Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Experimental Section
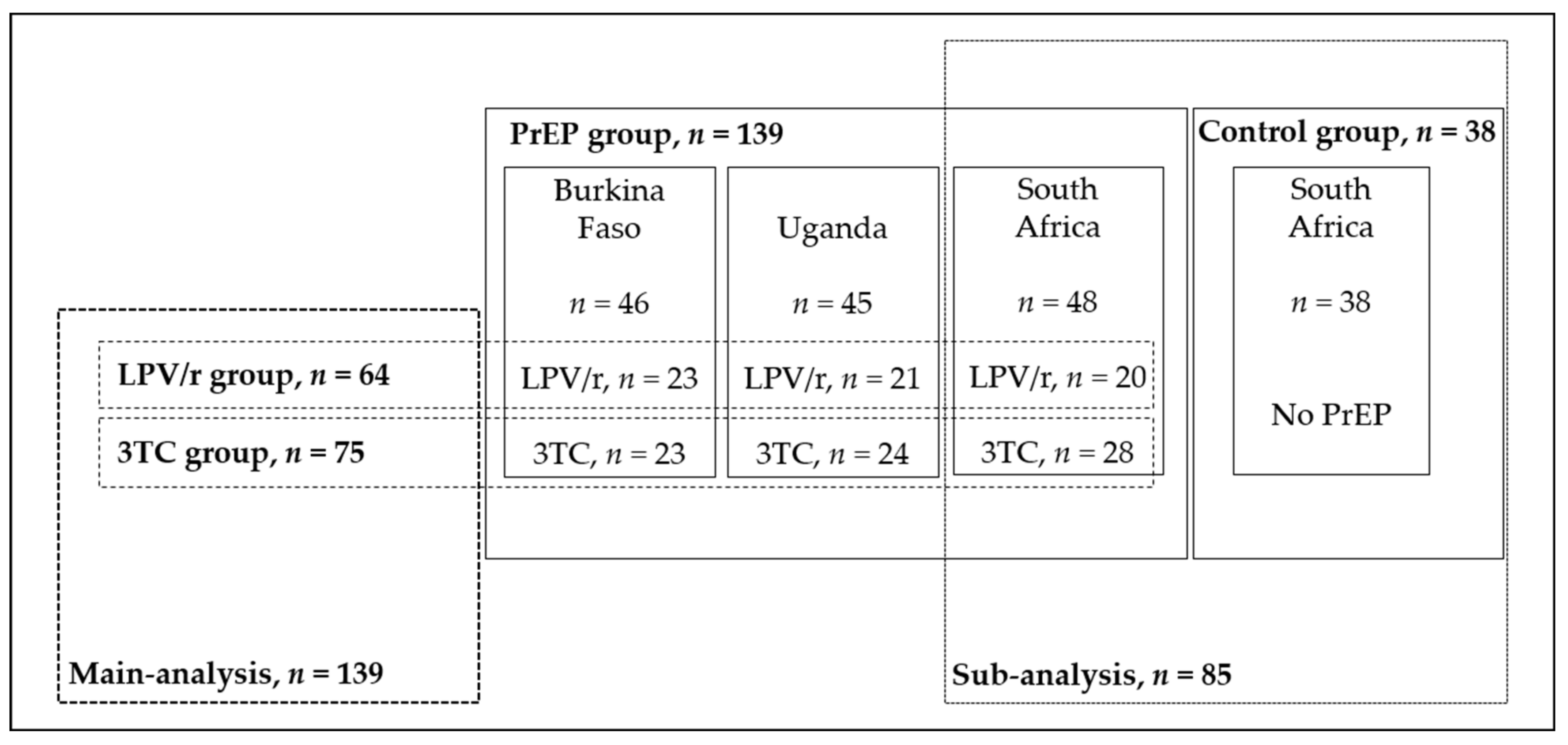
2.1. Design and Study Population
2.2. Sample Collection and Processing
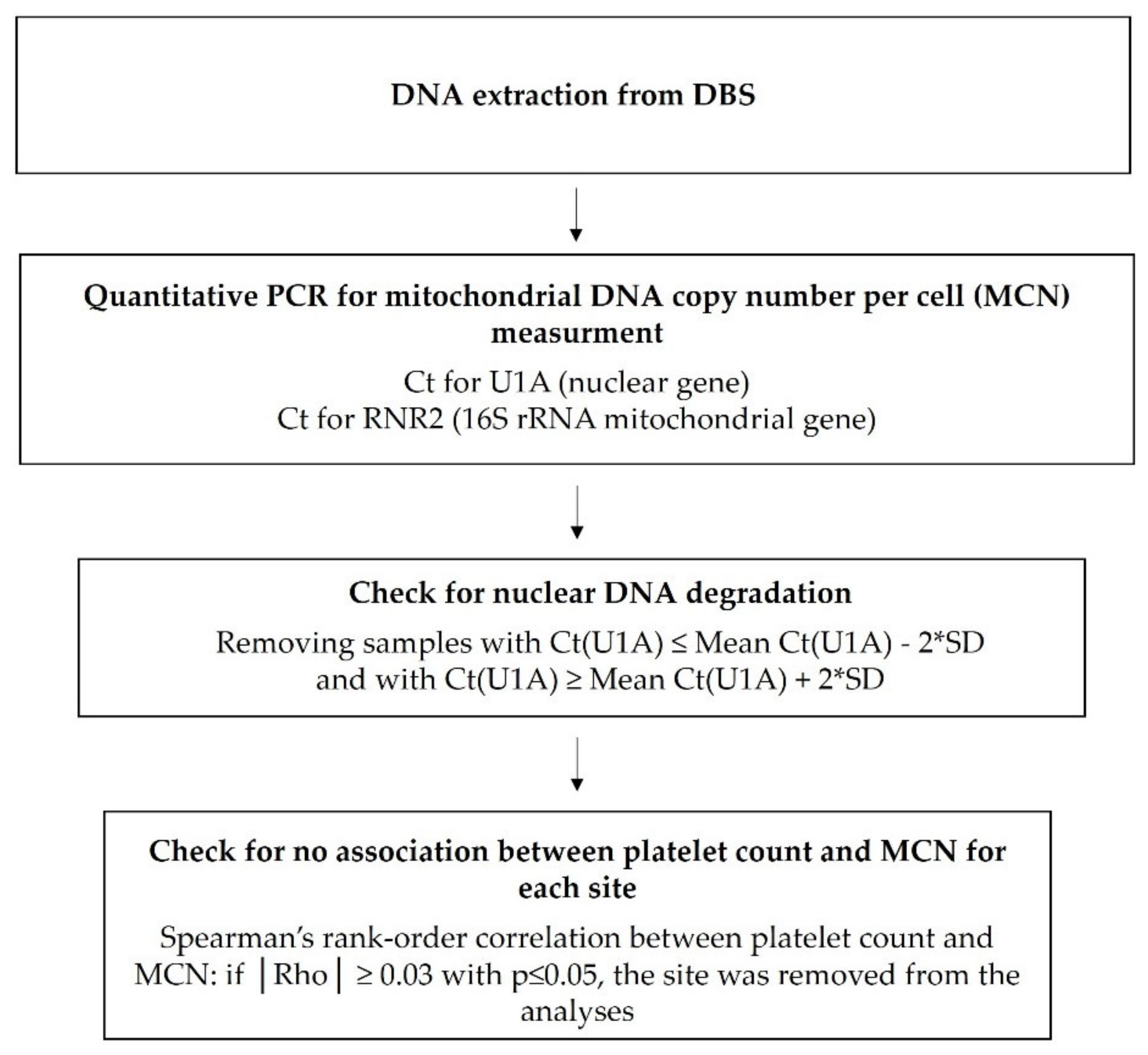
2.3. Mitochondrial DNA Parameters Assays
2.3.1. Mitochondrial DNA Copy Number Per Cell
2.3.2. Mitochondrial DNA Deletion
2.4. Statistical Analysis
2.5. Ethics Considerations
3. Results
3.1. Study Population Characteristics of the PrEP Group
3.2. mtDNA Content and Proportion of Deleted mtDNA at D7
3.3. mtDNA Content and Proportion of Deleted mtDNA after One Year of PrEP
3.4. Sub-Analysis of MCN and MDD Variations among CHEU from South Africa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Methodology Details for Mitochondrial DNA Parameters Assay
Appendix A.1. Mitochondrial DNA Copy Number Per Cell
Appendix A.2. Mitochondrial DNA Deletion
Appendix B. Methodology Details for the Validation of qPCR
References
- World Health Organization. Consolidated Guidelines on the Use of Antiretroviral Drugs for Treating and Preventing HIV Infection: Recommendations for a Public Health Approach, 2nd ed.; World Health Organization Press: Geneva, Switzerland, 2016; ISBN 978-92-4-154968-4. [Google Scholar]
- Ramírez-Ramírez, A.; Sánchez-Serrano, E.; Loaiza-Flores, G.; Plazola-Camacho, N.; Rodríguez-Delgado, R.G.; Figueroa-Damián, R.; Domínguez-Castro, M.; López-Martínez, M.; Flores-García, Z.; Hernández-Pineda, J. Simultaneous quantification of four antiretroviral drugs in breast milk samples from HIV-positive women by an ultra-high performance liquid chromatography tandem mass spectrometry (UPLC-MS/MS) method. PLoS ONE 2018, 13, e0191236. [Google Scholar] [CrossRef] [PubMed]
- Waitt, C.; Diliiy Penchala, S.; Olagunju, A.; Amara, A.; Else, L.; Lamorde, M.; Khoo, S. Development, validation and clinical application of a method for the simultaneous quantification of lamivudine, emtricitabine and tenofovir in dried blood and dried breast milk spots using LC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1060, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Salado-Rasmussen, K.; Theilgaard, Z.P.; Chiduo, M.G.; Bygbjerg, I.C.; Gerstoft, J.; Lüneborg-Nielsen, M.; Lemnge, M.; Katzenstein, T.L. Nevirapine, sodium concentration and HIV-1 RNA in breast milk and plasma among HIV-infected women receiving short-course antiretroviral prophylaxis. PLoS ONE 2015, 10, e0121111. [Google Scholar] [CrossRef] [PubMed]
- Mugwanya, K.K.; Hendrix, C.W.; Mugo, N.R.; Marzinke, M.; Katabira, E.T.; Ngure, K.; Semiyaga, N.B.; John-Stewart, G.; Muwonge, T.R.; Muthuri, G.; et al. Pre-exposure prophylaxis use by breastfeeding HIV-uninfected women: A prospective short-term study of antiretroviral excretion in breast milk and infant absorption. PLoS Med. 2016, 13, e1002132. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. Available online: http://aidsinfo.unaids.org/ (accessed on 27 July 2020).
- Williams, P.L.; Hazra, R.; Van Dyke, R.B.; Yildirim, C.; Crain, M.J.; Seage, G.R.; Civitello, L.; Ellis, A.; Butler, L.; Pediatric HIV/AIDS cohort study. Antiretroviral exposure during pregnancy and adverse outcomes in HIV-exposed uninfected infants and children using a trigger-based design. AIDS 2016, 30, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Warszawski, J.; Kariyawasam, D.; Le Chenadec, J.; Benhammou, V.; Czernichow, P.; Foissac, F.; Laborde, K.; Tréluyer, J.-M.; Firtion, G. Association of prenatal and postnatal exposure to lopinavir-ritonavir and adrenal dysfunction among uninfected infants of HIV-infected mothers. JAMA 2011, 306, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Newell, M.-L.; Coovadia, H.; Cortina-Borja, M.; Rollins, N.; Gaillard, P.; Dabis, F. Ghent international AIDS society (IAS) working group on HIV infection in women and children. Mortality of infected and uninfected infants born to HIV-infected mothers in Africa: A pooled analysis. Lancet 2004, 364, 1236–1243. [Google Scholar] [CrossRef]
- Taron-Brocard, C.; Le Chenadec, J.; Faye, A.; Dollfus, C.; Goetghebuer, T.; Gajdos, V.; Labaune, J.-M.; Perilhou, A.; Mandelbrot, L.; France REcherche Nord&Sud sida-HIV hepatitis—Enquete perinatale Francaise—CO1/CO11 study group; et al. Increased risk of serious bacterial infections due to maternal immunosuppression in HIV-exposed uninfected infants in a European country. Clin. Infect. Dis. 2014, 59, 1332–1345. [Google Scholar] [CrossRef]
- Arikawa, S.; Rollins, N.; Newell, M.-L.; Becquet, R. Mortality risk and associated factors in HIV-exposed, uninfected children. Trop. Med. Int. Health 2016, 21, 720–734. [Google Scholar] [CrossRef]
- Brennan, A.T.; Bonawitz, R.; Gill, C.J.; Thea, D.M.; Kleinman, M.; Useem, J.; Garrison, L.; Ceccarelli, R.; Udokwu, C.; Long, L.; et al. A meta-analysis assessing all-cause mortality in HIV-exposed uninfected compared with HIV-unexposed uninfected infants and children. AIDS 2016, 30, 2351–2360. [Google Scholar] [CrossRef]
- Omoni, A.O.; Ntozini, R.; Evans, C.; Prendergast, A.J.; Moulton, L.H.; Christian, P.S.; Humphrey, J.H. Child growth according to maternal and child HIV status in Zimbabwe. Pediatr. Infect. Dis. J. 2017, 36, 869–876. [Google Scholar] [CrossRef]
- Uthman, O.A.; Nachega, J.B.; Anderson, J.; Kanters, S.; Mills, E.J.; Renaud, F.; Essajee, S.; Doherty, M.C.; Mofenson, L.M. Timing of initiation of antiretroviral therapy and adverse pregnancy outcomes: A systematic review and meta-analysis. Lancet HIV 2017, 4, e21–e30. [Google Scholar] [CrossRef]
- Le Roux, S.M.; Abrams, E.J.; Donald, K.A.; Brittain, K.; Phillips, T.K.; Nguyen, K.K.; Zerbe, A.; Kroon, M.; Myer, L. Growth trajectories of breastfed HIV-exposed uninfected and HIV-unexposed children under conditions of universal maternal antiretroviral therapy: A prospective study. Lancet Child Adolesc. Health 2019, 3, 234–244. [Google Scholar] [CrossRef]
- Blanche, S.; Tylleskär, T.; Peries, M.; Kankasa, C.; Engebretsen, I.; Meda, N.; Tumwine, J.K.; Singata-Madliki, M.; Mwiya, M.; Van de Perre, P.; et al. Growth in HIV-1-exposed but uninfected infants treated with lopinavir-ritonavir versus lamivudine: A secondary analysis of the ANRS 12174 trial. Lancet HIV 2019, 6, e307–e314. [Google Scholar] [CrossRef]
- Wedderburn, C.J.; Evans, C.; Yeung, S.; Gibb, D.M.; Donald, K.A.; Prendergast, A.J. Growth and neurodevelopment of HIV-exposed uninfected children: A conceptual framework. Curr. HIV/AIDS Rep. 2019, 16, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Powis, K.M.; Smeaton, L.; Hughes, M.D.; Tumbare, E.A.; Souda, S.; Jao, J.; Wirth, K.E.; Makhema, J.; Lockman, S.; Fawzi, W.; et al. In-utero triple antiretroviral exposure associated with decreased growth among HIV-exposed uninfected infants in Botswana. AIDS 2016, 30, 211–220. [Google Scholar] [CrossRef]
- McHenry, M.S.; McAteer, C.I.; Oyungu, E.; McDonald, B.C.; Bosma, C.B.; Mpofu, P.B.; Deathe, A.R.; Vreeman, R.C. Neurodevelopment in young children born to HIV-infected mothers: A meta-analysis. Pediatrics 2018, 141, e20172888. [Google Scholar] [CrossRef] [PubMed]
- Le Roux, S.M.; Donald, K.A.; Brittain, K.; Phillips, T.K.; Zerbe, A.; Nguyen, K.K.; Strandvik, A.; Kroon, M.; Abrams, E.J.; Myer, L. Neurodevelopment of breastfed HIV-exposed uninfected and HIV-unexposed children in South Africa. AIDS 2018, 32, 1781–1791. [Google Scholar] [CrossRef]
- Wedderburn, C.J.; Yeung, S.; Rehman, A.M.; Stadler, J.A.M.; Nhapi, R.T.; Barnett, W.; Myer, L.; Gibb, D.M.; Zar, H.J.; Stein, D.J.; et al. Neurodevelopment of HIV-exposed uninfected children in South Africa: Outcomes from an observational birth cohort study. Lancet Child Adolesc. Health 2019, 3, 803–813. [Google Scholar] [CrossRef]
- Bunders, M.; Pembrey, L.; Kuijpers, T.; Newell, M.-L. Evidence of impact of maternal HIV infection on immunoglobulin levels in HIV-exposed uninfected children. AIDS Res. Hum. Retrovir. 2010, 26, 967–975. [Google Scholar] [CrossRef]
- Reikie, B.A.; Adams, R.C.M.; Leligdowicz, A.; Ho, K.; Naidoo, S.; Ruck, C.E.; de Beer, C.; Preiser, W.; Cotton, M.F.; Speert, D.P.; et al. Altered innate immune development in HIV-exposed uninfected infants. J. Acquir. Immune Defic. Syndr. 2014, 66, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Afran, L.; Garcia Knight, M.; Nduati, E.; Urban, B.C.; Heyderman, R.S.; Rowland-Jones, S.L. HIV-exposed uninfected children: A growing population with a vulnerable immune system? Clin. Exp. Immunol. 2014, 176, 11–22. [Google Scholar] [CrossRef]
- Gianesin, K.; Noguera-Julian, A.; Zanchetta, M.; Del Bianco, P.; Petrara, M.R.; Freguja, R.; Rampon, O.; Fortuny, C.; Camós, M.; Mozzo, E.; et al. Premature aging and immune senescence in HIV-infected children. AIDS 2016, 30, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raya, B.; Kollmann, T.R.; Marchant, A.; MacGillivray, D.M. The immune system of HIV-exposed uninfected infants. Front. Immunol. 2016, 7, 383. [Google Scholar] [CrossRef]
- Poirier, M.C.; Gibbons, A.T.; Rugeles, M.T.; Andre-Schmutz, I.; Blanche, S. Fetal consequences of maternal antiretroviral nucleoside reverse transcriptase inhibitor (NRTI) use in human and non-human primate pregnancy. Curr. Opin. Pediatr. 2015, 27, 233–239. [Google Scholar] [CrossRef]
- Noguera-Julian, A.; Morén, C.; Rovira, N.; Garrabou, G.; Catalán, M.; Sánchez, E.; Cardellach, F.; Miró, Ó.; Fortuny, C. Decreased mitochondrial function among healthy infants exposed to antiretrovirals during gestation, delivery and the neonatal period. Pediatr. Infect. Dis. J. 2015, 34, 1349–1354. [Google Scholar] [CrossRef]
- Jao, J.; Powis, K.M.; Kirmse, B.; Yu, C.; Epie, F.; Nshom, E.; Abrams, E.J.; Sperling, R.S.; Leroith, D.; Geffner, M.E.; et al. Lower mitochondrial DNA and altered mitochondrial fuel metabolism in HIV-exposed uninfected infants in Cameroon. AIDS 2017, 31, 2475–2481. [Google Scholar] [CrossRef]
- Eckard, A.R.; Kirk, S.E.; Hagood, N.L. Contemporary issues in pregnancy (and offspring) in the current HIV era. Curr. HIV/AIDS Rep 2019, 16, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Powis, K.M.; Siberry, G.K. Optimising HIV antiretrovirals for mothers and infants. Lancet HIV 2019, 6, e272–e273. [Google Scholar] [CrossRef]
- Koczor, C.A.; Lewis, W. Nucleoside reverse transcriptase inhibitor toxicity and mitochondrial DNA. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1493–1504. [Google Scholar] [CrossRef]
- Margolis, A.M.; Heverling, H.; Pham, P.A.; Stolbach, A. A review of the toxicity of HIV medications. J. Med. Toxicol. 2014, 10, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Kakuda, T.N. Pharmacology of nucleoside and nucleotide reverse transcriptase inhibitor-induced mitochondrial toxicity. Clin. Ther. 2000, 22, 685–708. [Google Scholar] [CrossRef]
- Estaquier, J.; Lelièvre, J.-D.; Petit, F.; Brunner, T.; Moutouh-de Parseval, L.; Richman, D.D.; Ameisen, J.C.; Corbeil, J. Effects of antiretroviral drugs on human immunodeficiency virus type 1-induced CD4+ T-cell death. J. Virol. 2002, 76, 5966–5973. [Google Scholar] [CrossRef]
- Tricarico, P.M.; De Oliveira Franca, R.F.; Pacor, S.; Ceglia, V.; Crovella, S.; Celsi, F. HIV protease inhibitors apoptotic effect in SH-SY5Y neuronal cell line. Cell Physiol. Biochem. 2016, 39, 1463–1470. [Google Scholar] [CrossRef]
- Ganta, K.K.; Chaubey, B. Endoplasmic reticulum stress leads to mitochondria-mediated apoptosis in cells treated with anti-HIV protease inhibitor ritonavir. Cell Biol. Toxicol. 2019, 35, 189–204. [Google Scholar] [CrossRef]
- Taura, M.; Kariya, R.; Kudo, E.; Goto, H.; Iwawaki, T.; Amano, M.; Suico, M.A.; Kai, H.; Mitsuya, H.; Okada, S. Comparative analysis of ER stress response into HIV protease inhibitors: Lopinavir but not darunavir induces potent ER stress response via ROS/JNK pathway. Free Radic. Biol. Med. 2013, 65, 778–788. [Google Scholar] [CrossRef]
- Xiang, T.; Du, L.; Pham, P.; Zhu, B.; Jiang, S. Nelfinavir, an HIV protease inhibitor, induces apoptosis and cell cycle arrest in human cervical cancer cells via the ROS-dependent mitochondrial pathway. Cancer Lett. 2015, 364, 79–88. [Google Scholar] [CrossRef]
- Gratton, R.; Tricarico, P.M.; Guimaraes, R.L.; Celsi, F.; Crovella, S. Lopinavir/Ritonavir treatment induces oxidative stress and caspase independent apoptosis in human glioblastoma U-87 MG cell line. Curr. HIV Res. 2018, 16, 106–112. [Google Scholar] [CrossRef]
- Morén, C.; Noguera-Julian, A.; Garrabou, G.; Catalán, M.; Rovira, N.; Tobías, E.; Cardellach, F.; Miró, Ò.; Fortuny, C. Mitochondrial evolution in HIV-infected children receiving first- or second-generation nucleoside analogues. J. Acquir. Immune Defic. Syndr. 2012, 60, 111–116. [Google Scholar] [CrossRef]
- Liu, K.; Sun, Y.; Liu, D.; Yin, J.; Qiao, L.; Shi, Y.; Dong, Y.; Li, N.; Zhang, F.; Chen, D. Mitochondrial toxicity studied with the PBMC of children from the Chinese national pediatric highly active antiretroviral therapy cohort. PLoS ONE 2013, 8, e57223. [Google Scholar] [CrossRef]
- Shen, J.; Liberty, A.; Shiau, S.; Strehlau, R.; Pierson, S.; Patel, F.; Wang, L.; Burke, M.; Violari, A.; Coovadia, A.; et al. Mitochondrial impairment in well-suppressed children with perinatal HIV-infection on antiretroviral therapy. AIDS Res. Hum. Retrovir. 2019, 36, 27–38. [Google Scholar] [CrossRef]
- Poirier, M.C.; Divi, R.L.; Al-Harthi, L.; Olivero, O.A.; Nguyen, V.; Walker, B.; Landay, A.L.; Walker, V.E.; Charurat, M.; Women and Infants Transmission Study (WITS) Group. Long-term mitochondrial toxicity in HIV-uninfected infants born to HIV-infected mothers. J. Acquir. Immune Defic. Syndr. 2003, 33, 175–183. [Google Scholar] [CrossRef]
- Divi, R.L.; Walker, V.E.; Wade, N.A.; Nagashima, K.; Seilkop, S.K.; Adams, M.E.; Nesel, C.J.; O’Neill, J.P.; Abrams, E.J.; Poirier, M.C. Mitochondrial damage and DNA depletion in cord blood and umbilical cord from infants exposed in utero to Combivir. AIDS 2004, 18, 1013–1021. [Google Scholar] [CrossRef]
- Divi, R.L.; Leonard, S.L.; Kuo, M.M.; Nagashima, K.; Thamire, C.; St. Claire, M.C.; Wade, N.A.; Walker, V.E.; Poirier, M.C. Transplacentally exposed human and monkey newborn infants show similar evidence of nucleoside reverse transcriptase inhibitor-induced mitochondrial toxicity. Environ. Mol. Mutagen. 2007, 48, 201–209. [Google Scholar] [CrossRef]
- Hernàndez, S.; Morén, C.; López, M.; Coll, O.; Cardellach, F.; Gratacós, E.; Miró, O.; Garrabou, G. Perinatal outcomes, mitochondrial toxicity and apoptosis in HIV-treated pregnant women and in-utero-exposed newborn. AIDS 2012, 26, 419–428. [Google Scholar] [CrossRef]
- Aldrovandi, G.M.; Chu, C.; Shearer, W.T.; Li, D.; Walter, J.; Thompson, B.; McIntosh, K.; Foca, M.; Meyer, W.A.; Ha, B.F.; et al. Antiretroviral exposure and lymphocyte mtDNA content among uninfected infants of HIV-1-infected women. Pediatrics 2009, 124, e1189–e1197. [Google Scholar] [CrossRef]
- McComsey, G.A.; Kang, M.; Ross, A.C.; Lebrecht, D.; Livingston, E.; Melvin, A.; Hitti, J.; Cohn, S.E.; Walker, U.A.; AIDS Clinical Trials Group A5084. Increased mtDNA levels without change in mitochondrial enzymes in peripheral blood mononuclear cells of infants born to HIV-infected mothers on antiretroviral therapy. HIV Clin. Trials 2008, 9, 126–136. [Google Scholar] [CrossRef]
- Côté, H.C.F.; Raboud, J.; Bitnun, A.; Alimenti, A.; Money, D.M.; Maan, E.; Costei, A.; Gadawski, I.; Diong, C.; Read, S.; et al. Perinatal exposure to antiretroviral therapy is associated with increased blood mitochondrial DNA levels and decreased mitochondrial gene expression in infants. J. Infect. Dis. 2008, 198, 851–859. [Google Scholar] [CrossRef]
- Ross, A.C.; Leong, T.; Avery, A.; Castillo-Duran, M.; Bonilla, H.; Lebrecht, D.; Walker, U.A.; Storer, N.; Labbato, D.; Khaitan, A.; et al. Effects of in utero antiretroviral exposure on mitochondrial DNA levels, mitochondrial function and oxidative stress. HIV Med. 2012, 13, 98–106. [Google Scholar] [CrossRef]
- Ajaykumar, A.; Zhu, M.; Kakkar, F.; Brophy, J.; Bitnun, A.; Alimenti, A.; Soudeyns, H.; Saberi, S.; Albert, A.Y.K.; CIHR team in cellular aging and HIV comorbidities in women and children (CARMA); et al. Blood mitochondrial DNA levels remain elevated from birth to early life in children HIV-exposed uninfected exposed to combination antiretroviral therapy in utero. J. Infect. Dis. Online ahead of print. [CrossRef]
- Schon, E.A.; DiMauro, S.; Hirano, M. Human mitochondrial DNA: Roles of inherited and somatic mutations. Nat. Rev. Genet. 2012, 13, 878–890. [Google Scholar] [CrossRef] [PubMed]
- Kazachkova, N. Mitochondrial DNA damage patterns and aging: Revising the evidences for humans and mice. Aging Dis. 2013, 4, 337–350. [Google Scholar] [CrossRef]
- Brogly, S.B.; DiMauro, S.; Van Dyke, R.B.; Williams, P.L.; Naini, A.; Libutti, D.E.; Choi, J.; Chung, M.; Gerschenson, M. Short communication: Transplacental nucleoside analogue exposure and mitochondrial parameters in HIV-uninfected children. AIDS Res. Hum. Retrovir. 2011, 27, 777–783. [Google Scholar] [CrossRef]
- Kunz, A.; von Wurmb-Schwark, N.; Sewangi, J.; Ziske, J.; Lau, I.; Mbezi, P.; Theuring, S.; Hauser, A.; Dugange, F.; Katerna, A.; et al. Zidovudine exposure in HIV-1 infected Tanzanian women increases mitochondrial DNA levels in placenta and umbilical cords. PLoS ONE 2012, 7, e41637. [Google Scholar] [CrossRef]
- Nagot, N.; Kankasa, C.; Meda, N.; Hofmeyr, J.; Nikodem, C.; Tumwine, J.K.; Karamagi, C.; Sommerfelt, H.; Neveu, D.; PROMISE-PEP group; et al. Lopinavir/Ritonavir versus Lamivudine peri-exposure prophylaxis to prevent HIV-1 transmission by breastfeeding: The PROMISE-PEP trial Protocol ANRS 12174. BMC Infect. Dis. 2012, 12, 246. [Google Scholar] [CrossRef]
- Nagot, N.; Kankasa, C.; Tumwine, J.K.; Meda, N.; Hofmeyr, G.J.; Vallo, R.; Mwiya, M.; Kwagala, M.; Traore, H.; Sunday, A.; et al. Extended pre-exposure prophylaxis with lopinavir-ritonavir versus lamivudine to prevent HIV-1 transmission through breastfeeding up to 50 weeks in infants in Africa (ANRS 12174): A randomised controlled trial. Lancet 2016, 387, 566–573. [Google Scholar] [CrossRef]
- World Health Organization. Antiretroviral Drugs for Treating Pregnant Women and Preventing HIV Infection in Infant: Towards Universal Access: Recommendations for a Public Health Approach; World Health Organization Press: Geneva, Switzerland, 2006; ISBN 978-92-4-159466-0. [Google Scholar]
- 2012–2013 SAPMTCTE Report. Early (4–8 Weeks Postpartum) Population-Level Effectiveness of WHO PMTCT Option A, South Africa. Available online: https://www.samrc.ac.za/sites/default/files/files/2016-07-12/SAPMTCTEReport2012.pdf (accessed on 25 August 2020).
- Timmermans, E.C.; Tebas, P.; Ruiter, J.P.N.; Wanders, R.J.A.; de Ronde, A.; de Baar, M.P. Real-time nucleic acid sequence-based amplification assay to quantify changes in mitochondrial DNA concentrations in cell cultures and blood cells from HIV-infected patients receiving antiviral therapy. Clin. Chem. 2006, 52, 979–987. [Google Scholar] [CrossRef][Green Version]
- Ashar, F.N.; Moes, A.; Moore, A.Z.; Grove, M.L.; Chaves, P.H.M.; Coresh, J.; Newman, A.B.; Matteini, A.M.; Bandeen-Roche, K.; Boerwinkle, E.; et al. Association of mitochondrial DNA levels with frailty and all-cause mortality. J. Mol. Med. 2015, 93, 177–186. [Google Scholar] [CrossRef]
- Hurtado-Roca, Y.; Ledesma, M.; Gonzalez-Lazaro, M.; Moreno-Loshuertos, R.; Fernandez-Silva, P.; Enriquez, J.A.; Laclaustra, M. Adjusting MtDNA quantification in whole blood for peripheral blood platelet and leukocyte counts. PLoS ONE 2016, 11, e0163770. [Google Scholar] [CrossRef]
- Xia, C.-Y.; Liu, Y.; Yang, H.-R.; Yang, H.-Y.; Liu, J.-X.; Ma, Y.-N.; Qi, Y. Reference intervals of mitochondrial DNA copy number in peripheral blood for Chinese minors and adults. Chin. Med. J. 2017, 130, 2435–2440. [Google Scholar] [CrossRef]
- Phillips, N.R.; Sprouse, M.L.; Roby, R.K. Simultaneous quantification of mitochondrial DNA copy number and deletion ratio: A multiplex real-time PCR assay. Sci. Rep. 2014, 4, 3887. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Tan, J.M.E.; Jonker, M.J.; Jongejan, A.; Buissink, T.; Veldhuijzen, S.; van Kampen, A.H.C.; Brul, S.; van der Spek, H. Beyond the polymerase-γ theory: Production of ROS as a mode of NRTI-induced mitochondrial toxicity. PLoS ONE 2017, 12, e0187424. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Maagaard, A.; Holberg-Petersen, M.; Kollberg, G.; Oldfors, A.; Sandvik, L.; Bruun, J.N. Mitochondrial (mt)DNA changes in tissue may not be reflected by depletion of mtDNA in peripheral blood mononuclear cells in HIV-infected patients. Antivir. Ther. 2006, 11, 601–608. [Google Scholar]
- Lawless, C.; Greaves, L.; Reeve, A.K.; Turnbull, D.M.; Vincent, A.E. The rise and rise of mitochondrial DNA mutations. Open Biol. 2020, 10, 200061. [Google Scholar] [CrossRef]
- Côté, H.C.F.; Brumme, Z.L.; Craib, K.J.P.; Alexander, C.S.; Wynhoven, B.; Ting, L.; Wong, H.; Harris, M.; Harrigan, P.R.; O’Shaughnessy, M.; et al. Changes in mitochondrial DNA as a marker of nucleoside toxicity in HIV-infected patients. N. Engl. J. Med. 2002, 346, 811–820. [Google Scholar] [CrossRef]
- Sun, J.; Longchamps, R.J.; Piggott, D.A.; Castellani, C.A.; Sumpter, J.A.; Brown, T.T.; Mehta, S.H.; Arking, D.E.; Kirk, G.D. Association between HIV infection and mitochondrial DNA copy number in peripheral blood: A population-based, prospective cohort study. J. Infect. Dis. 2019, 219, 1285–1293. [Google Scholar] [CrossRef]
- Tourmen, Y.; Baris, O.; Dessen, P.; Jacques, C.; Malthièry, Y.; Reynier, P. Structure and chromosomal distribution of human mitochondrial pseudogenes. Genomics 2002, 80, 71–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Characteristic | LPV/r (n = 64) | 3TC (n = 75) | Total (n = 139) | p Value a |
|---|---|---|---|---|
| Gender; n (%) | 0.96 | |||
| Male | 33 (51.6) | 39 (52.0) | 72 (51.8) | |
| Weight (kg); mean ± SD | 3.1 ± 0.5 | 3.3 ± 0.4 | 3.2 ± 0.5 | 0.19 |
| Height (cm); mean ± SD | 49.2 ± 2.1 | 49.8 ± 2.0 | 49.5 ± 2.0 | 0.11 |
| Gestational age (week); mean ± SD | 38.3 ± 1.7 | 38.5 ± 1.9 | 38.4 ± 1.8 | 0.30 |
| Preterm birth (week); n (%) | 0.82 | |||
| No prematurity ≥ 37 | 53 (82.8) | 61 (81.3) | 114 (82.0) | |
| Prematurity < 37 | 11 (17.2) | 14 (18.7) | 25 (18.0) | |
| Hæmoglobin (g/dL); median [IQR] | 16.1 [15.0;17.3] | 16.1 [14.6;17.3] † | 16.1 [14.9;17.3] † | 0.91 |
| ALT (U/L); n (%) | 0.88 | |||
| Normal ≤ 40 | 59 (92.2) | 70 (93.3) | 129 (92.8) | |
| Abnormal > 40 | 5 (7.8) | 5 (6.7) | 10 (7.2) | |
| Mild [40;100] | 3 (4.7) | 4 (5.3) | 7 (5.0) | |
| Moderate [100;200] | 2 (3.1) | 1 (1.3) | 3 (2.2) | |
| Haemoglobin (g/dL); n (%) | 0.10 | |||
| Normal > 13 | 62 (96.9) | 67 (90.5) † | 129 (93.5) † | |
| Anemia ≤ 13 | 2 (3.1) | 7 (9.5) † | 9 (6.5) † | |
| Mild [12;13] | 1 (1.6) | 6 (8.1) † | 7 (5.1) † | |
| Moderate [10;12] | - | 1 (1.4) † | 1 (0.72) † | |
| Very severe [0;9] | 1 (1.6) | - | 1 (0.72) † | |
| Platelet count (103/mm3); n (%) | 0.43 | |||
| Normal ≥125 | 59 (92.2) | 69 (97.2) ‡ | 128 (94.8) ‡ | |
| Thrombocytopenia <125 | 5 (7.8) | 2 (2.8) ‡ | 7 (5.2) ‡ | |
| Mild [100;125] | 3 (4.7) | 2 (2.8) ‡ | 5 (3.7) ‡ | |
| Moderate [50;100] | 1 (1.6) | - | 1 (0.7) ‡ | |
| Very severe [0;25] | 1 (1.6) | - | 1 (0.7) ‡ | |
| White cell count (103/mm3); n (%) | NA | |||
| Normal > 2.5 | 64 (100.0) | 73 (100.0) § | 137 (100.0) § | |
| Neutrophil count (103/mm3); n (%) | 0.22 | |||
| Normal > 1.5 | 62 (96.9) | 73 (100.0) § | 135 (98.5) § | |
| Neutropenia ≤ 1.5 | 2 (3.1) | - | 2 (1.5) § | |
| Mild [1.25;1.5] | 1 (1.6) | - | 1 (0.7) § | |
| Moderate [1.0;1.25] | 1 (1.6) | - | 1 (0.7) § |
| Characteristic | LPV/r (n = 64) | 3TC (n = 75) | Total (n = 139) | p Value a |
|---|---|---|---|---|
| At randomisation (D7) | ||||
| Social demographic characteristics | ||||
| Age (year); median [IQR] | 27.1 [24.1;30.7]] | 28.2 [24.2;32.6] | 27.4 [24.1;32.2] | 0.25 |
| Parity; median [IQR] | 3.0 [2.0;3.5] | 2.0 [1.0;4.0] | 3.0 [2.0;4.0] | 0.56 |
| Clinical and biological characteristics | ||||
| BMI; median [IQR] | 24.0 [21.4;27.5] | 23.4 [21.4;27.7] | 23.8 [21.4;27.7] | 0.88 |
| CD4 cells count (cells/mm3); median [IQR] | 556.0 [407.0;784.0] † | 584 [433.0;760] ‡ | 558.0 [420.0;766.0] § | 0.91 |
| HIV viral load (Log copies/mL); median [IQR] | 2.2 [2.2;3.3] | 2.2 [2.2;3.3] ¶ | 2.2 [2.2;3.3] ¶ | 0.79 |
| HIV viral load control; n (%) | 0.33 | |||
| Controlled <1000 copies/mL | 40 (62.5) | 52 (70.3) ¶ | 92 (66.7) ¶ | |
| Uncontrolled ≥1000 copies/mL | 24 (37.5) | 22 (29.7) ¶ | 46 (33.3) ¶ | |
| WHO HIV staging; n (%) | 1.00 | |||
| Stage 1 | 60 (93.8) | 71 (94.7) | 131 (94.2) | |
| Stage 2 | 4 (6.2) | 4 (5.3) | 8 (5.8) | |
| Antiretroviral prophylaxis | ||||
| ARV prophylaxis during pregnancy; n (%) | 0.47 | |||
| ZDV | 45 (70.3) | 51 (68.0) | 96 (69.1) | |
| ZDV + 3TC | 11 (17.2) | 18 (24.0) | 29 (20.9) | |
| No ARV prophylaxis | 8 (12.5) | 6 (8.0) | 14 (10.1) | |
| Duration of ARV prophylaxis taken during pregnancy (week); median [IQR] | 8.0 [5.5;12.5] ⱡ | 8.0 [4.0;14.0] ¤ | 8.0 [5.0;13.0] £ | 0.63 |
| During the trial | ||||
| HIV viral load at W38 (Log copies/mL); median [IQR] | 3.7 [2.2;4.8] | 3.4 [2.2;4.5] | 3.5 [2.2;4.7] | 0.23 |
| HIV viral load control at W38; n (%) | 0.18 | |||
| Controlled < 1000 copies/mL | 21 (32.8) | 33 (44.0) | 54 (38.9) | |
| Uncontrolled ≥ 1000 copies/mL | 43 (67.2) | 42 (56.0) | 85 (61.2) | |
| Duration of breastfeeding (week); median [IQR] | 45.6 [38.8;48.8] | 46.0 [35.7;48.6] | 45.9 [37.6;48.1] | 0.95 |
| Site | PrEP | n | MCN Median [IQR] | n | Children with MDD, n (%) |
|---|---|---|---|---|---|
| Burkina Faso | All | 46 | 922 [700;1112] | 44 | 43 (97.7) |
| LPV/r | 23 | 947 [757;1253] | 23 | 22 (95.7) | |
| 3TC | 23 | 833 [671;1112] | 21 | 21 (100.0) | |
| South Africa | All | 48 | 473 [329;799] | 48 | 48 (100.0) |
| LPV/r | 20 | 624 [447;1191] | 20 | 20 (100.0) | |
| 3TC | 28 | 422 [273;715] | 28 | 28 (100.0) | |
| Uganda | All | 45 | 976 [814;1099] | 45 | 45 (100.0) |
| LPV/r | 21 | 975 [814;1083] | 21 | 21 (100.0) | |
| 3TC | 24 | 1002 [819;1100] | 24 | 24 (100.0) | |
| All | All | 139 | 846 [555;1082] | 137 | 136 (99.3) |
| LPV/r | 64 | 897 [614;1135] | 64 | 63 (98.4) | |
| 3TC | 75 | 803 [484;1056] | 73 | 73 (100.0) |
| Site | PrEP | n | Decrease of MCN Median [IQR] | p Value a | Children with Depletion, n (%) | p Value b |
|---|---|---|---|---|---|---|
| Burkina Faso | All | 46 | 30.2% [2.5;54.4] | 13 (28.3) | ||
| LPV/r | 23 | 39.3% [12.1;65.9] | 0.12 | 8 (34.8) | 0.33 | |
| 3TC | 23 | 17.3% [−7.7;49.8] | 5 (21.7) | |||
| South Africa | All | 48 | 63.3% [37.8;75.0] | 30 (62.5) | ||
| LPV/r | 20 | 73.2% [50.9;85.6] | 0.02 | 15 (75.0) | 0.13 | |
| 3TC | 28 | 52.8% [34.8;70.4] | 15 (53.5) | |||
| Uganda | All | 45 | 35.4% [4.8;52.9] | 15 (33.3) | ||
| LPV/r | 21 | 17.8% [0.4;43.1] | 0.08 | 4 (19.0) | 0.06 | |
| 3TC | 24 | 49.2% [21.1;59.3] | 11 (45.8) | |||
| All | All | 139 | 41.7% [12.1;64.4] | 58 (41.7) | ||
| LPV/r | 64 | 40.1% [14.6;72.7] | 0.35 | 27 (42.2) | 0.97 | |
| 3TC | 75 | 42.2% [6.9;60.2] | 31 (41.3) |
| Group | PrEP | n | W6 MCN Median [IQR] | p Value a | W26 MCN Median [IQR] | p Value a |
|---|---|---|---|---|---|---|
| Control | No | 38 | 347 [258;407] | 194 [167;248] | ||
| PrEP | Yes | 47 | 458 [310;796] | <0.01 | 138 [122;148] | <0.01 |
| LPV/r | 19 | 682 [364;1220] | <0.01 | 132 [123;143] | <0.01 | |
| 3TC | 28 | 422 [273;691] | 0.07 | 141 [122;148] | <0.01 |
| Group | PrEP | n | Decrease of MCN Median [IQR] | p Value a | Children with mtDNA Depletion, n (%) |
|---|---|---|---|---|---|
| Control | No | 38 | 47.7% [10.4;55.9] | 16 (42.0) | |
| PrEP | Yes | 47 | 71.3% [59.0;88.1] | <0.01 | 39 (83.0) |
| LPV/r | 19 | 79.4% [61.8;88.2] | <0.01 | 17 (89.5) | |
| 3TC | 28 | 70.5% [55.3;78.9] | <0.01 | 22 (78.6) |
| Independent Variable | Crude Estimates | Adjusted Estimates | ||||
|---|---|---|---|---|---|---|
| PR | 95% CI (LB-UB) | p Value | PR | 95% CI (LB-UB) | p Value | |
| Gender | ||||||
| Girl | 1.0 | 1.00 | ||||
| Boy | 1.42 | 1.03–1.97 | 0.03 | 1.39 | 1.05–1.84 | 0.02 |
| PrEP * | ||||||
| No | 1.0 | 1.00 | ||||
| Yes | 1.97 | 1.33–2.92 | <0.01 | 1.63 | 1.08–2.45 | 0.02 |
| Mother age (per 5 years) | 1.05 | 0.92–1.19 | 0.47 | |||
| Parity | 0.95 | 0.81–1.12 | 0.56 | |||
| Duration of breastfeeding (weeks) | ||||||
| <39 | 1.00 | |||||
| ≥39 | 0.85 | 0.63–1.15 | 0.30 | |||
| Gestational age (weeks) | 1.05 | 0.98–1.13 | 0.17 | 1.08 | 0.98–1.19 | 0.11 |
| Weight of the infant at W6 (per 500 g) | 0.98 | 0.91–1.07 | 0.69 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monnin, A.; Nagot, N.; Periès, M.; Vallo, R.; Meda, N.; Singata-Madliki, M.; Tumwine, J.K.; Kankasa, C.; Ngandu, N.; Goga, A.; et al. Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1. J. Clin. Med. 2020, 9, 2972. https://doi.org/10.3390/jcm9092972
Monnin A, Nagot N, Periès M, Vallo R, Meda N, Singata-Madliki M, Tumwine JK, Kankasa C, Ngandu N, Goga A, et al. Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1. Journal of Clinical Medicine. 2020; 9(9):2972. https://doi.org/10.3390/jcm9092972
Chicago/Turabian StyleMonnin, Audrey, Nicolas Nagot, Marianne Periès, Roselyne Vallo, Nicolas Meda, Mandisa Singata-Madliki, James K. Tumwine, Chipepo Kankasa, Nobubelo Ngandu, Ameena Goga, and et al. 2020. "Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1" Journal of Clinical Medicine 9, no. 9: 2972. https://doi.org/10.3390/jcm9092972
APA StyleMonnin, A., Nagot, N., Periès, M., Vallo, R., Meda, N., Singata-Madliki, M., Tumwine, J. K., Kankasa, C., Ngandu, N., Goga, A., Reynier, P., Tylleskär, T., Van de Perre, P., & Molès, J.-P. (2020). Mitochondrial DNA Parameters in Blood of Infants Receiving Lopinavir/Ritonavir or Lamivudine Prophylaxis to Prevent Breastfeeding Transmission of HIV-1. Journal of Clinical Medicine, 9(9), 2972. https://doi.org/10.3390/jcm9092972