Assessing Salicornia europaea Tolerance to Salinity at Seed Germination Stage




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Germination Conditions
2.2. Germination Indices
2.3. Statistical Analysis
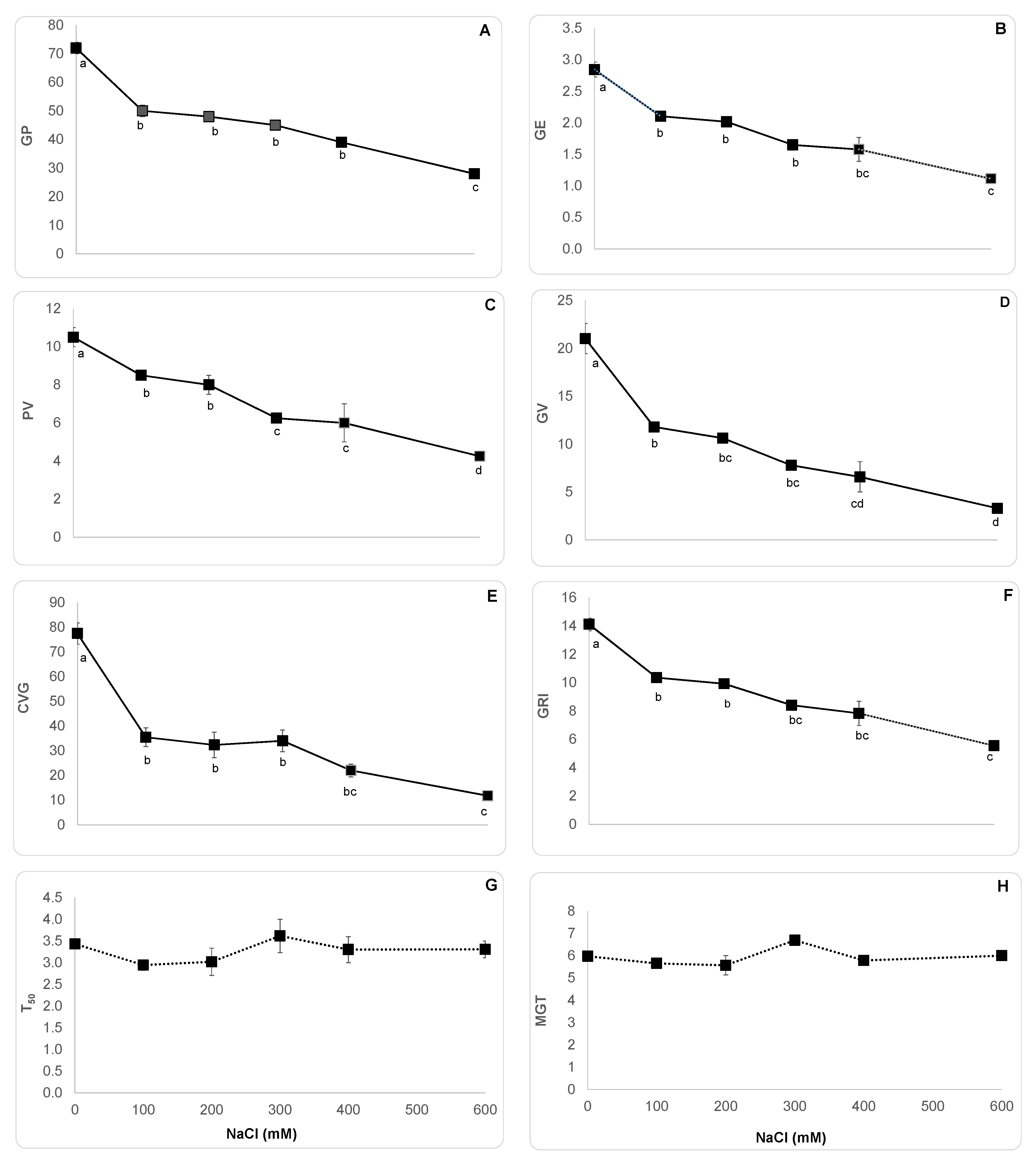
3. Results
3.1. Salt Effects on Seed Germination Indices
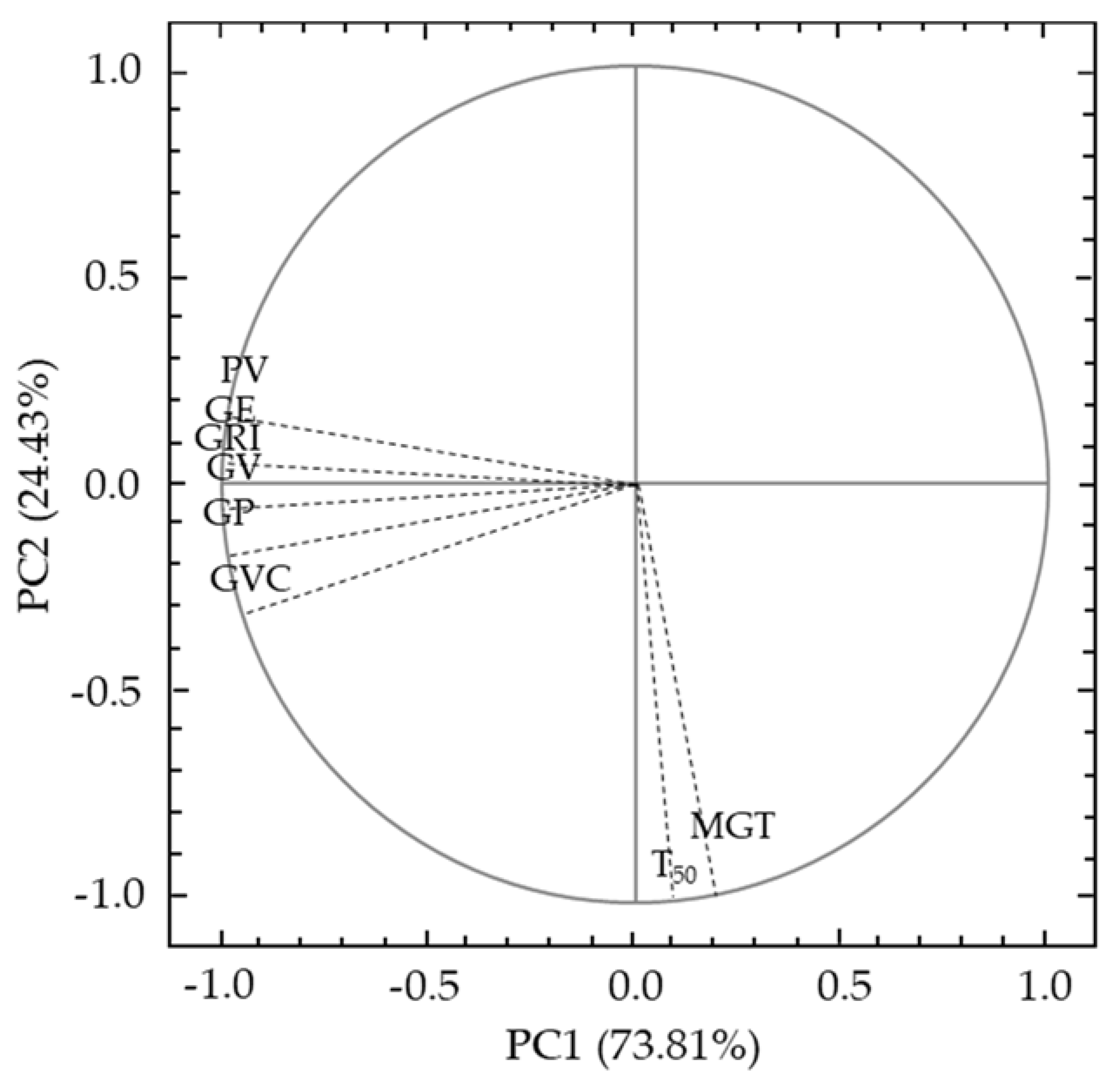
3.2. Principal Component Analysis of Germination Indices
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shannon, M.C. Adaptation of Plants to Salinity. Adv. Agron. 1997, 60, 75–120. [Google Scholar]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. CRC. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of Salt Tolerance in Nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Patel, S. Salicornia: Evaluating the halophytic extremophile as a food and a pharmaceutical candidate. 3 Biotech 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Buhmann, A.K.; Flowers, T.J.; Seal, C.E.; Papenbrock, J. Salicornia as a crop plant in temperate regions: Selection of genetically characterized ecotypes and optimization of their cultivation conditions. AoB Plants 2014, 6, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Oguri, S.; Chiba, S.; Momonoki, Y.S. Molecular cloning of acetylcholinesterase gene from Salicornia europaea L. Plant Signal. Behav. 2009, 4, 361–366. [Google Scholar] [CrossRef]
- Kim, S.; Kim, E.; Yoon, S.; Jo, N.; Jung, S.K.; Kwon, S.; Chang, Y.H.; Jeong, Y. Physicochemical and Microbial Properties of the Korean Traditional Rice Wine, Makgeolli, Supplemented with Banana during Fermentation. Prev. Nutr. Food Sci. 2013, 18, 203–209. [Google Scholar] [CrossRef]
- Song, S.H.; Lee, C.; Lee, S.; Park, J.M.; Lee, H.J.; Bai, D.H.; Yoon, S.S.; Choi, J.B.; Park, Y.S. Analysis of microflora profile in Korean traditional Nuruk. J. Microbiol. Biotechnol. 2013, 23, 40–46. [Google Scholar] [CrossRef]
- Gunning, D. Cultivating Salicornia europaea (Marsh Samphire); Daithi O’Murchu Marine Research Station & University College Cork: Dublin, Ireland, 2016; p. 92. [Google Scholar]
- Cybulska, I.; Chaturvedi, T.; Brudecki, G.P.; Kádár, Z.; Meyer, A.S.; Baldwin, R.M.; Thomsen, M.H. Chemical characterization and hydrothermal pretreatment of Salicornia bigelovii straw for enhanced enzymatic hydrolysis and bioethanol potential. Bioresour. Technol. 2014, 153, 165–172. [Google Scholar] [CrossRef]
- Smillie, C. Salicornia spp. as a biomonitor of Cu and Zn in salt marsh sediments. Ecol. Indic. 2015, 56, 70–78. [Google Scholar] [CrossRef]
- Shpigel, M.; Ben-Ezra, D.; Shauli, L.; Sagi, M.; Ventura, Y.; Samocha, T.; Lee, J.J. Constructed wetland with Salicornia as a biofilter for mariculture effluents. Aquaculture 2013, 412–413, 52–63. [Google Scholar] [CrossRef]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically Rethinking Agriculture for the 21st Century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed]
- Essaidi, I.; Brahmi, Z.; Snoussi, A.; Ben Haj Koubaier, H.; Casabianca, H.; Abe, N.; El Omri, A.; Chaabouni, M.M.; Bouzouita, N. Phytochemical investigation of Tunisian Salicornia herbacea L., antioxidant, antimicrobial and cytochrome P450 (CYPs) inhibitory activities of its methanol extract. Food Control 2013, 32, 125–133. [Google Scholar] [CrossRef]
- Shin, M.G.; Lee, G.H. Spherical granule production from micronized saltwort (Salicornia herbacea) powder as salt substitute. Prev. Nutr. Food Sci. 2013, 18, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Lim, G.S.; Piao, Y.L.; Choi, O.Y.; Cho, K.A.; Park, C.B.; Chang, Y.C.; Song, Y.I.; Lee, M.K.; Cho, H. Characterization, stability, and antioxidant activity of Salicornia herbacea seed oil. Korean J. Chem. Eng. 2014, 31, 2221–2228. [Google Scholar] [CrossRef]
- Ryu, D.-S.; Kim, S.-H.; Lee, D.-S. Anti-Proliferative Effect of Polysaccharides from Salicornia herbacea on Induction of G2/M Arresta nd Apoptosis in Human Colon Cancer Cells. J. Microbiol. Biotechnol. 2009, 19, 1482–1489. [Google Scholar] [CrossRef]
- Lee, S.S.; Seo, H.; Ryu, S.; Kwon, T.-D. The effect of swimming exercise and powdered-Salicornia herbacea L. ingestion on glucose metabolism in STZ-induced diabetic rats. J. Exerc. Nutr. Biochem. 2015, 19, 235–245. [Google Scholar] [CrossRef][Green Version]
- Norman, H.C.; Masters, D.G.; Barrett-Lennard, E.G. Halophytes as forages in saline landscapes: Interactions between plant genotype and environment change their feeding value to ruminants. Environ. Exp. Bot. 2013, 92, 96–109. [Google Scholar] [CrossRef]
- Sharma, A.; Gontia, I.; Agarwal, P.K.; Jha, B. Accumulation of heavy metals and its biochemical responses in Salicornia brachiata, an extreme halophyte. Mar. Biol. Res. 2010, 6, 511–518. [Google Scholar] [CrossRef]
- Lei, L.; Liang, D.; Yu, D.; Chen, Y.; Song, W.; Li, J. Human health risk assessment of heavy metals in the irrigated area of Jinghui, Shaanxi, China, in terms of wheat flour consumption. Environ. Monit. Assess. 2015, 187, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.X.; Huang, Z.Y.; Baskin, J.M.; Baskin, C.C. Effect of temperature, light and salinity on seed germination and radicle growth of the geographically widespread halophyte shrub Halocnemum strobilaceum. Ann. Bot. 2008, 101, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Atia, A.; Debez, A.; Barhoumi, Z.; Smaoui, A.; Abdelly, C. ABA, GA3, and nitrate may control seed germination of Crithmum maritimum (Apiaceae) under saline conditions. Comptes Rendus Biol. 2009, 332, 704–710. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, X.; Zheng, G.; Gutterman, Y. Influence of light, temperature, salinity and storage on seed germination of Haloxylon ammodendron. J. Arid Environ. 2003, 55, 453–464. [Google Scholar] [CrossRef]
- Muhammad, Z.; Hussain, F. Effect of NaCl salinity on the germination and seedling growth of seven wheat genotypes. Pakistan J. Bot. 2012, 44, 1845–1850. [Google Scholar]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Williams, M.; Ungar, I. The Effect of Environmental Parameters on the Germination, Growth, and Development of Suaeda depressa (Pursh) Wats. Am. J. Bot. 1972, 59, 912–918. [Google Scholar] [CrossRef]
- Orlovsky, N.; Japakova, U.; Zhang, H.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers. 2016, 38, 183–189. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Al-Mudaris, M.A. Notes on Various Parameters Recording the Speed of Seed Germination. J. Agric. Trop. Subtrop. 1998, 99, 147–154. [Google Scholar]
- Mangure, J.D. Speed of germination: Aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar]
- Czabator, F.J. Germination Value: An Index Combining Speed and Completeness of Pine Seed Germination. For. Sci. 1962, 8, 386–396. [Google Scholar]
- Jones, K.W.; Sanders, D.C. The Influence of Soaking Pepper Seed in Water or Potassium Salt Solutions on Germination at Three Temperatures. J. Seed Technol. 1987, 11, 97–102. [Google Scholar]
- Farooq, M.; Basra, S.M.A.; Ahmad, N.; Hafeez, K. Thermal hardening: A new seed vigor enhancement tool in rice. J. Integr. Plant Biol. 2005, 47, 187–193. [Google Scholar] [CrossRef]
- Orchard, T.J. Estimating the parameters of plant seedling emergence. Seed Sci. Technol. 1977, 5, 61–69. [Google Scholar]
- Boubaker, M. Salt tolerance of durum wheat cultivars during germination and early seedling growth. Agric. Medit. 1996, 126, 32–39. [Google Scholar]
- Rahman, M.S.; Miyake, H.; Takeoka, Y. Effect of Sodium Chloride Salinity on Seed Germination and Early Seedling Growth of Rice (Oryza sativa L.). Pakistan J. Biol. Sci. 2001, 4, 351–355. [Google Scholar]
- Hakim, M.A.; Juraimi, A.S.; Begum, M.; Hanafi, M.M.; Ismail, M.R.; Selamat, A. Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). African J. Biotechnol. 2010, 9, 1911–1918. [Google Scholar]
- Sanoubar, R.; Cellini, A.; Veroni, A.M.; Spinelli, F.; Masia, A.; Vittori Antisari, L.; Orsini, F.; Gianquinto, G. Salinity thresholds and genotypic variability of cabbage (Brassica oleracea L.) grown under saline stress. J. Sci. Food Agric. 2016, 96, 319–330. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; De Pascale, S. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Wei, Y.; Dong, M.; Huang, Z.Y.; Tan, D.Y. Factors influencing seed germination of Salsola affinis (Chenopodiaceae), a dominant annual halophyte inhabiting the deserts of Xinjiang, China. Flora Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 134–140. [Google Scholar] [CrossRef]
- Yang, H.; Huang, Z.; Baskin, C.C.; Baskin, J.M.; Cao, Z.; Zhu, X.; Dong, M. Responses of caryopsis germination, early seedling growth and ramet clonal growth of Bromus inermis to soil salinity. Plant Soil 2009, 316, 265–275. [Google Scholar] [CrossRef]
- Keshavarzi, M.H.B. The effect of different NaCl concentration on germination and early seedling growth of Artemisia annua L. Int. J. Agric. 2012, 2, 135–140. [Google Scholar]
- Li, Y. Effect of salt stress on seed germination and seedling growth of three salinity plants. Pakistan J. Biol. Sci. PJBS 2008, 11, 1268–1272. [Google Scholar] [CrossRef] [PubMed]
- Xianzhao, L.; Chunzhi, W.; Qing, S. Screening for Salt Tolerance in Eight Halophyte Species from Yellow River Delta at the Two Initial Growth Stages. ISRN Agron. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, Y.-C. Influence of light, temperature and salinity on seed germination of Reaumuria trigyna Maxim. Plant Physiol. Commun. 2007, 43, 708–710. [Google Scholar]
- Guma, I.R.; Padrón-Mederos, M.A.; Santos-Guerra, A.; Reyes-Betancort, J.A. Effect of temperature and salinity on germination of Salsola vermiculata L. (Chenopodiaceae) from Canary Islands. J. Arid Environ. 2010, 74, 708–711. [Google Scholar] [CrossRef]
- Chookhampaeng, S. The Effect of Salt Stress on Growth, Chlorophyll Content Proline content and antioxidative enzymes of Pepper (Capsicum annuum L.) seedling. Eur. J. Sci. Res. 2011, 49, 103–109. [Google Scholar]
- Kriegel, H.P.; Kröger, P.; Schubert, E.; Zimek, A. A General Framework for Increasing the Robustness of PCA-Based Correlation Clustering Algorithms; Springer: Berlin/Heidelberg, Germany, 2005. [Google Scholar]
- Jolliffe, I.T. Principal components analysis. In International Encyclopedia of Education; Springer: Berlin, Germany, 2010; pp. 374–377. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Index | Unit | Formula | Ref. |
|---|---|---|---|
| Germination Percentage (GP) | % | [31] | |
| Germination Energy (GE) | ------ | [32] | |
| where N is the number of germinated seeds on the counting date and D the number of days | |||
| Peak Value (PV) | ------ | [33] | |
| where Mag is the maximum of seeds accumulative germination | |||
| Germination Value (GV) | ------ | 1 | [33] |
| Coefficient of Germination Velocity (CVG) | ------ | [34] | |
| where N is the number of germinated seeds every day and D is the number of days from seeding corresponding to N. | |||
| Germination Rate Index (GRI) | % day−1 | [31] | |
| where G1 is the germination percentage on day 1 (D1), and so on | |||
| Time to 50% germination (T50) | days | [35] | |
| where N is the final number of germinated seeds, and ni and nj are the total number of seeds that had germinated (by adjacent counts) at times ti and tj, when ni < N/2 < nj. | |||
| Mean Germination Time (MGT) | days | [36] | |
| where N is the number of seeds germinated on day D |
| Principal Component Analysis | PC1 | PC2 |
|---|---|---|
| Eigenvalue | 5.905139 | 1.954265 |
| Total variance (%) | 73.81423 | 24.42831 |
| Cumulative Eigenvalue | 5.905139 | 7.859403 |
| Cumulative variance (%) | 73.8142 | 98.2425 |
| Variable Squared Cosines | ||
| Variable | PC1 | PC2 |
| GP | 0.98 | 1.00 |
| GE | 1.00 | 1.00 |
| PV | 0.97 | 0.99 |
| GV | 0.99 | 0.99 |
| CVG | 0.93 | 0.99 |
| GRI | 1.00 | 1.00 |
| T50 | 0.00 | 0.95 |
| MGT | 0.03 | 0.94 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calone, R.; Sanoubar, R.; Noli, E.; Barbanti, L. Assessing Salicornia europaea Tolerance to Salinity at Seed Germination Stage. Agriculture 2020, 10, 29. https://doi.org/10.3390/agriculture10020029
Calone R, Sanoubar R, Noli E, Barbanti L. Assessing Salicornia europaea Tolerance to Salinity at Seed Germination Stage. Agriculture. 2020; 10(2):29. https://doi.org/10.3390/agriculture10020029
Chicago/Turabian StyleCalone, Roberta, Rabab Sanoubar, Enrico Noli, and Lorenzo Barbanti. 2020. "Assessing Salicornia europaea Tolerance to Salinity at Seed Germination Stage" Agriculture 10, no. 2: 29. https://doi.org/10.3390/agriculture10020029
APA StyleCalone, R., Sanoubar, R., Noli, E., & Barbanti, L. (2020). Assessing Salicornia europaea Tolerance to Salinity at Seed Germination Stage. Agriculture, 10(2), 29. https://doi.org/10.3390/agriculture10020029