Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
- Presence of at least two of the following: temperature > 38 °C or <36.5 °C or temperature instability, tachycardia or bradycardia, apnea, prolonged capillary refill, metabolic acidosis, hypoglycaemia, other signs of bloodstream infections such as lethargy;
- Recognized pathogen cultured from one or more blood cultures or CoNS isolated from at least one blood culture or intravascular line and one of the following: C-reactive protein > 2.0 mg/dL, immature/total neutrophil ratio (I/T ratio) > 0.2, leukocytes < 5000/μL, platelets < 10,000/μL.
2.2. Ethics Approval
2.3. Consent Form

3. Microbiological Cultures
3.1. Blood Samples
3.2. Stool Samples
3.3. Catheter Tips
4. Molecular Typing
5. PCR Amplification of Genes Coding for Virulence Factors
6. Adherence of CoNS to CaCo2 Cells
- (a)
- strong adherence (3): >80 bacterial cells per field
- (b)
- moderate adherence (2): 61–80 bacterial cells per field
- (c)
- weak adherence (1): 41–60 bacterial cells per field
- (d)
- no adherence (0): <40 bacterial cells per field
7. Phenotypic Characteristic of Slime Production Ability on Congo Red Agar (CRA)
8. Statistical Analyses
9. Results
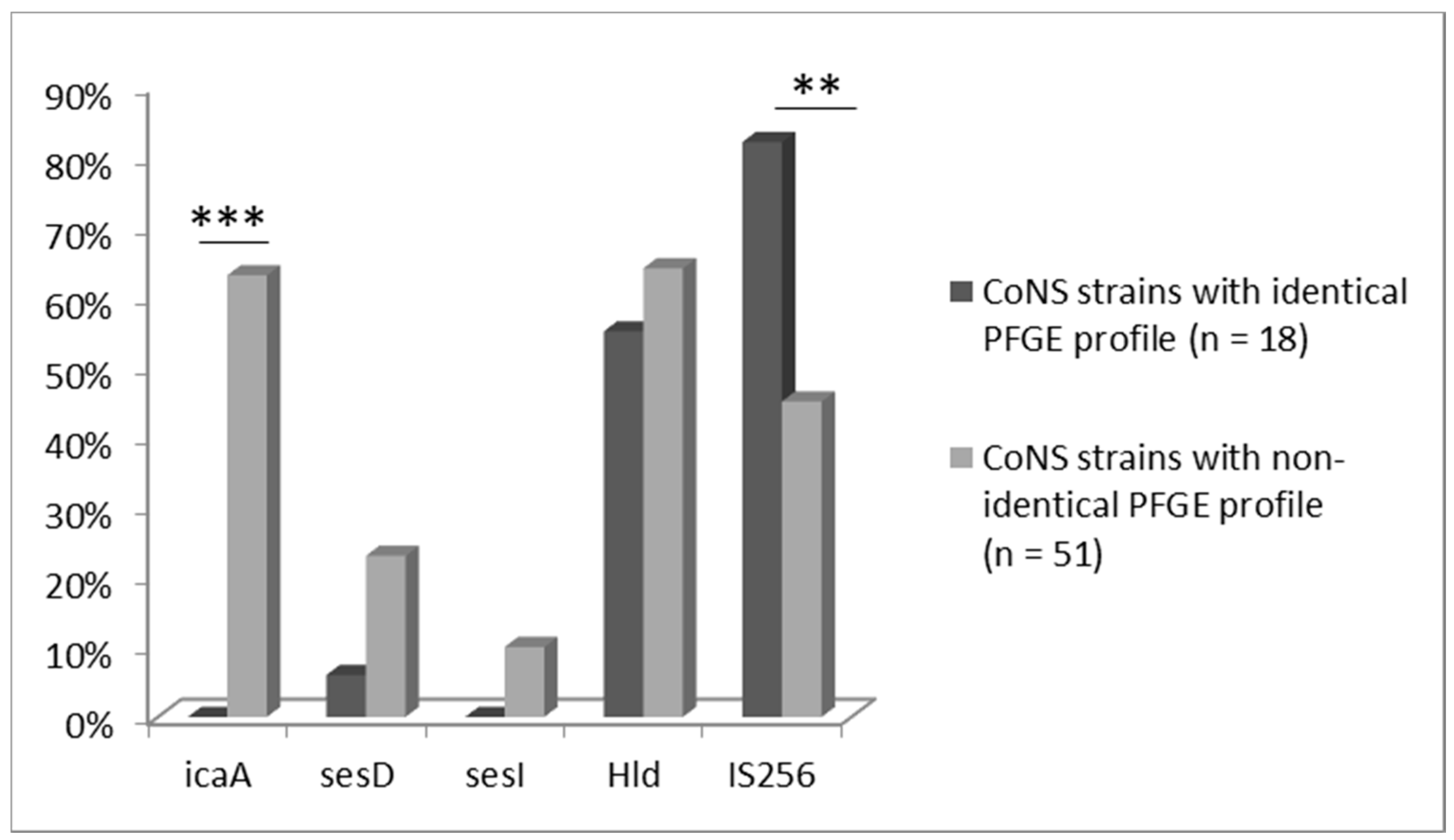
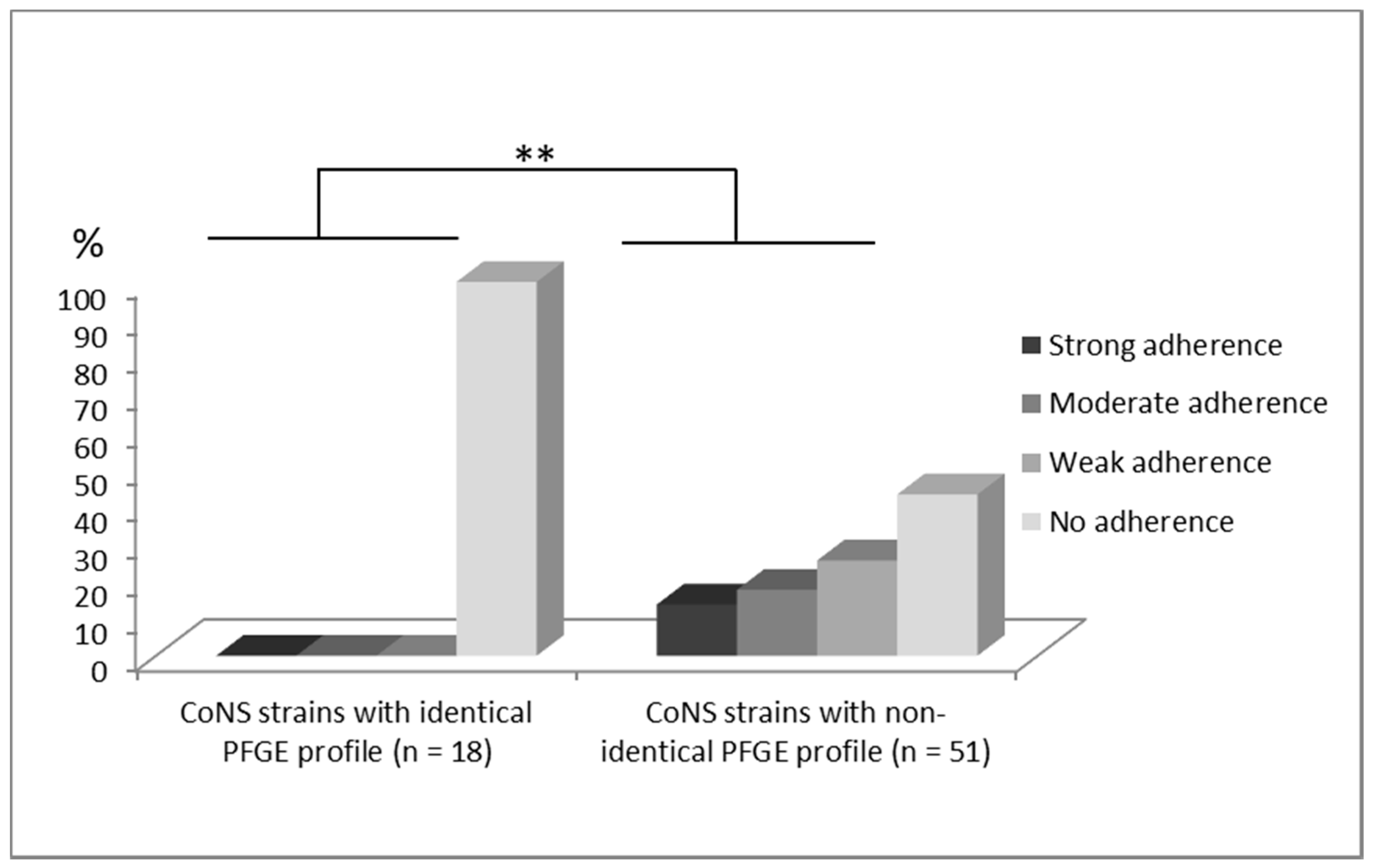
10. Discussion
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shah, B.A.; Padbury, J.F. Neonatal sepsis: An old problem with new insights. Virulence 2014, 5, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Cortese, F.; Scicchitano, P.; Gesualdo, M.; Filaninno, A.; De Giorgi, E.; Schettini, F.; Laforgia, N.; Ciccone, M.M. Early and late infections in newborns: Where do we stand? A review. Pediatr. Neonatol. 2016, 57, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Wójkowska-Mach, J.; Chmielarczyk, A.; Strus, M.; Lauterbach, R.; Heczko, P. Neonate bloodstream infections in organization for economic cooperation and development countries: An update on epidemiology and prevention. J. Clin. Med. 2019, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Cantey, J.B.; Anderson, K.R.; Kalagiri, R.R.; Mallett, L.H. Morbidity and mortality of coagulase-negative staphylococcal sepsis in very-low-birth-weight infants. World J. Pediatr. 2018, 14, 269–273. [Google Scholar] [CrossRef]
- Bizzarro, M.J.; Shabanova, V.; Baltimore, R.S.; Dembry, L.M.; Ehrenkranz, R.A.; Gallagher, P.G. Neonatal sepsis 2004–2013: The rise and fall of coagulase-negative staphylococci. J. Pediatr. 2015, 166, 1193–1199. [Google Scholar] [CrossRef]
- Wójkowska-Mach, J.; Gulczyńska, E.; Nowiczewski, M.; Borszewska-Kornacka, M.; Domańska, J.; Merritt, T.A.; Helwich, E.; Kordek, A.; Pawlik, D.; Gadzinowski, J.; et al. Late-onset bloodstream infections of very-low-birth-weight infants: Data from the Polish Neonatology Surveillance Network in 2009–2011. BMC Infect. Dis. 2014, 14, 339. [Google Scholar] [CrossRef]
- Marchant, E.A.; Boyce, G.K.; Sadarangani, M.; Lavoie, P.M. Neonatal sepsis due to coagulase-negative staphylococci. Clin. Dev. Immunol. 2013, 2013, 586076. [Google Scholar] [CrossRef]
- Bjarnsholt, T. The role of bacterial biofilms in chronic infections. APMIS Suppl. 2013, 136, 1–51. [Google Scholar] [CrossRef]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Adv. Exp. Med. Biol. 1999, 473, 11–30. [Google Scholar]
- Deitch, E.A. Gut-origin sepsis: Evolution of a concept. Surgeon 2012, 10, 350–356. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability: A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Yajima, M.; Nakayama, M.; Hatano, S.; Yamazaki, K.; Aoyama, Y.; Yajima, T.; Kuwata, T. Bacterial translocation in neonatal rats: The relation between intestinal flora, translocated bacteria, and influence of milk. J. Pediatr. Gastroenterol. Nutr. 2001, 33, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Ciftci, I.; Ozdemir, M.; Aktan, M.; Aslan, K. Bacterial translocation and intestinal injury in experimental necrotizing enterocolitis model. Bratisl. Lek. Listy 2012, 113, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Wujtewicz, M.A.; Śledzińska, A.; Owczuk, R.; Wujtewicz, M. Escherichia coli bacteraemias in intensive care unit patients. Anaesthesiol. Intensive Ther. 2016, 48, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Soeorg, H.; Huik, K.; Parm, Ü.; Ilmoja, M.L.; Metsvaht, T.; Lutsar, I. Molecular epidemiology of staphylococcus epidermidis in neonatal intensive care units. APMIS 2017, 125, 63–73. [Google Scholar] [CrossRef]
- Luo, C.C.; Shih, H.H.; Chiu, C.H.; Lin, J.N. Translocation of coagulase-negative bacterial staphylococci in rats following intestinal ischemia-reperfusion injury. Biol. Neonate 2004, 85, 151–154. [Google Scholar] [CrossRef]
- Soeorg, H.; Huik, K.; Parm, U.; Ilmoja, M.L.; Metelskaja, N.; Metsvaht, T.; Lutsar, I. Genetic relatedness of coagulase-negative staphylococci from gastrointestinal tract and blood of preterm neonates with late-onset sepsis. Pediatr. Infect. Dis. J. 2013, 32, 389–393. [Google Scholar] [CrossRef]
- Gastmeier, P.; Geffers, C.; Schwab, F.; Fitzner, J.; Obladen, M.; Rüden, H. Development of a surveillance system for nosocomial infections: The component for neonatal intensive care units in Germany. J. Hosp. Infect. 2004, 57, 126–131. [Google Scholar] [CrossRef]
- Guembe, M.; Martín-Rabadán, P.; Cruces, R.; Pérez Granda, M.J.; Bouza, E. Slicing silicone neonatal vascular catheter tips improves colonization detection by the roll-plate technique. Clin. Microbiol. Infect. 2017, 23, 410.e1–410.e3. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Villari, P.; Sarnataro, C.; Iacuzio, L. Molecular epidemiology of staphylococcus epidermidis in a neonatal intensive care unit over a three-year period. J. Clin. Microbiol. 2000, 38, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Chmielarczyk, A.; Pomorska-Wesołowska, M.; Szczypta, A.; Romaniszyn, D.; Pobiega, M.; Wójkowska-Mach, J. Molecular analysis of methicillin-resistant staphylococcus aureus strains isolated from different types of infections from patients hospitalized in 12 regional, non-teaching hospitals in southern Poland. J. Hosp. Infect. 2017, 95, 259–267. [Google Scholar] [CrossRef]
- Söderquist, B.; Andersson, M.; Nilsson, M.; Nilsdotter-Augustinsson, A.; Persson, L.; Friberg, O.; Jacobsson, S. Staphylococcus epidermidis surface protein I (SesI): A marker of the invasive capacity of S. epidermidis? J. Med. Microbiol. 2009, 58, 1395–1397. [Google Scholar] [CrossRef]
- Soumya, K.R.; Sheela, S.; Jyothis, M.; Radhakrishnan, E.K. Studies on coexistence of mec gene, IS256 and novel sasX gene among human clinical coagulase-negative staphylococci. 3 Biotech 2016, 6, 233. [Google Scholar] [CrossRef]
- Marconi, C.; Cunha, M.L.R.S.; Araujo, J.R.J.P.; Rugolo, L.M.S.S. Standarization of the PCR technique for the detection of delta toxin in Staphylococcus spp. J. Venom. Anim. Toxins Incl. Trop. Dis. 2005, 11, 117–128. [Google Scholar] [CrossRef]
- Thomas, J.C.; Zhang, L.; Robinson, D.A. Differing lifestyles of staphylococcus epidermidis as revealed through Bayesian clustering of multilocus sequence types. Infect. Genet. Evol. 2014, 22, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Eftekhar, F.; Dadaei, T. Biofilm formation and detection of icaAB genes in clinical isolates of methicillin resistant Staphylococcus aureus. Iran. J. Basic Med. Sci. 2010, 14, 132–136. [Google Scholar]
- Golińska, E.; van der Linden, M.; Więcek, G.; Mikołajczyk, D.; Machul, A.; Samet, A.; Piórkowska, A.; Dorycka, M.; Heczko, P.B.; Strus, M. Virulence factors of streptococcus pyogenes strains from women in peri-labor with invasive infections. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 747–754. [Google Scholar] [CrossRef]
- Arciolaa, C.R.; Campocciaa, D.; Gamberinia, S.; Cervellatia, M.; Donatia, E.; Montanaro, L. Detection of slime production by means of an optimised Congo red agar plate test based on a colourimetric scale inStaphylococcus epidermidis clinical isolates genotyped for ica locus. Biomaterials 2002, 23, 4233–4239. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Stewart, C.J.; Embleton, N.D.; Marrs, E.C.L.; Smith, D.P.; Fofanova, T.; Nelson, A.; Skeath, T.; Perry, J.D.; Petrosino, J.F.; Berrington, J.E.; et al. Longitudinal development of the gut microbiome and metabolome in preterm neonates with late onset sepsis and healthy controls. Microbiome 2017, 5, 75. [Google Scholar] [CrossRef]
- Dong, Y.; Speer, C.P.; Glaser, K. Beyond sepsis: Staphylococcus epidermidis is an underestimated but significant contributor to neonatal morbidity. Virulence 2018, 9, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Donnell, S.C.; Taylor, N.; van Saene, H.K.F.; Magnall, V.L.; Pierro, A.; Lloyd, D.A. Infection rates in surgical neonates and infants receiving parenteral nutrition: A five-year prospective study. J. Hosp. Infect. 2002, 4, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Milani, C.; Duranti, S.; Lugli, G.A.; Bernasconi, S.; Margolles, A.; Di Pierro, F.; van Sinderen, D.; Ventura, M. The infant gut microbiome as a microbial organ influencing host well-being. Ital. J. Pediatr. 2020, 46, 16. [Google Scholar] [CrossRef] [PubMed]
- Soeorg, H.; Metsvaht, H.K.; Keränen, E.E.; Eelmäe, I.; Merila, M.; Ilmoja, M.L.; Metsvaht, T.; Lutsar, I. Genetic relatedness of staphylococcus haemolyticus in gut and skin of preterm neonates and breast milk of their mothers. Pediatr. Infect. Dis. J. 2019, 38, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Jin, Y.; Duan, J.; Hao, Z.; Wang, S.; Guo, Y.; Lv, J.; Hu, L.; Wang, L.; Yu, F. SesI may be associated with the invasiveness of staphylococcus epidermidis. Front. Microbiol. 2018, 4, 2474. [Google Scholar] [CrossRef]
- Da, F.; Joo, H.S.; Cheung, G.Y.C.; Villaruz, A.E.; Rohde, H.; Luo, X.; Otto, M. Phenol-soluble modulin toxins of staphylococcus haemolyticus. Front. Cell. Infect. Microbiol. 2017, 24, 206. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis: A major player in bacterial sepsis? Future Microbiol. 2017, 12, 1031–1033. [Google Scholar] [CrossRef]
- Kleinert, F.; Kallies, R.; Hort, M.; Zweynert, A.; Szekat, C.; Nagel, M.; Bierbaum, G. Influence of IS256 on genome variability and formation of small-colony variants in staphylococcus aureus. Antimicrob. Agent Chemother. 2017, 61, 607–624. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Koskela, A.; Nilsdotter-Augustinsson, A.; Persson, L.; Soderquist, B. Prevalence of the ica operon and insertion sequence IS256 among Staphylococcus epidermidis prosthetic joint infection isolates. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 655–660. [Google Scholar] [CrossRef]
- Gu, J.; Li, H.; Li, M.; Vuong, C.; Otto, M.; Wen, Y.; Gao, Q. Bacterial insertion sequence IS256 as a potential molecular marker to discriminate invasive strains from commensal strains of staphylococcus epidermidis. J. Hosp. Infect. 2005, 61, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhu, Y.; Song, Y.; Li, T.; Luo, T.; Sun, G.; Yang, C.; Cao, C.; Lu, Y.; Li, M. Molecular analysis of staphylococcus epidermidis strains isolated from community and hospital environments in China. PLoS ONE 2013, 8, e62742. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, M.; Stalin, M.; Indhumathy, K.; Padma, K. Role of insertion sequence element ISi256 as a virulence marker and its association with biofilm formation among methicillin-resistant staphylococcus epidermidis from hospital and community settings in Chennai, South India. Brief Commun. 2018, 36, 124. [Google Scholar]
- Massonet, C.; Pintens, V.; Merckx, R.; Anné, J.; Lammertyn, E.; Van Eldere, J. Effect of iron on the expression of sirR and sitABC in biofilm-associated staphylococcus epidermidis. BMC Microbiol. 2006, 19, 103. [Google Scholar]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef]
- Strus, M.; Helwich, E.; Lauterbach, R.; Rzepecka-Węglarz, B.; Nowicka, K.; Wilińska, M.; Szczapa, J.; Rudnicka, M.; Sławska, H.; Szczepański, M.; et al. Effects of oral probiotic supplementation on gut Lactobacillus and Bifidobacterium populations and the clinical status of low-birth-weight preterm neonates: A multicenter randomized, double-blind, placebo-controlled trial. Infect. Drug Resist. 2018, 21, 1557–1571. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Product Size (bp) | PCR Conditions | References |
|---|---|---|---|---|---|
| icaA | AAA CTT GGT GCG GTT ACA GG | TCT GGG CCT GAC CAT GTT G | 750 | 1 min 94 °C 30 s 57 °C 1 min 72 °C 39 cycles | [26] |
| IS256 | TGAAAAGCGAAGAGATTCAAAGC | ATGTAGGTCCATAAGAACGGC | 1102 | 1 min 94 °C 1 min 59 °C 90 s 72 °C 34 cycles | [23] |
| sesD | TGCCAATCATCAAACCACTG | GTCACTGAACCGATTAACCCACTT | 635 | 1 min 94 °C 1 min 50 °C 1 min 72 °C 30 cycles | [25] |
| sesI | GCT GAT TAT GTA AAT GAC TCA AAT | AGC TTT TGT TGT TTG AGC TTC | 389 | 1 min 95 °C 1 min 49 °C 1 min 72 °C 34 cycles | [22] |
| Hld | ATG GCA GCA GAT ATC ATT TC | CGT GAG CTT GGG AGA GAC | 444 | 2 min 94 °C 1 min 45 °C 1 min 72 °C 29 cycles | [24] |
| Etiological Factors: | Number of Isolates (n = 511) from 468 Blood Samples | Number of Isolates (n = 148) from 119 Stool Samples | Number of Isolates (n = 7) from 8 Catheter Tips |
|---|---|---|---|
| Coagulase-negative staphylococci | 368 (72%) | 82 (55.4%) | 6 (85.7%) |
| S. epidermidis | 164 | 47 | 4 |
| S. haemolyticus | 80 | 16 | 1 |
| S. capitis | 58 | 2 | 0 |
| S. hominis | 30 | 0 | 0 |
| S. lugdunensis | 2 | 0 | 0 |
| S. warneri | 2 | 1 | 0 |
| S. xylosus | 1 | 0 | 0 |
| S. caprae | 1 | 0 | 0 |
| CNS | 30 | 16 | 1 |
| Coagulase-positive staphylococci | 15 (2.9%) | 4 (2.7%) | 1 (14.3%) |
| S. aureus | 15 | 4 | 1 |
| Gram negative rods | 79 (15.5%) | 32 (21.6%) | 0 |
| Klebsiella spp. | 41 | 4 | 0 |
| E. coli | 16 | 9 | 0 |
| S. marcescens | 9 | 0 | 0 |
| E. cloace | 7 | 0 | 0 |
| P. mirabilis | 3 | 0 | 0 |
| A. baumannii | 1 | 0 | 0 |
| S. linguefaciens | 1 | 0 | 0 |
| P. aeruginosa | 1 | 0 | 0 |
| other | 0 | 19 | 0 |
| Other (Micrococcus spp., Streptococcus spp., Candida spp.) | 49 (9.6%) | 30 (20.3%) | 0 |
| Sample Pairs (Blood + Stool) Taken from the Same Newborn (n = 69) | Sample Pairs (Blood + Catheter Tips) Taken from the Same Newborn (n = 6) | |||||
|---|---|---|---|---|---|---|
| PFGE | PFGE | |||||
| Total Number | Identical Profile | Not Identical Profile | Total Number | Identical Profile | Not Identical Profile | |
| Coagulaso negative staphylococci | 69 | 18 | 51 | 6 | 0 | 6 |
| S. epidermidis | 32 | 4 | 28 | 4 | 0 | 4 |
| S. haemolyticus | 22 | 12 | 10 | 1 | 0 | 1 |
| S. capitis | 10 | 2 | 8 | 0 | 0 | 0 |
| S. hominis | 2 | 0 | 2 | 0 | 0 | 0 |
| S. warneri | 1 | 0 | 1 | 0 | 0 | 0 |
| CNS | 2 | 0 | 2 | 1 | 0 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golińska, E.; Strus, M.; Tomusiak-Plebanek, A.; Więcek, G.; Kozień, Ł.; Lauterbach, R.; Pawlik, D.; Rzepecka-Węglarz, B.; Kędzierska, J.; Dorycka, M.; et al. Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates. J. Clin. Med. 2020, 9, 2517. https://doi.org/10.3390/jcm9082517
Golińska E, Strus M, Tomusiak-Plebanek A, Więcek G, Kozień Ł, Lauterbach R, Pawlik D, Rzepecka-Węglarz B, Kędzierska J, Dorycka M, et al. Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates. Journal of Clinical Medicine. 2020; 9(8):2517. https://doi.org/10.3390/jcm9082517
Chicago/Turabian StyleGolińska, Edyta, Magdalena Strus, Anna Tomusiak-Plebanek, Grażyna Więcek, Łucja Kozień, Ryszard Lauterbach, Dorota Pawlik, Beata Rzepecka-Węglarz, Jolanta Kędzierska, Małgorzata Dorycka, and et al. 2020. "Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates" Journal of Clinical Medicine 9, no. 8: 2517. https://doi.org/10.3390/jcm9082517
APA StyleGolińska, E., Strus, M., Tomusiak-Plebanek, A., Więcek, G., Kozień, Ł., Lauterbach, R., Pawlik, D., Rzepecka-Węglarz, B., Kędzierska, J., Dorycka, M., & Heczko, P. B. (2020). Coagulase-Negative Staphylococci Contained in Gut Microbiota as a Primary Source of Sepsis in Low- and Very Low Birth Weight Neonates. Journal of Clinical Medicine, 9(8), 2517. https://doi.org/10.3390/jcm9082517