Contrasting Response Mechanisms of Maize Lines to Striga hermonthica

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
2.1. Plant Materials
2.2. Preparation of Long-Ashton Solution
2.3. Germination-Inducing Activity of Root Exudates
2.3.1. Test Tube Experiment
2.3.2. Pot Experiment
2.4. Root Architecture Evaluation
2.5. Rhizotron Studies
2.5.1. Determination of the Number of Attached S. hermonthica Plants and Maize Plant Responses to Infestation
2.5.2. Evaluation of Physiological Changes Due to S. hermonthica Infestation
2.5.3. Determination of the Growth Rate of Attached S. hermonthica Plants
2.6. Field Evaluation of the Two Maize Lines (5057 and TZISTR1108)
2.7. Statistical Analysis
3. Results
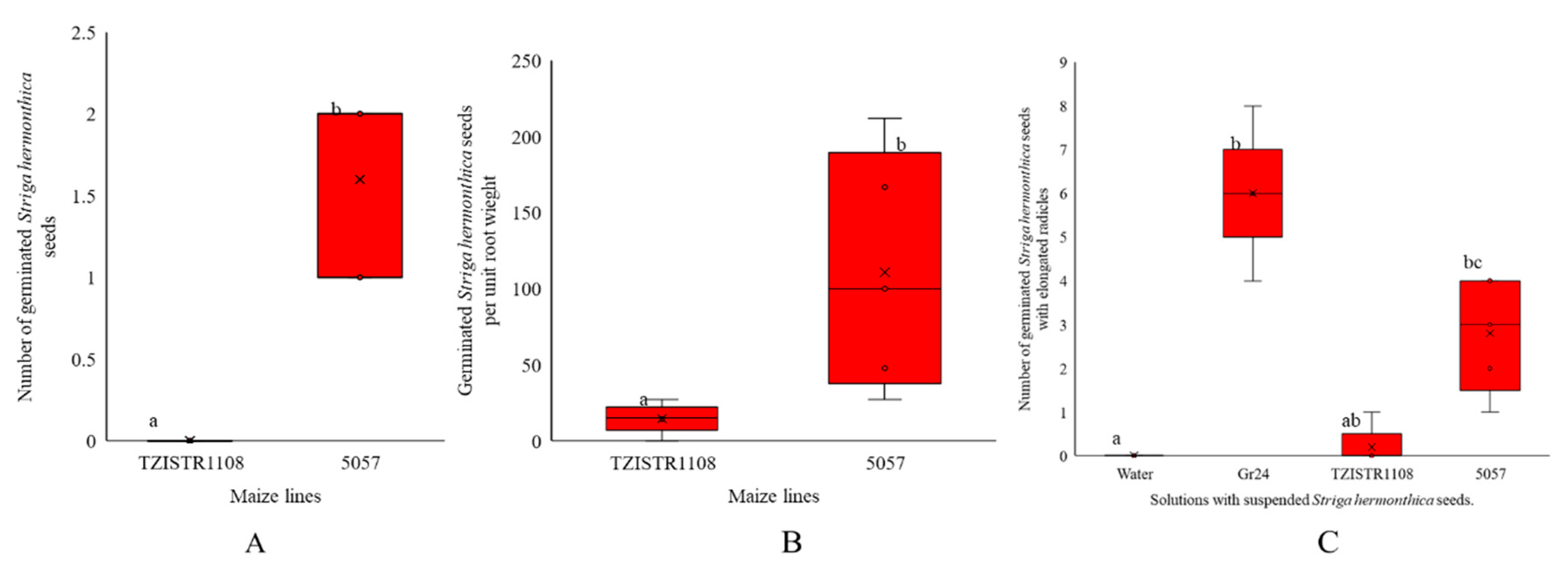
3.1. Evaluation of the Germination-Inducing Activity of Root Exudates
3.1.1. Test Tube Experiments
3.1.2. Pot Experiments
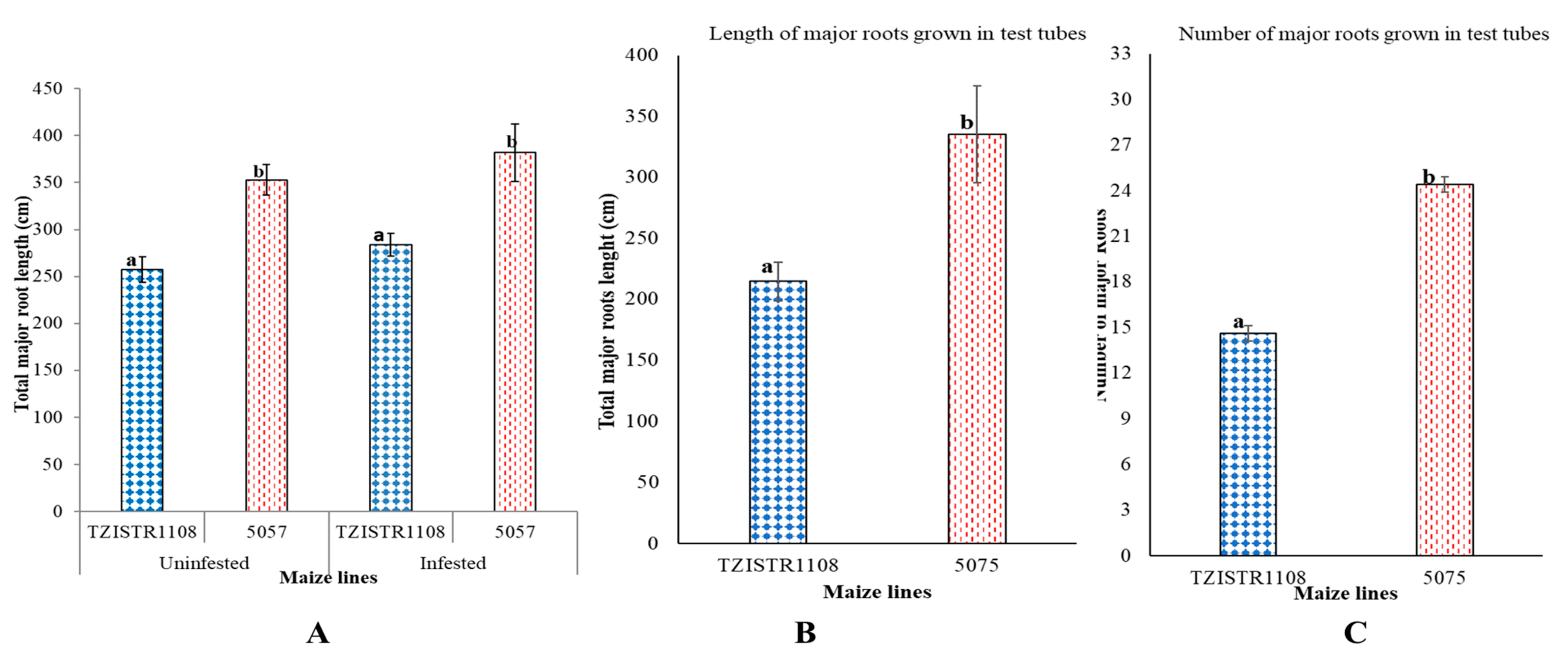
3.2. Root Architecture Evaluation
3.3. Rhizotron Studies (Post-Attachment Resistance Evaluation)
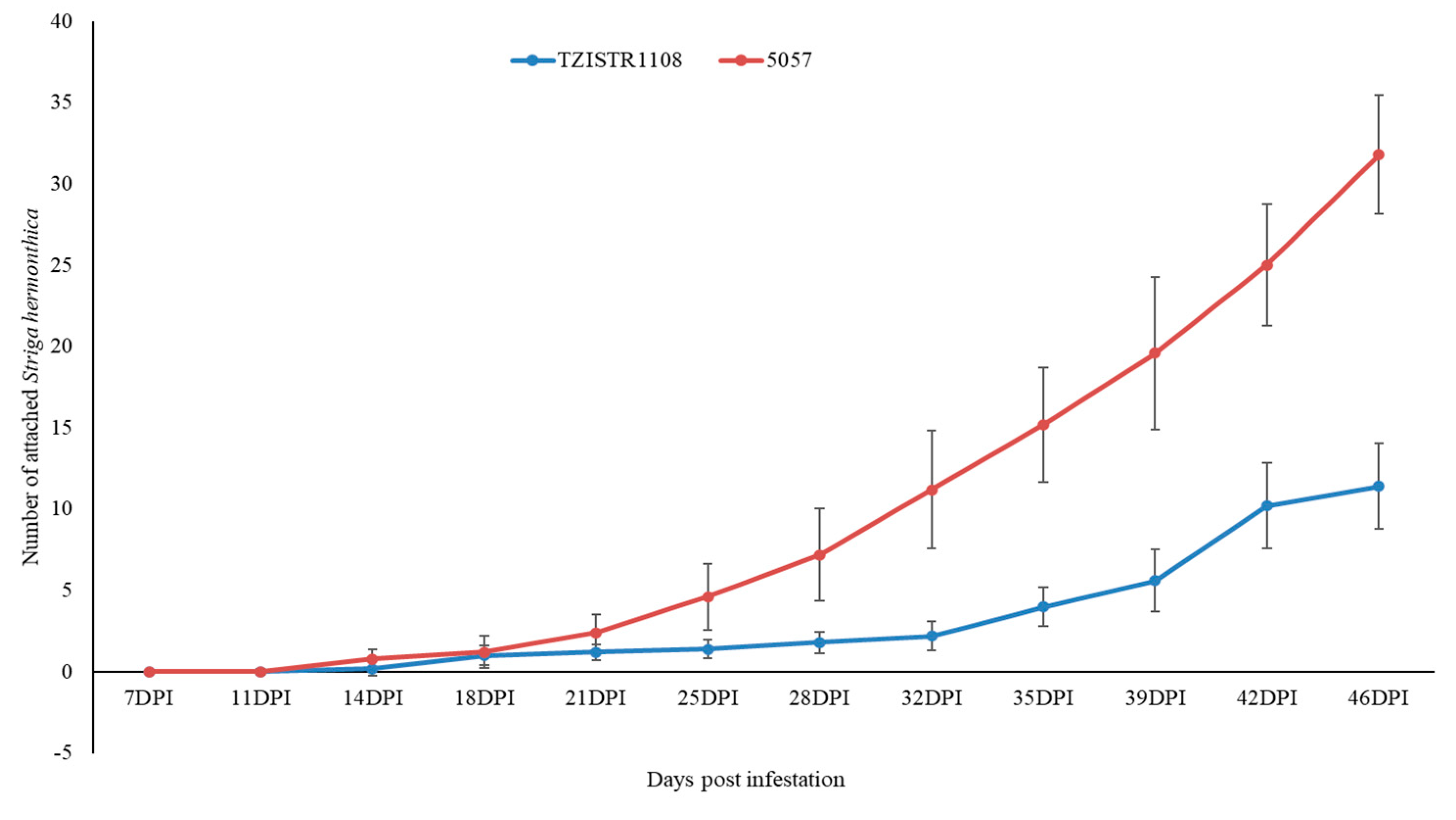
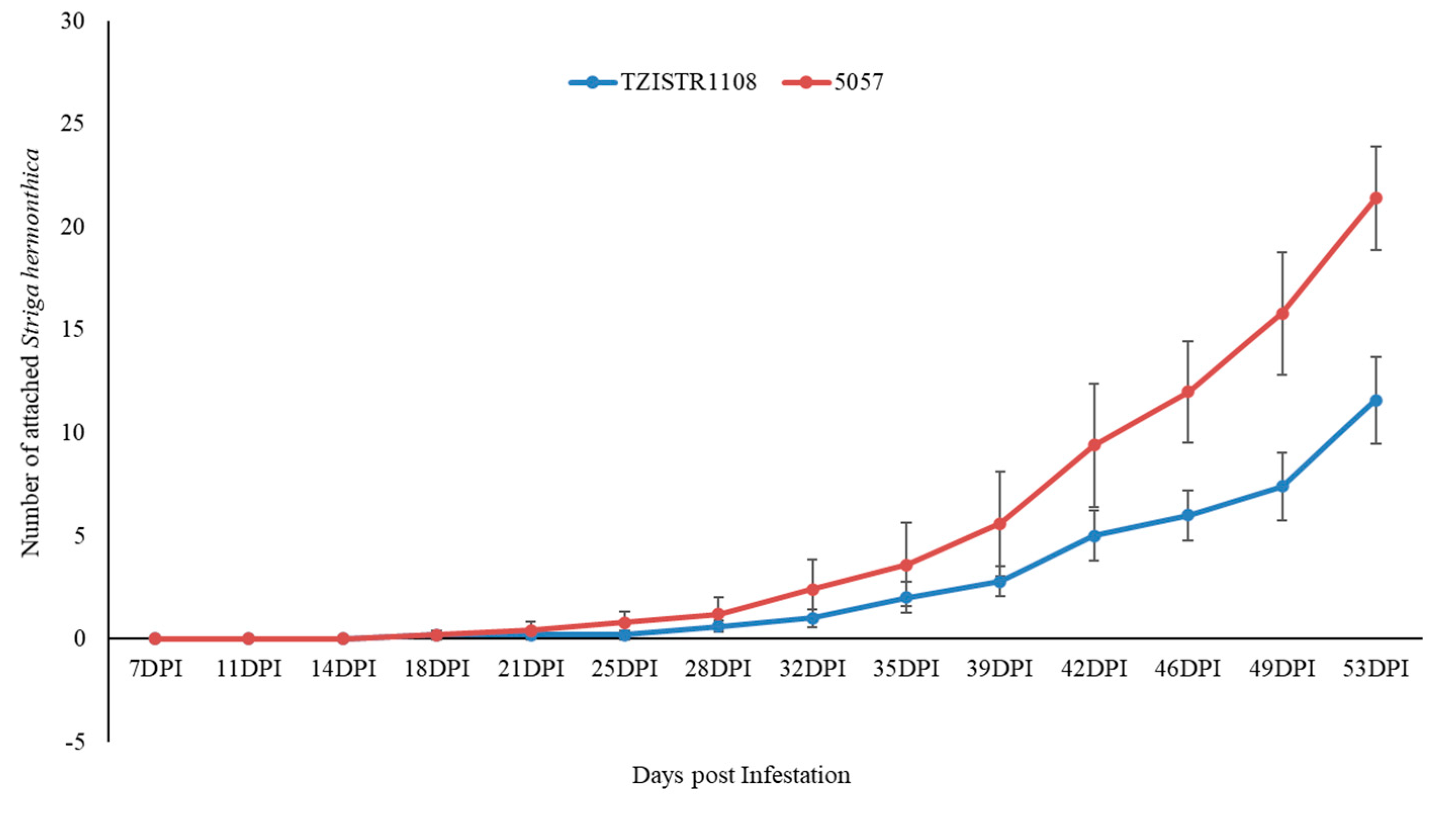
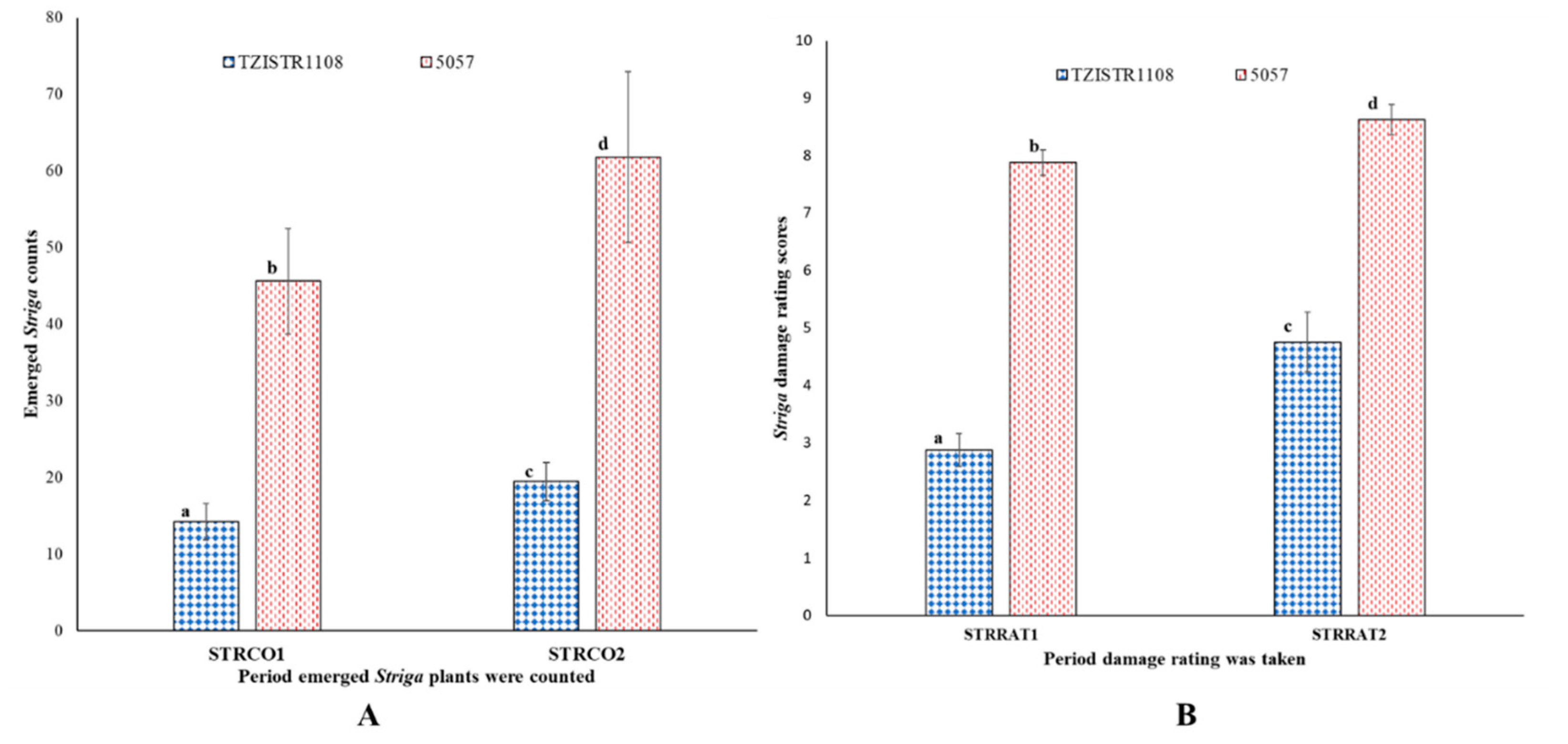
3.3.1. Evaluation of the Number of Attached S. hermonthica Plants
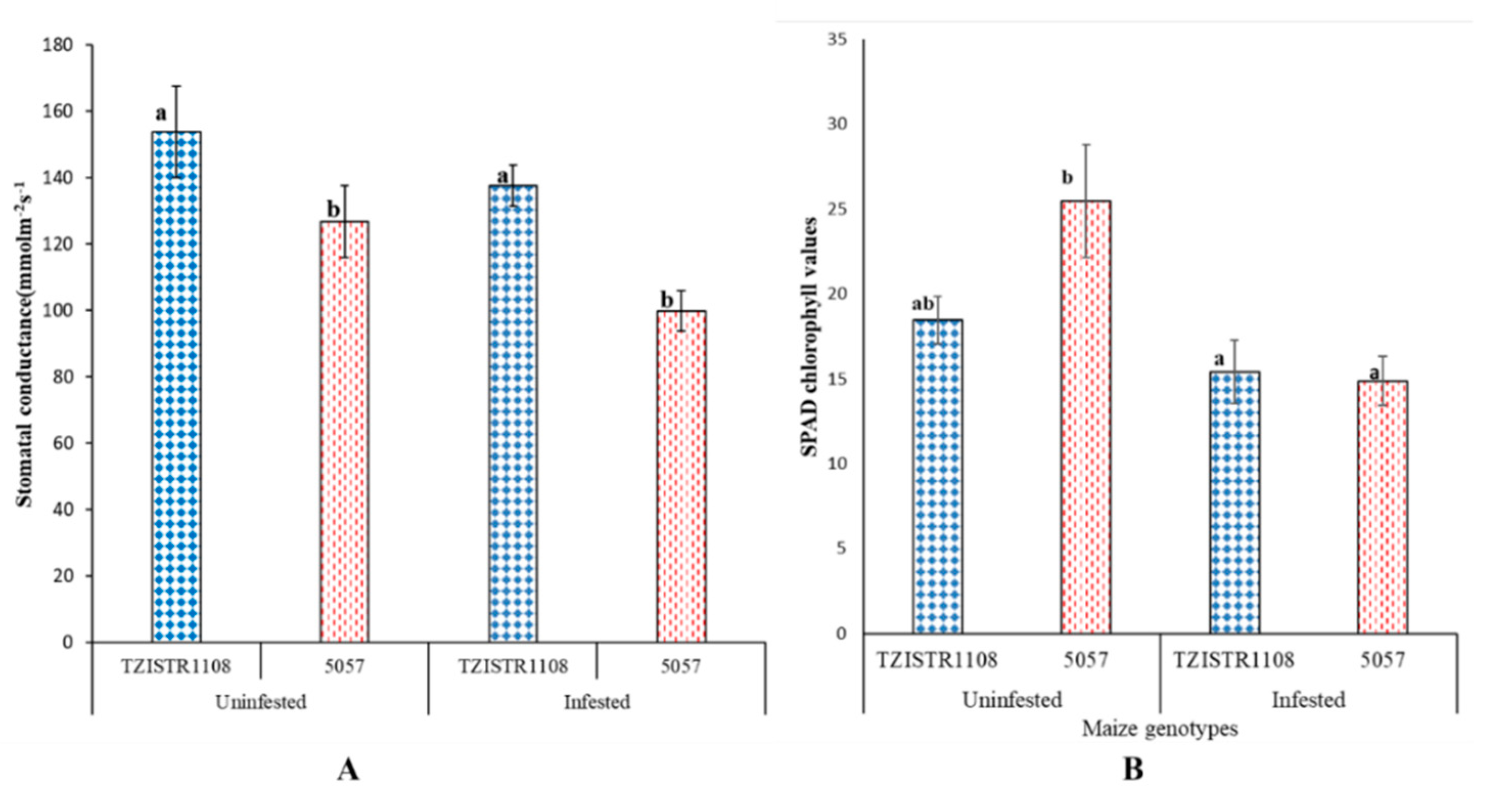
3.3.2. Physiological Changes Due to S. hermonthica Infestation
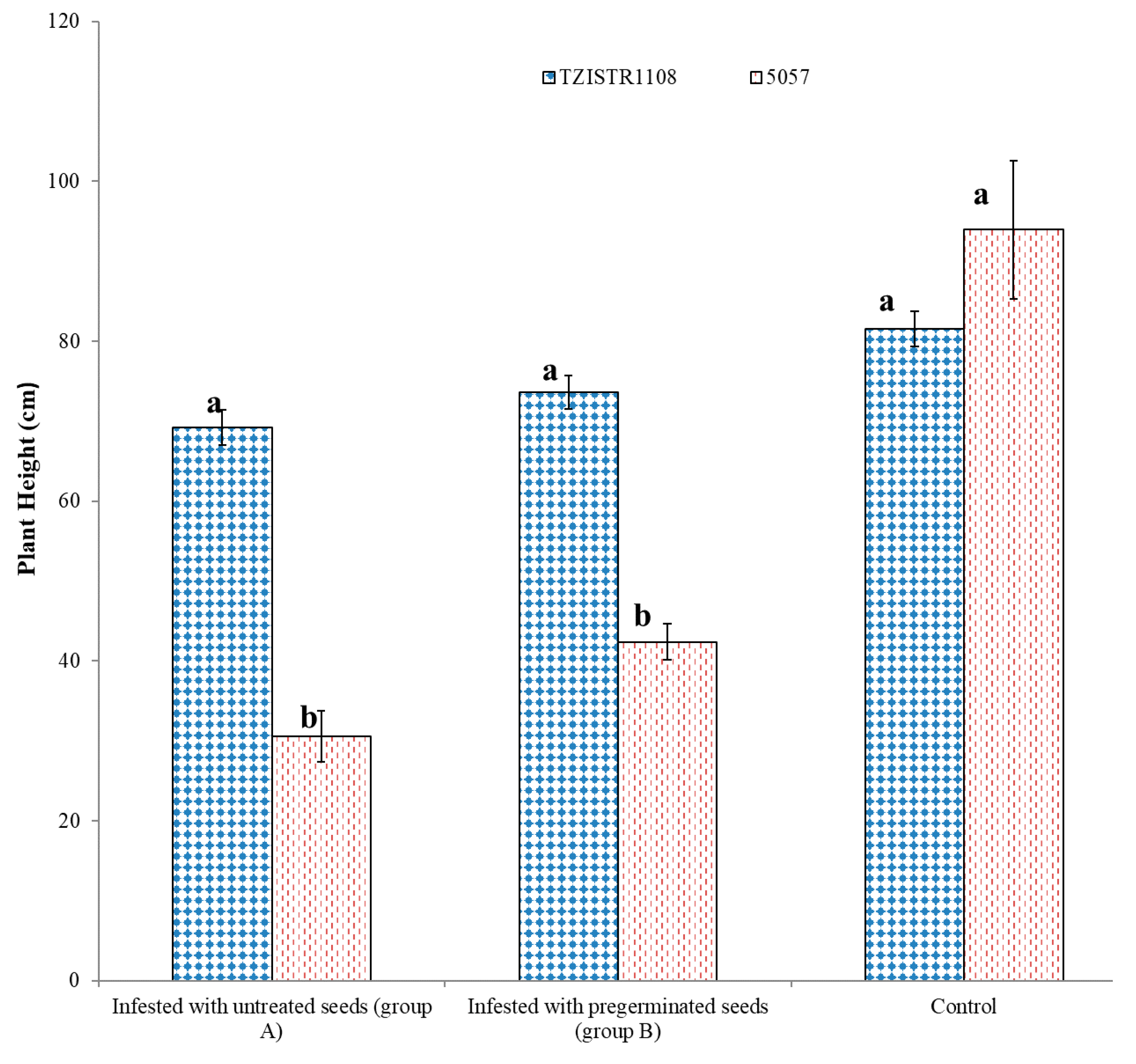
3.3.3. Effect of Parasitization on Plant Height
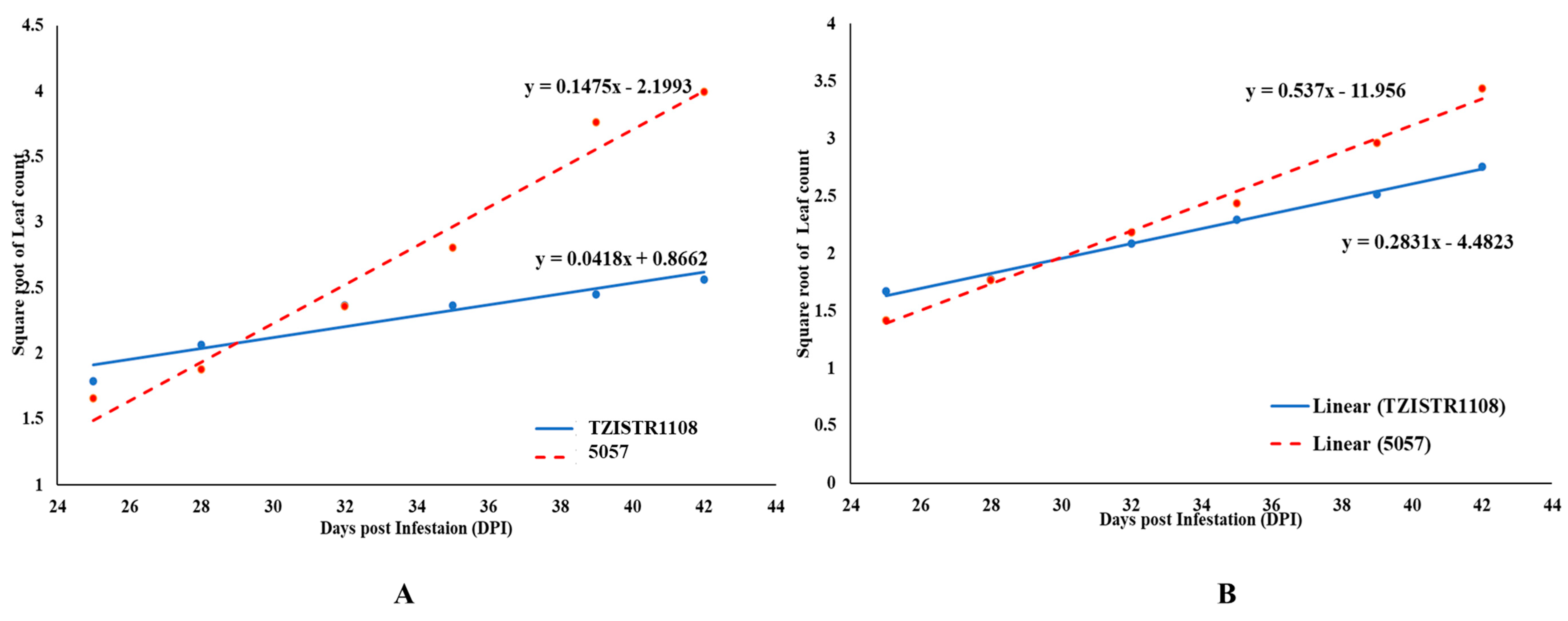
3.3.4. Growth Rate of Attached S. hermonthica Plants
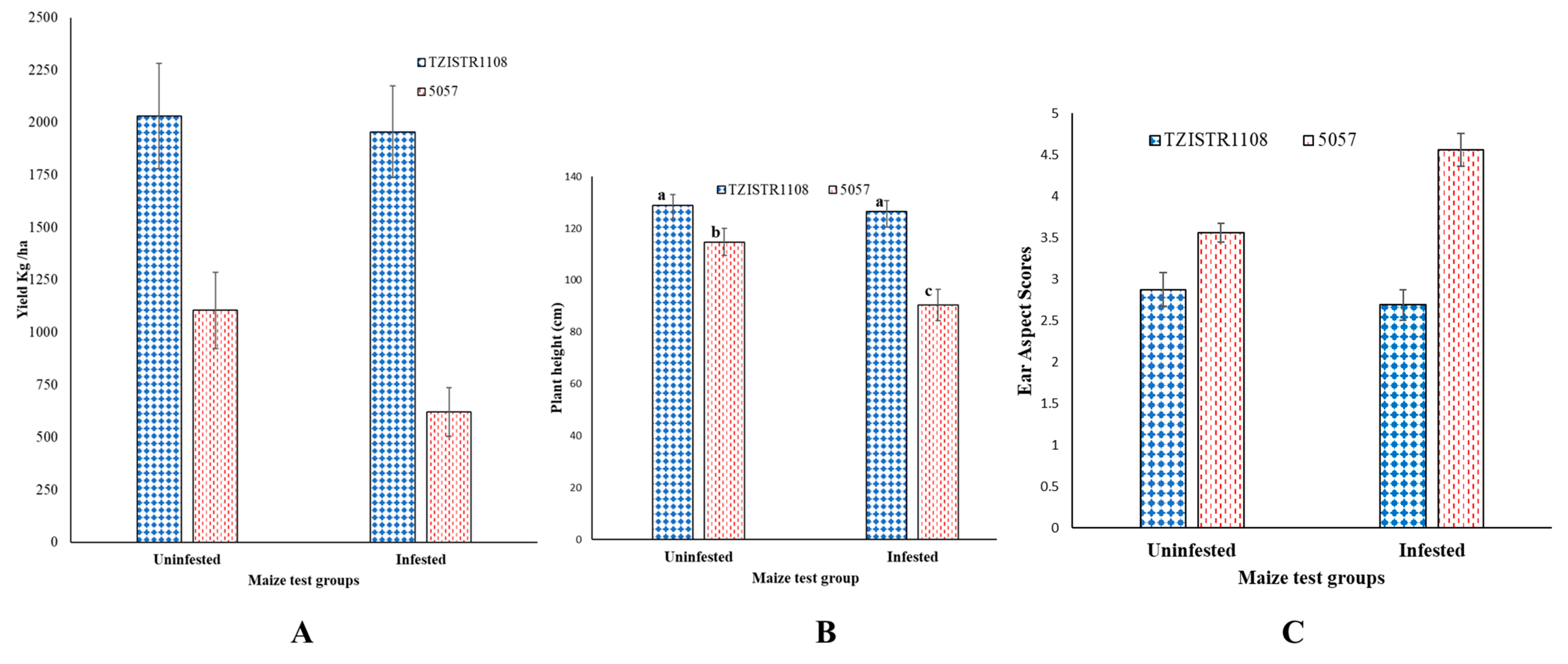
3.4. Field Evaluation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bozkurt, M.L.; Muth, P.; Parzies, H.K.; Haussmann, B.I.G. Genetic diversity of East and West African Striga hermonthica populations and virulence effects on a contrasting set of sorghum cultivars. Weed Res. 2014, 55, 71–81. [Google Scholar] [CrossRef]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, CT, USA, 1993. [Google Scholar]
- Hearne, S.J. Control—The Striga conundrum. Pest Manag. Sci. 2009, 65, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Press, M.C. How do the parasitic weeds Striga and Orobanche influence host carbon relations? Asp. Appl. Biol. 1995, 42, 63–70. [Google Scholar]
- Press, C.; Scholes, J.D.; Watling, J.R. Parasitic plants: Physiological and ecological interactions with their hosts. In Physiological Plant Ecology; Press, M.C., Scholes, J.D., Barker, M.G., Eds.; Blackwell Science: Oxford, UK, 1999; pp. 175–197. [Google Scholar]
- Frost, D.L.; Gurney, A.L.; Press, M.C.; Scholes, J.D. Striga hermonthica reduces photosynthesis in sorghum: The importance of stomatal limitations and a potential role for ABA? Plant Cell Environ. 1997, 20, 483–492. [Google Scholar] [CrossRef]
- Taylor, A.; Martin, J.; Seel, W.E. Physiology of the parasitic association between maize and witchweed (Striga hermonthica): Is ABA involved? J. Exp. Bot. 1996, 47, 1057–1065. [Google Scholar] [CrossRef]
- Parker, C. The Parasitic Weeds of the Orobanchaceae. In Parasitic Orobanchaceae. Parasitic Mechanisms and Control Strategies; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: New York, NY, USA, 2013; pp. 313–343. [Google Scholar]
- Graves, J.D.; Press, M.C.; Stewart, G.R. A carbon balance model of the sorghum-Striga hermonthica host-parasite association. Plant Cell Environ. 1989, 12, 101–107. [Google Scholar] [CrossRef]
- Eplee, R.E.; Norris, R. Control of Parasitic Weeds. Parasitic Plants; Press, M.C., Graves, J.D., Eds.; Chapman and Hall: London, UK, 1995; pp. 256–278. [Google Scholar]
- Kim, S.-K.; Adetimirin, V.O. Responses of Tolerant and Susceptible Maize Varieties to Timing and Rate of Nitrogen under Striga hermonthica Infestation. Agron. J. 1997, 89, 38–44. [Google Scholar] [CrossRef]
- Egley, G.H.; Eplee, R.E.; Norris, R.S. Discovery and development of ethylene as a witchweed seed germination stimulant. In Witchweed Research and Control in the U.S.; Sand, P.F., Eplee, R.E., Westbrooks, R.G., Eds.; WSSA: Champain, IL, USA, 1990; pp. 56–67. [Google Scholar]
- De Groote, H.; Wangare, L.; Kanampiu, F.; Odendo, M.; Diallo, A.; Karaya, H.; Friesen, D. The potential of a herbicide resistant maize technology for Striga control in Africa. Agric. Syst. 2008, 97, 83–94. [Google Scholar] [CrossRef]
- Parker, C. Protection of crops against parasitic weeds. Crop. Prot. 1991, 10, 6–22. [Google Scholar] [CrossRef]
- Kling, J.G.; Fajemisin, J.M.; Badu-Apraku, B.; Diallo, A.; Menkir, A.; Melake-Berhan, A. Striga resistance breeding in maize. In Breeding for Striga Resistance in Cereals; Haussmann, B.I.G., Hess, D.E., Koyama, M.L., Grivet, L., Rattunde, H.F.W., Geiger, H.H., Eds.; Margraf Verlag. Haussmann: Weikersheim, Germany, 2000; pp. 103–118. [Google Scholar]
- Kim, S. Genetics of Maize Tolerance of Striga hermonthica. Crop. Sci. 1994, 34, 900–907. [Google Scholar] [CrossRef]
- Riches, C.R.; Parker, C. Parasitic plants as weeds. In Press Parasitic; Plants, M.C., Graves, J.D., Eds.; Chapman and Hall: London, UK, 1995; pp. 226–255. [Google Scholar]
- Timko, M.P.; Scholes, J.D. Host Reaction to Attack by Root Parasitic Plants. In Parasitic Orobanchaceae; Joel, D.M., Gressel, J., Musselman, L.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 115–141. [Google Scholar] [CrossRef]
- Rodenburg, J.; Riches, C.R.; Kayeke, J.M. Addressing current and future problems of parasitic weeds in rice. Crop. Prot. 2010, 29, 210–221. [Google Scholar] [CrossRef]
- Rich, P.J.; Ejeta, G. Towards effective resistance to Striga in African maize. Plant Signal. Behav. 2008, 3, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A.; Makumbi, D.; Franco, J. Assessment of Reaction Patterns of Hybrids to Striga hermonthica (Del.) Benth. under Artificial Infestation in Kenya and Nigeria. Crop. Sci. 2012, 52, 2528–2537. [Google Scholar] [CrossRef]
- Menkir, A. Assessment of reactions of diverse maize inbred lines to Striga hermonthica (Del.) Benth. Plant Breed. 2006, 125, 131–139. [Google Scholar] [CrossRef]
- Menkir, A.; Kling, J.; Badu-Apraku, B.; Ibikunle, O. Registration of 26 Tropical Maize Germplasm Lines with Resistance to Striga hermonthica. Crop. Sci. 2006, 46, 1007–1009. [Google Scholar] [CrossRef]
- Amusan, I.O.; Rich, P.J.; Menkir, A.; Housley, T.; Ejeta, G. Resistance to Striga hermonthica in a maize inbred line derived from Zea diploperennis. New Phytol. 2008, 178, 157–166. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2018. Available online: https://www.r-project.org/ (accessed on 13 February 2012).
- Pierce, S.; Mbwaga, A.M.; Press, M.C.; Scholes, J.D. Xenognosin production and tolerance to Striga asiatica infection of high-yielding maize cultivars. Weed Res. 2003, 43, 139–145. [Google Scholar] [CrossRef]
- Gobena, D.; Shimels, M.; Rich, P.J.; Ruyter-Spira, C.; Bouwmeester, H.; Kanuganti, S.; Mengiste, T.; Ejeta, G. Mutation in Sorghum LOW GERMINATION STIMULANT 1 alters Strigolactones and Causes Striga resistance. Proc. Natl. Acad. Sci. USA 2017, 1149, 4471–4476. [Google Scholar] [CrossRef]
- Ejeta, G.; Gressel, J. Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt; World Scientific Pub. Co Pte Lt: Singapore, 2007; pp. 3–16. [Google Scholar]
- Cherif-Ari, O.; Housley, T.L.; Ejeta, G. Sorghum root length density and the potential for avoiding striga parasitism. Plant Soil 1990, 121, 67–72. [Google Scholar] [CrossRef]
- Van Delft, G.-J.; Graves, J.D.; Fitter, A.H.; Pruiksma, M.A. Spatial distribution and population dynamics of Striga hermonthica seeds in naturally infested farm soils. Plant Soil 1997, 195, 1–15. [Google Scholar] [CrossRef]
- Dodd, I.C. Hormonal Interactions and Stomatal Responses. J. Plant Growth Regul. 2003, 22, 32–46. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors Influencing Leaf Chlorophyll Content in Natural Forests at the Biome Scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef]
- Franić, M.; Galić, V.; Lončarić, Z.; Šimić, D. Genotypic Variability of Photosynthetic Parameters in Maize Ear-Leaves at Different Cadmium Levels in Soil. Agronomy 2020, 10, 986. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Unachukwu, N.N.; Menkir, A.; Stanley, A.; Farombi, E.O.; Gedil, M. Contrasting Response Mechanisms of Maize Lines to Striga hermonthica. Agriculture 2020, 10, 485. https://doi.org/10.3390/agriculture10100485
Unachukwu NN, Menkir A, Stanley A, Farombi EO, Gedil M. Contrasting Response Mechanisms of Maize Lines to Striga hermonthica. Agriculture. 2020; 10(10):485. https://doi.org/10.3390/agriculture10100485
Chicago/Turabian StyleUnachukwu, Nnanna N., Abebe Menkir, Adekemi Stanley, Ebenezer O. Farombi, and Melaku Gedil. 2020. "Contrasting Response Mechanisms of Maize Lines to Striga hermonthica" Agriculture 10, no. 10: 485. https://doi.org/10.3390/agriculture10100485
APA StyleUnachukwu, N. N., Menkir, A., Stanley, A., Farombi, E. O., & Gedil, M. (2020). Contrasting Response Mechanisms of Maize Lines to Striga hermonthica. Agriculture, 10(10), 485. https://doi.org/10.3390/agriculture10100485