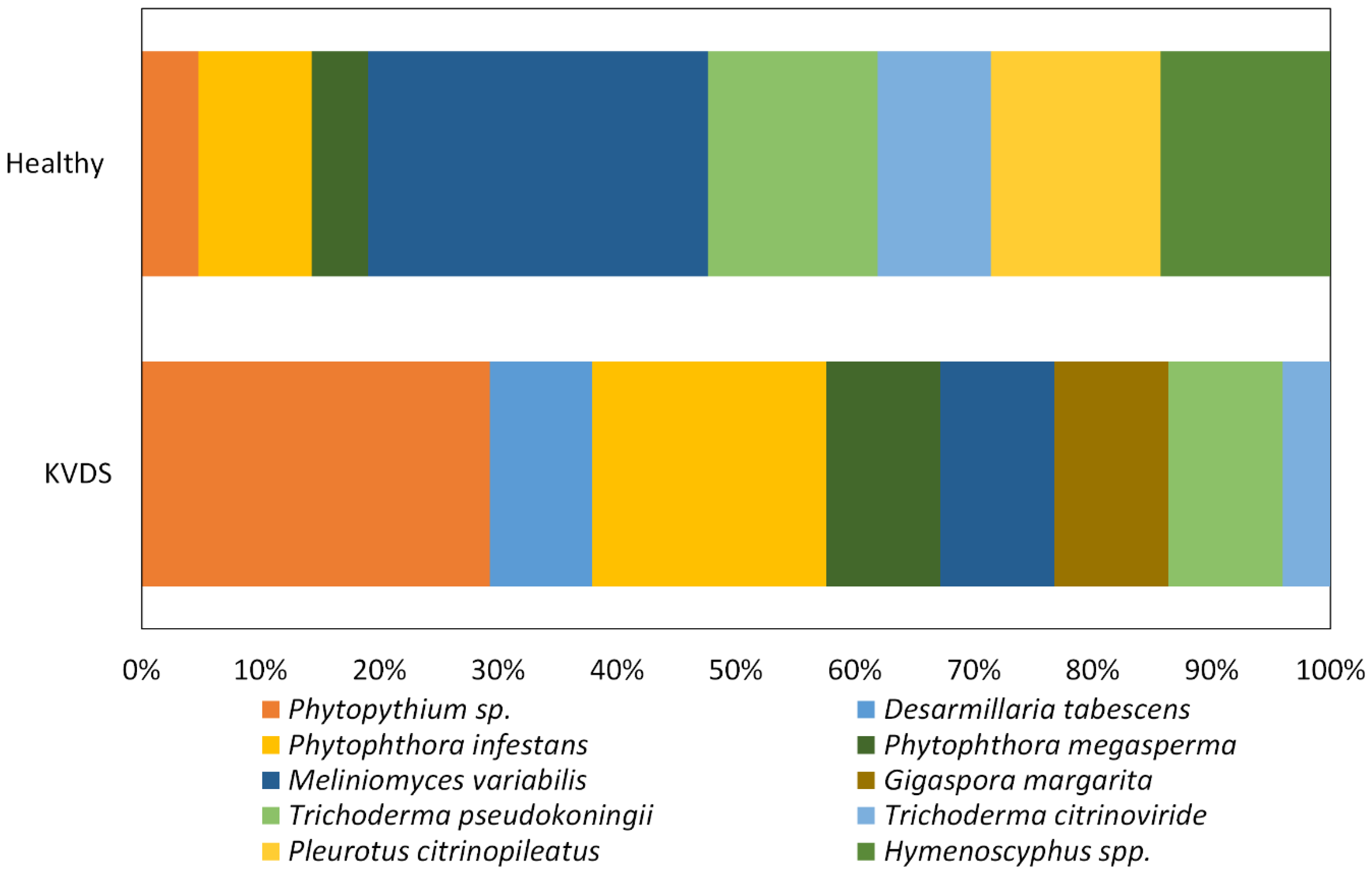
Three main pathogenic fungal genera were isolated from roots of kiwifruit tissues, namely
Phytophthora,
Phytopythium and
Desarmillaria. Therefore, similarly to other woody plant declines, different fungal pathogens may contribute to the development of KVDS [
31]. These fungi were present both in healthy and in symptomatic kiwifruit vines. Nonetheless, healthy and symptomatic vines were discriminated by the frequency of isolation of these pathogens. Moreover, KVDS plants showed a lower frequency of symbiotic fungal species. Indeed, woody plant decline is often not directly associated with single specific pathogens, but it is related to the dysbiosis of the plant-associated microbiome, resulting in an imbalance in the functions exerted by the microbial community [
29,
32].
Finally, D. tabescens was exclusively isolated from the roots of symptomatic plants. However, the fact that these fungi colonised the roots once vines were already suffering from KVDS cannot be excluded.
Several
Phytophthora species are responsible for important losses both in agricultural and natural ecosystems.
Phytophthora root rot diseases are common in heavy and wet soils as well as in excessively watered orchards [
17]. Previous studies have indicated that most of the pathogenic fungi associated to kiwifruit root rot belong to the genus
Phytophthora, and different species or varieties occur in different countries. For example,
Ph. cryptogea and
Ph. citrophthora were reported in Chile [
33];
Ph. nicotianae in Italy [
34];
Ph. megasperma in France [
17];
Ph. cactorum,
Ph. citrophthora,
Ph. cryptogea,
Ph. drechsleri, and
Ph. megasperma in USA [
35];
Ph. cryptogea,
Ph. cinnamoni,
Ph. citricola,
Ph. gonapodyides and
Ph. stellata in New Zealand. Among these species,
Ph. cryptogea and
Ph. cinnamoni are the most pathogenic [
36].
Phytopythium is a newly established genus including fungi formerly classified as
Pythium species [
33]; to date, sixteen species belonging to this genus have been described.
Phytopythium is characterised by morphological features common to both
Pythium and
Phytophthora leading to the reclassification of these species out of the genus
Pythium in 2010, after genomic studies [
37]. These fungi inhabit both water and soil, and several species, such as
P. helicoides, are soil and water-borne plant pathogens causing root rot diseases.
P. helicoides has already been identified in kiwifruit as the causal agent of root and collar rot [
20] that produces symptoms at root level partially overlapping with the one observed in KVDS-affected vines. The molecular identification could not discriminate between
Phytopythium helicoides and
Phytopythium fagopyri. These two species are extremely close to each other, sharing wide portions of their genome, thus hindering a univocal identification by DNA sequencing based only on ITS region (
Table 1). The strain of
Phytopythium (GPW-1) isolated in this work had a behaviour similar to
P. helicoides (CBS286.31) used as a reference, independently of the irrigation strategy. This result suggests that the strain GPW1 has more ecological and pathogenic similarity with
P. helicoides than
P. fagopyri. DNA barcoding based on different regions of the ribosomal operon may allow us to discriminate between these two species [
38]. If confirmed, this finding would be the first report of
Phytopythium helicoides affecting kiwifruit vines in Italy.
The species belonging to
Armillaria and closely related genera are characterised by different lifestyles, as saprophytic, pathogenic and symbiotic species belong to the genus. Pathogenic species such as
Desarmillaria tabescens and
Armillaria mellea are responsible for severe root rot on many horticultural crop plants [
39,
40]. The natural habitat of these fungi is primarily represented by forests of hardwoods or mixed conifers, but very often populations of these fungi parasitise woody crops. Indeed,
Armillaria also causes root rot and death in kiwifruit vines [
22]. Orchard epidemics may start from infected wood remaining in the soil after land was cleared of forest trees to establish the plantations, or from roots of shelterbelt trees that had been felled, and whose stumps had been colonised by basidiospores [
22]. Despite the wide geographical distribution of
D. tabescens and its occurrence in southern Europe [
41,
42,
43], the current work is the first report of kiwifruit infection in Italy. Moreover, the isolation of
D. tabescens from kiwifruit vines has not been confirmed in previous studies.
Woody plant infection caused by
D. tabescens is exacerbated by climate change [
44,
45]. Indeed, waterlogging could facilitate infection, since
D. tabescens produces melanised rhizomorphs in water-saturated environments [
39]. Furthermore, conditions of periodic water saturation can facilitate
D. tabescens passive spread to roots in a single root system, among adjacent root systems, and even among root systems of sequential generations of trees [
46]. Finally, stressing conditions may hinder root defences against pathogens, for instance, as a result of depletion of energy reserves or metabolic imbalance.
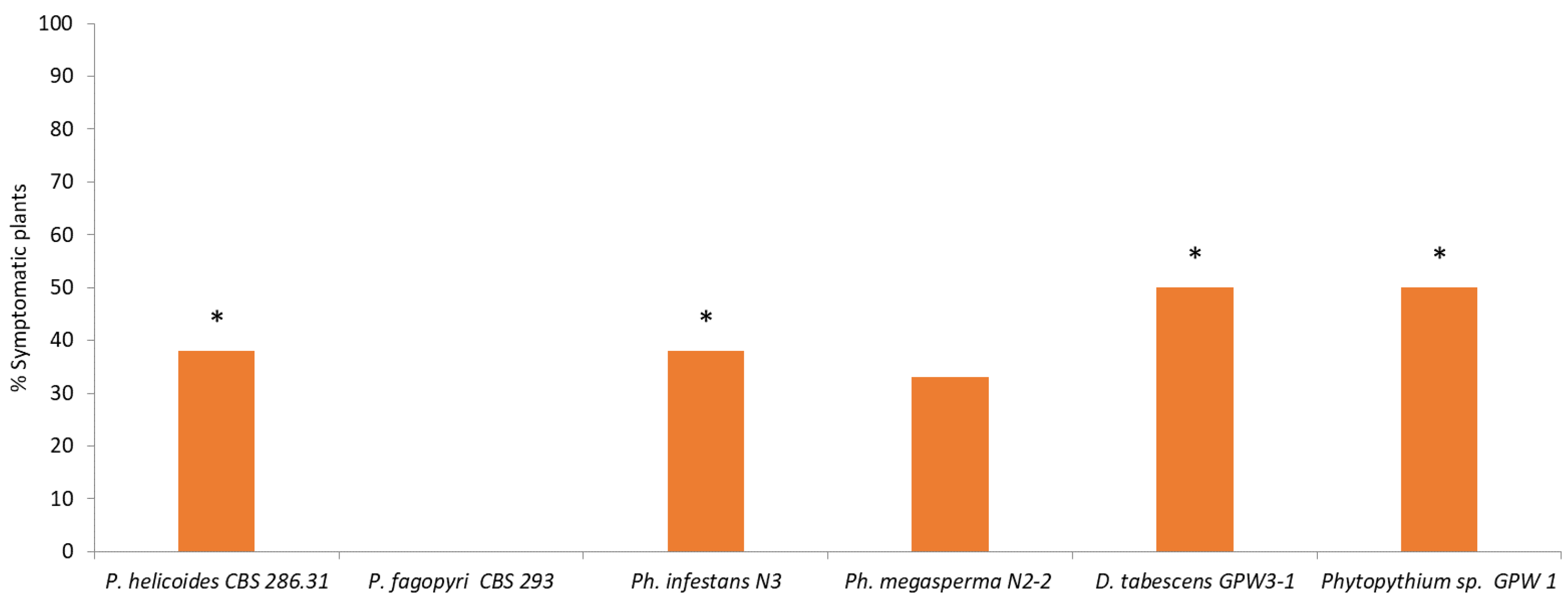
The greenhouse experiment confirmed that the four pathogens identified as Ph. infestans, Ph. megasperma, P. helicoides and D. tabescens are able to induce macroscopic KVDS symptoms both at root and canopy level. An excessive soil water content induces the growth of larger populations of pathogenic fungi on the root surface and inner tissues. The presence of high pathogenic populations (105 CFU g−1 FW) on the root surface of the samples exposed to higher water supply may account for the rapid spread of the decline from an infected vine to the neighbouring ones. In fact, when high pathogen populations are present on the outer layer of the roots, rather than in the inner tissues, they may act as an inoculation source for the surrounding vines. Moreover, the higher incidence and size of the epiphytic population may indicate that, in waterlogging conditions, pathogens may have a faster growth and a possible leak from internal root tissues. In this view, the high water content in the soil can further facilitate the diffusion of the pathogens, especially for those, such as Phytophthora and Phytopythium, which produce motile zoospores.
5. Conclusions
Our research suggests that fungal pathogens may play a role in KVDS epidemiology, but other drivers, such as waterlogging conditions and asphyxia, are triggering factors for the expression of KVDS symptomatology. Among the pathogenic fungi, Phytopythium was the most frequently isolated pathogen from infected roots and D. tabescens was isolated only in highly symptomatic vines, suggesting that these pathogens could have a crucial role in KVDS development. However, this study does not identify a univocal pathogen as the causal agent of KVDS symptoms. In fact, different fungal isolates may act as a pathogenic consortium, able to critically damage kiwifruit root system and causing KVDS on the epigeal part of the vine.
This study shed light on the possible correlation among the population of different pathogens contributing to KVDS. However, to corroborate the obtained results, the study should be expanded to a higher number of orchards and to different geographic areas, where KVDS has been observed. Furthermore, coupling environmental monitoring of soil conditions with the temporal dynamic studies of the different fungal populations associated with kiwifruit roots may distinguish among the fungal populations that act as drivers of KVDS and the ones that are modified as a consequence of KVDS. For this purpose, next generation sequencing (NGS) technologies and high-throughput screening (HTS) for metagenomic libraries would help in better understanding the role of microbial communities in KVDS onset and development. Finally, our study cannot exclude that other pathogenic organisms, such as phytoplasmas or viruses, may be involved in KVDS onset.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}