1. Introduction
Urinary bladder cancer causes approximately 550,000 new cases annually in the world, that are the 3.0% of all new cancer diagnoses, and 2.1% of all cancer deaths [
1]. Among the geographical area, Europe and North America record a three-fold higher rates of incidence, probably due to the risk factors’ exposure as cigarette smoking, chemical carcinogens, arsenic drinking water, obesity, and chronic urinary infections caused by
Schistosoma hematobium [
2]. Among all bladder cancer cases, transitional cell carcinoma (TCC) accounts for 95% of them. TCC is a malignant cancer that originates from the bladder transitional epithelial cells, and often establishes the metastatic disease. It should be note that although from 2000 the TCC incidence-based mortality has started to decrease, these data exclude the mortality cases due to TCC that causes distant metastasis, which has continued to increase since 2012 [
3].
ATRA is one of the most biologically-active forms of vitamin A in cells. It influences cellular growth and differentiation in several ways, but mostly by transcriptionally regulating gene expression by binding to nuclear retinoic acid receptors (RARs) and retinoid X receptors (RXRs) [
4]. From the U.S. Food and Drug Administration approval in 1995, the ATRA oral administration, at pharmacological doses (45 mg/m
2/day for adults, in comparison to 900 and 700 μg/day of average requirement for adult men and women, respectively), has revolutionized the treatment of APL, increasing the complete remission rate up to 90–95% [
5]. Despite the high complete remission rates, the relative contribution of maintenance therapy (i.e., mostly 45 mg/m
2/day oral ATRA treatment for 15 days every 90 days for 2 years) is controversial, mostly for high-risk patients (i.e., white cell count > 10 × 10
9/L) [
6]. However, beyond the close connection with APL, pharmacological ATRA treatment may have different systemic effects. Indeed, it is well known that in the treatment of solid cancers the ATRA action is questionable. Although, ATRA can inhibit cancer cell proliferation at different levels, from membrane to gene expression, in parallel the cancer cells have evolved several escape routes to bypass these ATRA actions, developing ATRA-resistance [
4]. In some cases, molecular pathways deregulations reverse the ATRA action from the growth inhibitory/differentiating to the anti-apoptotic/proliferative action. Indeed, for example in the classical pathway, ATRA in the cytosol is tied to CRABP-II, which delivers it to RARs. However, it was shown that some solid cancers activate a non-genomic pathway due to less CRABP-II concentration in comparison to another retinol-binding protein, FABP5. In these cases, FABP5 delivered ATRA to the nuclear PPARβ/δ stimulating cellular proliferation [
4,
7]. Although few papers showed that the maintenance treatment of APL with ATRA is not associated with higher incidence of secondary cancers, it is worth noting that long-term follow-up studies that analyze the APL maintenance therapy adverse effects are actually limited and/or examined a very limited statistical population [
8,
9].
T24 is a TCC cell line, grade 3, already studied in the literature in correlation to pharmacological ATRA treatment, even if with conflicting results. Indeed, already in 1999 Waliszewski and co-workers failed to found viability inhibition in T24 cell line after 10 µM ATRA and 13-
cis retinoic acid (13-
cis RA) treatments [
10]. In the same paper the authors found undetectable or low levels of RARβ, cellular retinol-binding protein type-I (CRBP-I), CRABP-I, and CRABP-II in the constitutive and stimulated (48 h 10 µM ATRA stimulation) T24 cells [
10]. Zou and co-workers in the paper of 2001, found the same result [
11]. Conversely, in the following paper by Zou and co-workers, a different result was found: Indeed increasing the ATRA concentration (from 1 to 10 µM) increased the growth inhibitory effect in T24 cell line, even if the apoptosis induction was similar to control [
12].
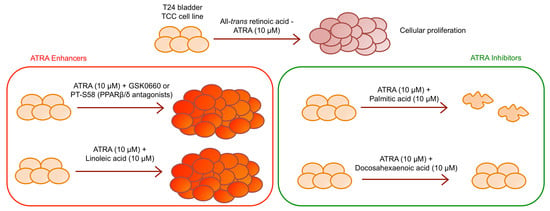
Considering the discrepancies present in the literature, the present paper evaluated the hypothesized ATRA resistance of T24 cell line, finding that ATRA activates survival mechanism inducing T24 cell proliferation. First investigations about molecular mechanism were carried out analyzing the FABP5-PPARβ/δ pathway, and even if over-expression of FABP5 was found concomitantly with CRABPII down-expression, the direct inhibition of PPARβ/δ did not abolish T24 cellular proliferation, but rather potentiated it. Moreover, considering the paper by Levi [
13] and colleagues that demonstrated the action of LCFAs to displacing ATRA from FABP5, where saturated LCFAs inhibit FABP5-PPARβ/δ pathway, while conversely, unsaturated LCFAs activate it, the actions of the saturated PA, unsaturated omega-6 LA, and unsaturated omega-3 DHA were evaluated to counteract ATRA-related proliferation. In line with Levi and co-workers we found that ATRA-PA co-treatment induces cellular growth inhibition and ATRA-LA co-treatment induces cellular growth enhancement [
13]. However, conversely to the generalization stated from Levi and co-workers that unsaturated LCFAs induce cell proliferation [
13], we found that the unsaturated omega-3 DHA reverse the ATRA-induced cellular proliferation of T24 bladder cancer cell line.
2. Materials and Methods
2.1. Cells and Treatments
T24 TCC line was purchased from ATCC (Manassas, VA, USA) and cultured in McCoy’s 5A medium supplemented with 10% FBS or FBS charcoal-stripped (FBS-CS), 1.5 mM L-glutamine, and 10 mL/L penicillin/streptomycin, at 37 °C under 5% CO2 (at most 5 passages—three days in culture each—from the original ATCC stock). Cells were daily treated with: ATRA 1, 10, and 20 µM; synthetic retinoid fenretinide or 4-HPR (4-hydroxy(phenyl)retinamide) 10 µM; GSK0660 selective PPARδ antagonist 2.5, 5, 10, 20, 30, and 50 µM; PT-S58 (BOC Sciences, Shirley, NY, USA) full PPARβ/δ antagonist 5, 10, 15, and 20 µM; PA, LA, and DHA 10 µM. Controls were done with the corresponding vehicles (DMSO for ATRA, and ethanol for LCFA) in the same percentage present in the treatments. Treatment lapses are reported in the figures for each analysis, anyway with a maximum of 72 h or 96 h. All materials were purchased from Merck (Darmstadt, Germany) except when differently indicated.
2.2. Cellular Viability and Proliferation Tests
MTT proliferation assay was used to analyze cellular viability. Briefly, 1500 cells were plated in a 96-well plate in replicates (n = 12) and indicated treatment. Cells were incubated with MTT reagent (5 mg/mL) until the formation of formazan crystal (3.5 h). Crystals were dissolved in 1 M HCl in isopropanol, and absorbance at 570 nm was measured using a microplate reader (Infinite 2000, Tecan, Männedorf, Switzerland).
Colony formation assay was used to analyze cellular proliferation. A bottom layer of 0.8% agarose in cell media was cast, in sterile hood, in a six-well plate and set at room temperature to solidify. Cells were suspended in 0.25% agarose in media, 2 mL from this mixture containing 4500 cells was added to each well, and after solidification, 1 mL media with vehicle or treatment was added in each well. Cells were cultured for 21 days. Media were replenished every 3 days. Colonies were visualized by staining with 0.005% crystal violet and counted under a light microscope. The images were acquired through ChemiDoc XRS + (BioRad, Hercules, CA, USA).
Absolute cell count was made through NovoCyte flow cytometer (ACEA Biosciences Inc., San Diego, CA, USA) with Novo Express Software (ACEA Biosciences Inc., San Diego, CA, USA) on T24 cells DMSO- and 10 µM ATRA-treated at 72 h.
2.3. Differential Proteomics Analysis
2.3.1. Protein Extraction and Solubilization
DMSO and ATRA treated cells were collected and centrifuged at 1500 rpm for 5 min. Cells were suspended in lysis buffer (7 M urea, 2 M tiourea, 4% w/v CHAPS, 40 mM Tris-HCl, 0.1 mM EDTA, 1 mM DTT, 50 mM NaF, and 0.25 mMNa3VO4). The mixture was centrifuged at 12,000× g at room temperature for 15 min and a small aliquot was used to determine the protein content by 2D-Quant Kit (GE Healthcare, Chicago, IL, USA).
2.3.2. 2D Gel Electrophoresis and Image Analysis
To remove lipids, 500 μg of protein were precipitated from a desired volume of each sample with a cold mix of tri-n-butyl phosphate/acetone/methanol (1:12:1). After incubation at 4 °C for 90 min, the precipitate was pelleted by centrifugation at 2800 ×g, for 20 min at 4 °C. The pellet was air-dried and then solubilized in the buffer containing 7 M urea, 2 M thiourea, 4% (w/v) CHAPS, 40 mM Tris-HCl. The sample was subsequently reduced (5 mM tributylphosphine, 1 h) and alkylated (7.7 Mm IAA, 1 h). To prevent over-alkylation, iodoacetamide (IAA) excess was neutralized by adding 10 mM DTE. Sample is included in the rehydration solution (7 M Urea, 2 M Thiourea, 4% CHAPS and 0.5% w/v pH 4–7 carrier ampholyte (Bio-Rad, Hercules, CA, USA)) and the sample is taken up into the 17 cm IPG strips pH 3–10 (Bio-Rad, Hercules, CA, USA) passively during rehydration overnight. IEF was performed using Bio-Rad Multiphore II and Dry Strip Kit (Bio-Rad-Protean IEF-Cell-System). Seventeen centimeters IPG strips (Bio-Rad, Hercules, CA, USA) pH 4–7 were passively rehydrated overnight with 500 μg of protein in 300 μL of solubilization solution (7 M Urea, 2 M Thiourea, 4% CHAPS and 0.5% w/v pH 4–7 carrier ampholyte (Bio-Rad, Hercules, CA, USA)). The total product time × voltage applied was 80,000 V h for each strip at 20 °C. Strips were equilibrated (30 min in 50 mM Tris-HCl pH 8.8, 6 M urea, 30% glycerol v/v, 1% SDS, bromophenol blue) and then placed on SDS-polyacrylamide gels, 18.5 cm × 20 cm, 13% acrylamide, and sealed with 0.5% agarose. SDS-PAGE was performed using the Bio-Rad Protean II XL Cell, large gel format, at constant current (40 mA per gel) at 7 °C until the bromophenol blue tracking dye was approximately 2–3 mm from the bottom of the gel. Protein spots were stained with colloidal Coomassie Brilliant Blue G-250. To ensure protein pattern reproducibility, two technical replicates for each biological replicate were done. The scanned gel images were transferred to the Progenesis SameSpots software package (Nonlinear Dynamics, Newcastle, UK), which allows spot detection, background subtraction, and protein spot OD intensity quantification (spot quantity definition). The gel image showing the highest number of spots and the best protein pattern was chosen as a reference template and the images were aligned onto it. All spots were prefiltered and manually checked before applying the statistical criteria (ANOVA p < 0.05 and fold ≥ 1.5).
2.3.3. In-Gel Digestion and LC-MS/MS Analysis
Gel bands were carefully excised from the gel and subjected to in-gel trypsin digestion according to Shevchenko and colleagues [
14]. Peptide extracts were analyzed by using a split-free nano-flow liquid chromatography system (EASY-nLC II, Proxeon, Odense, Denmark) coupled with a 3D-ion trap (model AmaZon ETD, Bruker Daltonik, Billerica, MA, USA) equipped with an online ESI nanosprayer (the spray capillary was a fused silica capillary, 0.090 mm OD, 0.020 mm ID) in the positive-ion mode. For all experiments, a sample volume of 15 μL was loaded by the autosampler onto a homemade 2-cm fused silica precolumn (100 μm I.D.; 375 μm O.D.; Reprosil C18-AQ, 5 μm, Dr. Maisch GmbH, Ammerbuch-Entringen, Germany). Sequential elution of peptides was accomplished by using a flow rate of 300 nL/min and a linear gradient from Solution A (100% water; 0.1% formic acid) to 50% of Solution B (100% acetonitrile; 0.1% formic acid) in 40 min over the precolumn on line with a homemade 15 cm resolving column (75 μm ID; 375 μm OD; Reprosil C18-AQ, 3 μm, Dr. Maisch GmbH, Ammerbuch-Entringen, Germany). The acquisition parameters for the mass spectrometer were as previously reported. Acquired MS/MS spectra were processed in DataAnalysis 4.0 and submitted to the Mascot search program (Matrix Science, London, UK). The following parameters were adopted for database searches: SwissProt database (release date 7 April 2017; 5,011,440 sequences); taxonomy = Homo sapiens; peptide and fragment mass tolerance = ±0.3 Da; missed cleavages = 1; fixed modifications = carbamidomethyl (C); variable modifications: oxidation (M) and significance threshold level (
p < 0.05) for Mascot scores (−10 Log (P)). In the case of hits with only one statistically significant unique peptide, even though high Mascot scores were obtained with significant values, a combination of automated database searches and manual interpretation of peptide fragmentation spectra was used to validate protein assignments.
For the analysis of functional relationships between the proteins that exhibited differences in expression String DB was used.
2.4. Quantitative Real-Time PCR
T24 RNA was extracted from 106 cells using the “high pure RNA isolation kit” (Roche, Basel, Switzerland) and quantified through Qubit 3.0 Fluorometer (Life Technologies, Carlsbad, CA, USA) and Qubit RNA BR Assay kit (Life Technologies, Carlsbad, CA, USA). cDNA was generated using the “transcriptor universal cDNA master” (Roche, Basel, Switzerland). Quantitative real-time PCR was carried out using the “LightCycler 480 SYBR Green I Master” (Roche, Basel, Switzerland) on LightCycler 480 instrument (Roche, Basel, Switzerland), with the following primer pairs (TIB MOLBIOL S.r.l., Genova, Italy):
CRABP-II:
Forward primer—ATGCCCAACTTCTCTGGCAA
Reverse primer—CGTCATGGTCAGGATCAGTT
FABP5:
Forward primer—AGCAGCTGGAAGGAAGATGG
Reverse primer—CTGATGCTGAACCAATGCAC
β-actin:
Forward primer—GCATGGAGTCCTGTGGCAT
Reverse primer—CTAGAAGCATTTGCGGTGG
For each treatment three technical replicates of each three biological replicates were performed (n = 9). As calibrator RNA of non-cancerous human urothelial cells (HUC) was used (ScienCell Research Laboratories, Carlsband, CA, USA). Relative expression was calculated by the E-Method, with β-actin as the reference.
2.5. Immunoblotting
Total cell proteins were extracted from T24 cell line using a buffer containing 250 mM TRIS-HCl pH 7.5, 750 mM NaCl, 25 mM EDTA, 10% glycerol, 1% IGEPAL CA-630, and 1% protease inhibitors. Non-cancerous HUC lysate was purchased from ScienCell Research Laboratories (Carlsband, CA, USA). Human bladder normal tissue lysate total protein was purchased from Abcam (Cambridge, UK). The protein concentration of the supernatants was determined with the BCA protein assay. 40 µg of proteins were resolved by electrophoresis on SDS-PAGE precast gels (4–20% mini protean TGX gels, BioRad, Hercules, CA, USA) and transferred onto nitrocellulose membrane (Amersham, GE healthcare, Chicago, IL, USA). Membranes were incubated with primary antibodies, followed by washes with Tween-PBS, and incubation with horse-radishperoxidase-conjugated antibodies. Protein expression was detected by exposure to westar supernova (Cyanagen, Bologna, Italy) and the images were collected through ChemiDoc XRS + (BioRad, Hercules, CA, USA). Antibody against CRABP-II II was purchased from Thermo Fischer scientific (Waltham, IL, USA), antibodies against, FABP5 and β-actin were purchased from Santa Cruz Biotechnology (Dallas, TX, USA), and they were used at 1:500 dilutions for CRABP-II and FABP5, and 1:1000 dilution for β-actin. Densitometry of various analyzed proteins and their respective loading controls from the same blot was performed using ImageJ Software (
https://imagej.nih.gov/ij/index.html).
2.6. Flow-Cytometer Analysis of Apoptosis
Apoptosis was determined with annexin V-FITC/propidium iodide (PI) apoptosis detection kit (Bender MedSystems, Wien, Austria). Following treatment, aliquots containing 5 × 105 cells in 195 µL buffer were stained with 5 µL FITC-conjugated Annexin V for 10 min and after washing and resuspension in 190 µL buffer, cells were stained with 10 µL PI solution. Samples were stored on ice until data acquisition. The analysis was performed by NovoCyte flow cytometer (ACEA Biosciences Inc., San Diego, CA, USA) with Novo Express Software (ACEA Biosciences Inc., San Diego, CA, USA).
2.7. Statistical Analysis
The mean and standard deviation (SD) of at least three biological replicates were calculated for all the analyzed data. Statistical analysis was performed with the XLSTAT 2018.5 (Addinsoft SARL, Paris, France) software using one-way ANOVA. Fisher’s least significant differences test was used to describe statistical differences between means at the p < 0.05 significance level.
4. Discussion
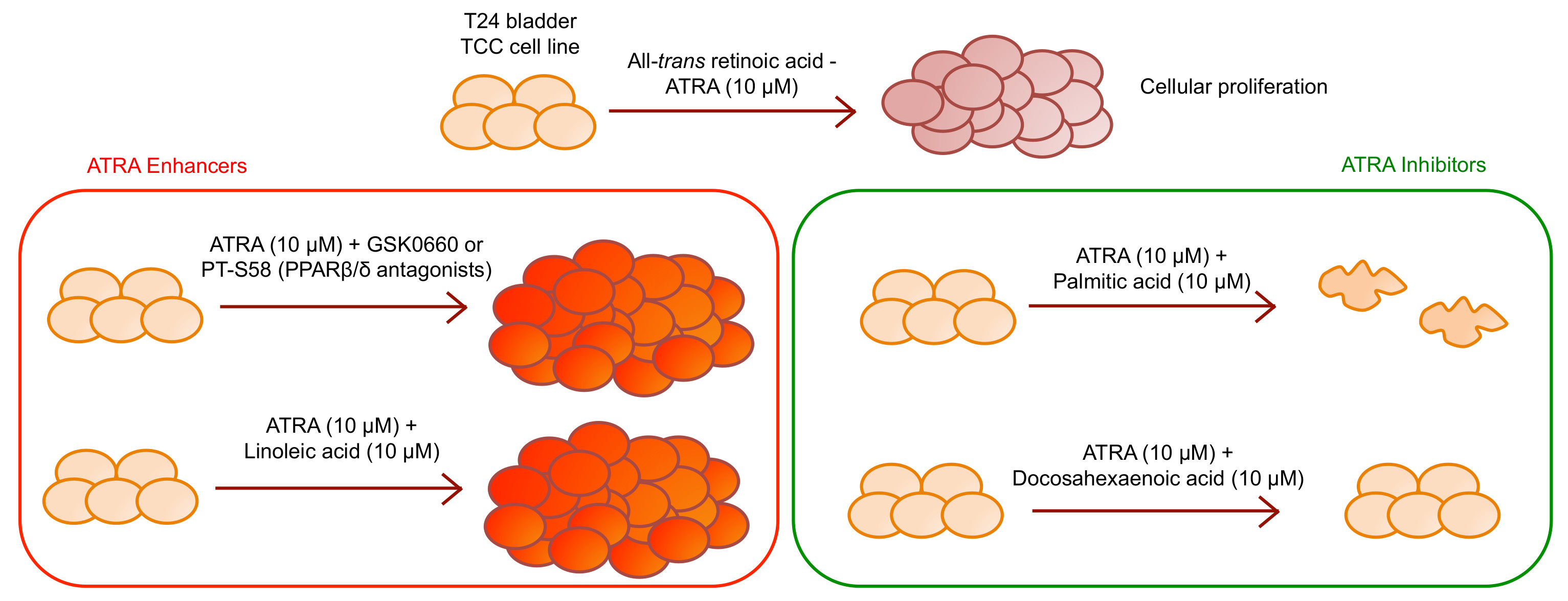
In the present paper we have shown, for the first time, that T24 cell line are not only resistant to rising ATRA concentrations, up to 20 µM, but also that ATRA induces a dose and time-dependent cellular proliferation (
Figure 1b).
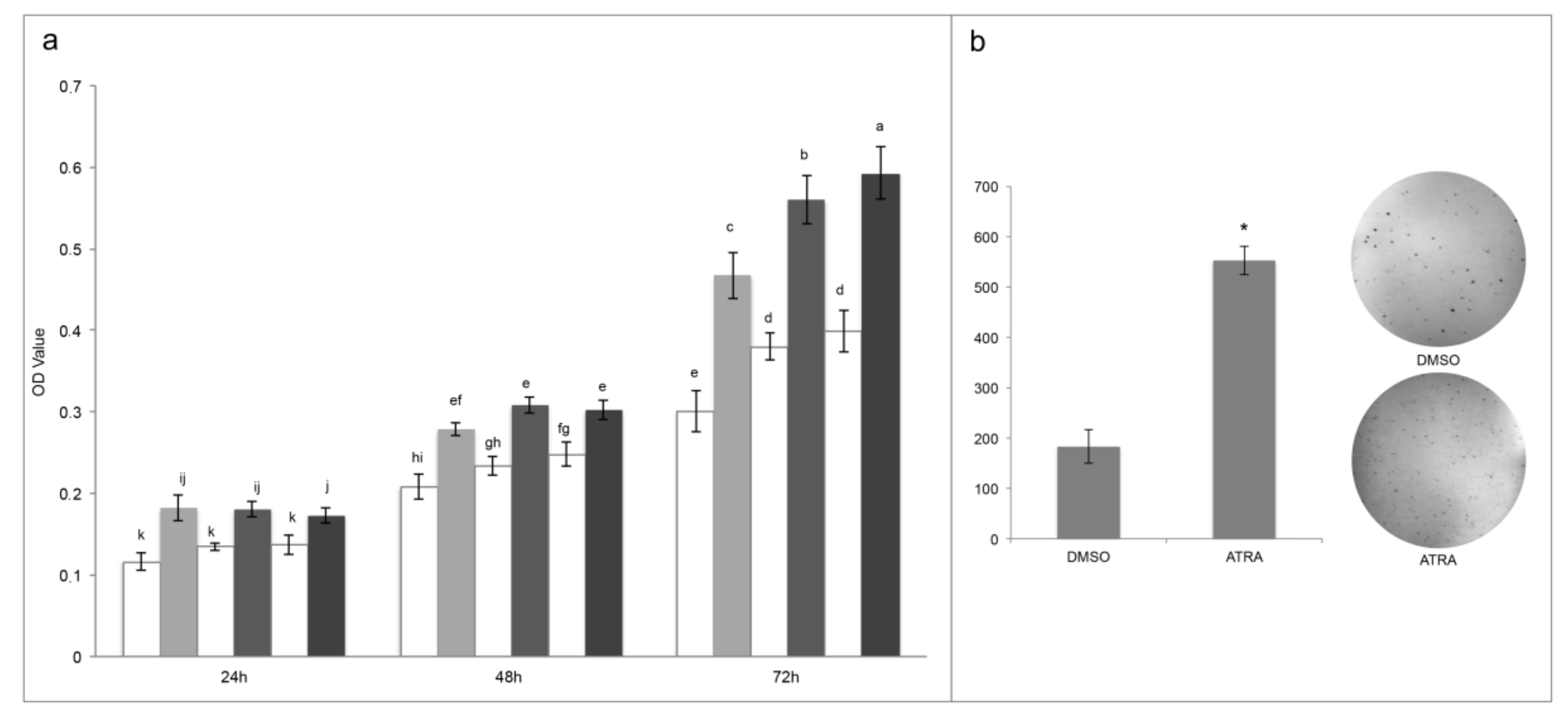
As confirmation of ATRA proliferative action in T24 cells a differential proteomic analysis was done comparing cells treated with DMSO and 10 µM ATRA for 72 h. Among the down-regulated proteins reasonably there were proteins belonging to the regulation of apoptosis pathway, but also proteins belonging to the Hippo-signaling pathway. In particular, there was two proteins belonging to the 14-3-3 family (YWHAB and YWHAE), which by binding to YAP and TAZ proteins in the cytosol are able to inactivate the pro-proliferative function of YAP/TAZ inducing cell contact-growth inhibition, necessary to the organ size control [
16]. Instead, among the up regulated proteins, beyond those implicated in the metabolism of proteins and in the post-translation protein modifications, obviously necessary to proliferation, others involved in the regulation of IGF transport and uptake by IGFBPs, and in the RAF/MAP kinase cascade were found. IGF exerts a pro-survival action stimulating the PI3k/Akt pathway, and other evidence in the literature showed that in ATRA-sensitive cancer cells, ATRA suppresses IGFs proliferative action, whilst in ATRA-resistant cell lines the IGFs proliferative action occurs for several modified molecular mechanism [
4]. In support of this data, previous papers showed the up regulation of the IGFs proliferation pathway in T24 cells in comparison to normal urothelial cells [
17,
18]. However, here for the first time, it was showed the up-regulation of the IGFs pathway following 10 µM ATRA treatment in T24 cells, which mostly supports the ATRA pro-proliferative action in this cellular model of bladder cancer. Similarly, also proteins of the RAF/MAP kinase pathway turned out to be up-regulated following ATRA treatment in T24 cells. Also in this case, RAF/MAP kinase pathway is inhibited by ATRA treatment in ATRA-sensitive cells, but in ATRA-resistant this pathway is functional and through c-jun and c-fos lead to cellular proliferation [
4]. However, the molecular mechanisms by which ATRA up or down regulated the described pathways in T24 cells need to be investigated.
As first step to understand the molecular mechanism behind the ATRA proliferative action in T24 cells, the cytoplasmic retinoid receptors were analyzed, considering that the same resistance found for ATRA did not occur with the synthetic retinoid 4-HRP, which has a cytotoxic mechanism independent from the classical retinoid receptor pathways (
Supplementary Figure S2). From the first literature evidence of T24 CRABP-II down-regulation [
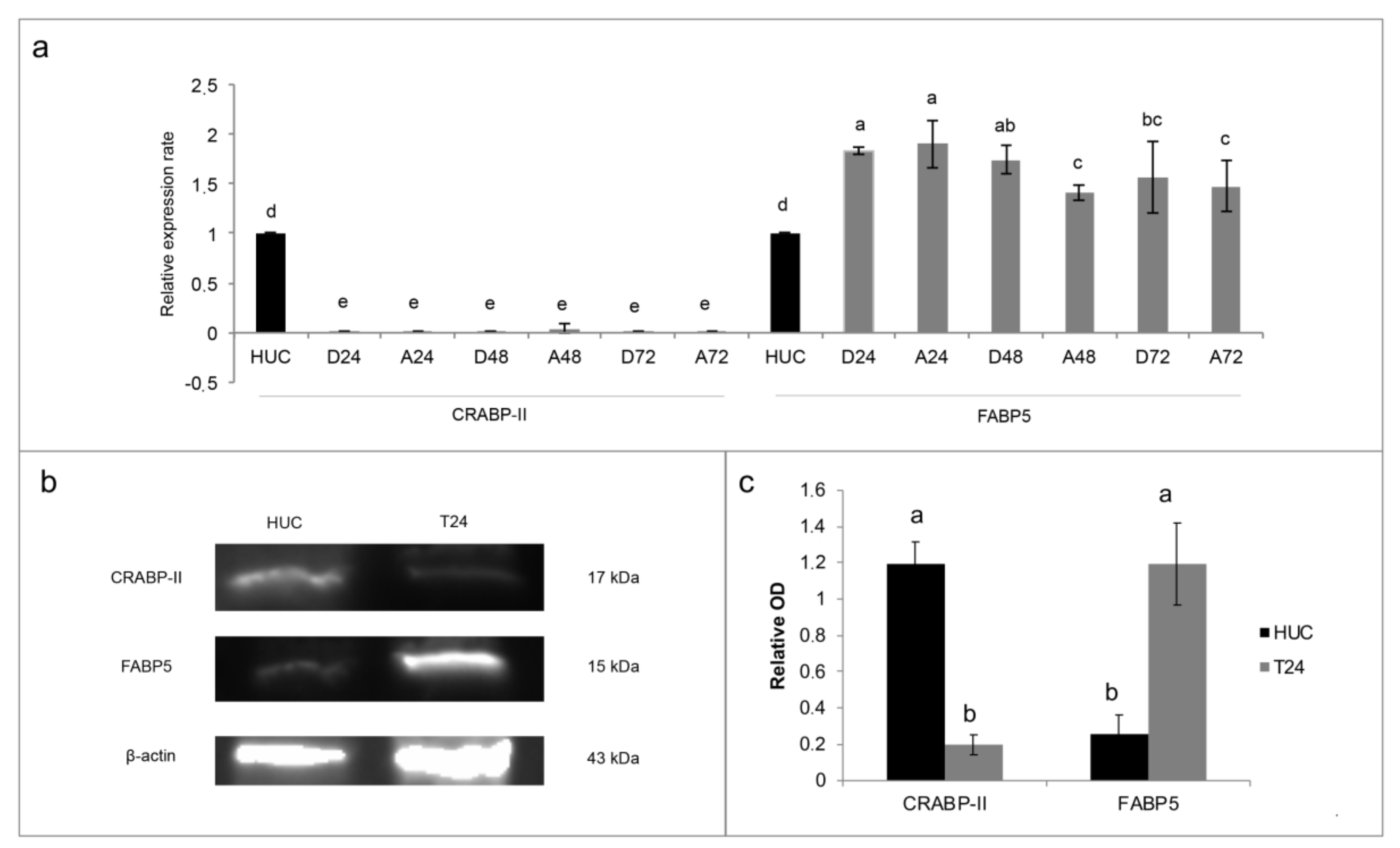
10], in the present paper the FABP5-PPARβ/δ pathway was for the first time analyzed in the T24 TCC cells and compared with the normal HUC cells. The data found and reported in
Figure 3 confirmed our hypothesis: the high FABP5/CRABP-II ratio in T24 cells, in comparison to HUC normal cells, could justify the activation of FABP5-PPARβ/δ non-genomic pathway, which was found to induce proliferation in other ATRA resistant solid cancer cells [
4,
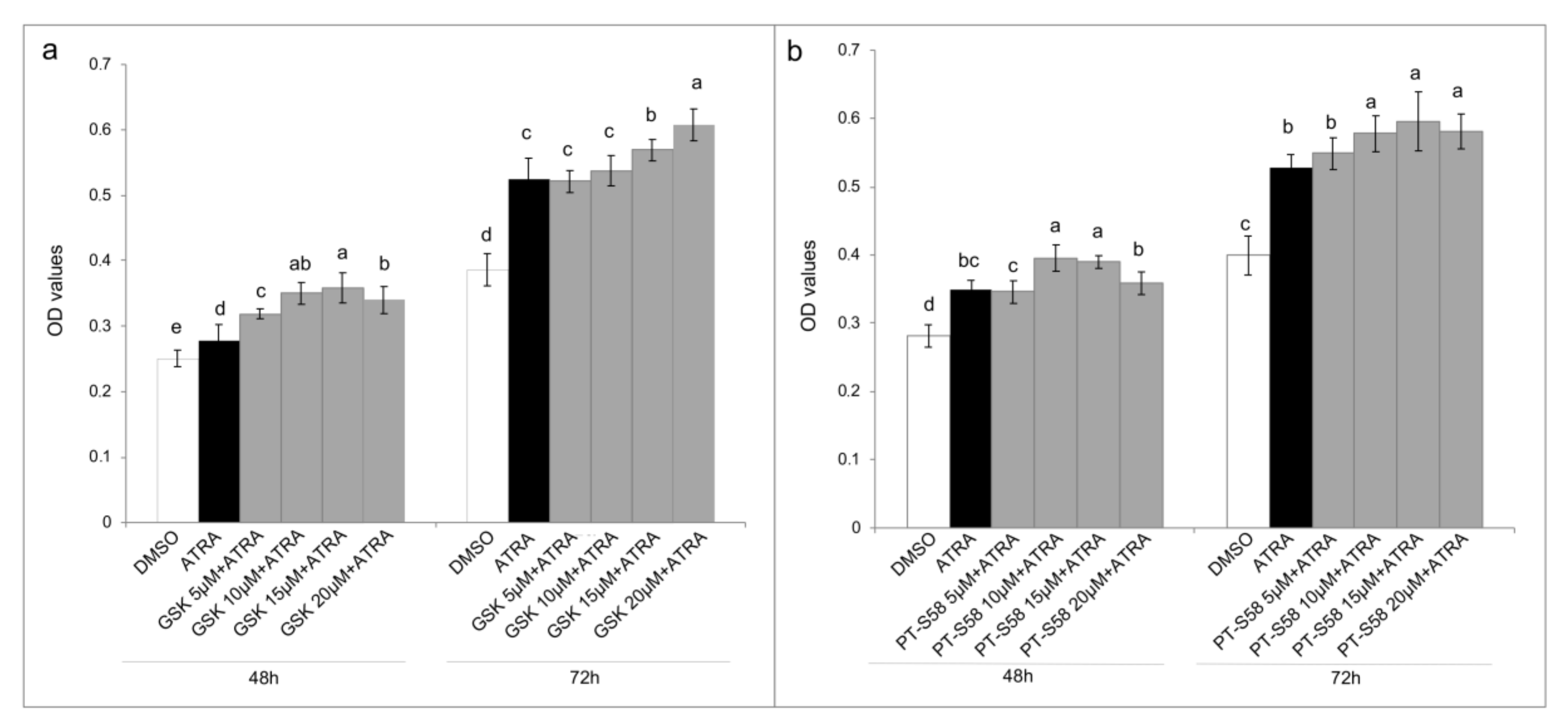
7]. For this reason, inhibition of PPARβ/δ receptor was attempted through the use of two different antagonists, GSK0660 and PTS-58. However, surprisingly, both the antagonists, dose-dependently and significantly, potentiated the viability increase (
Figure 4), even more when ATRA co-treatment was administered, indicating that probably different molecular mechanisms occur. Several assumptions can explain these results from the presence of mutations in the PPARβ/δ receptor, to the interactions with other mutant nuclear molecules that lead to cellular proliferation, until the presence of other nuclear pathways that do not require the PPARβ/δ involvement. In the literature some evidence is present in relation to RXRα [
19] and PPARγ [
20] genes mutations in bladder cancer that can cause PPARs/RXRα pathway hyperactivation resulting in the cellular proliferation, even though they were not related to ATRA treatments. Moreover, other molecular modifications associated with T24 TCC line and/or bladder cancer tissues have been found and they can be correlated with ATRA resistance as RARβ silencing by aberrant methylation [
21], and the absence of the estrogen receptor, or rather estrogen receptor negative (ER-) [
22,
23]. Finally, even though IGFs and RAF/MAP kinase pathways were here found up-regulated following 10 µM ATRA treatment, further analyses are needed to understand what actually stimulates the cellular proliferation at the nuclear level, especially for retinoids nuclear receptors.
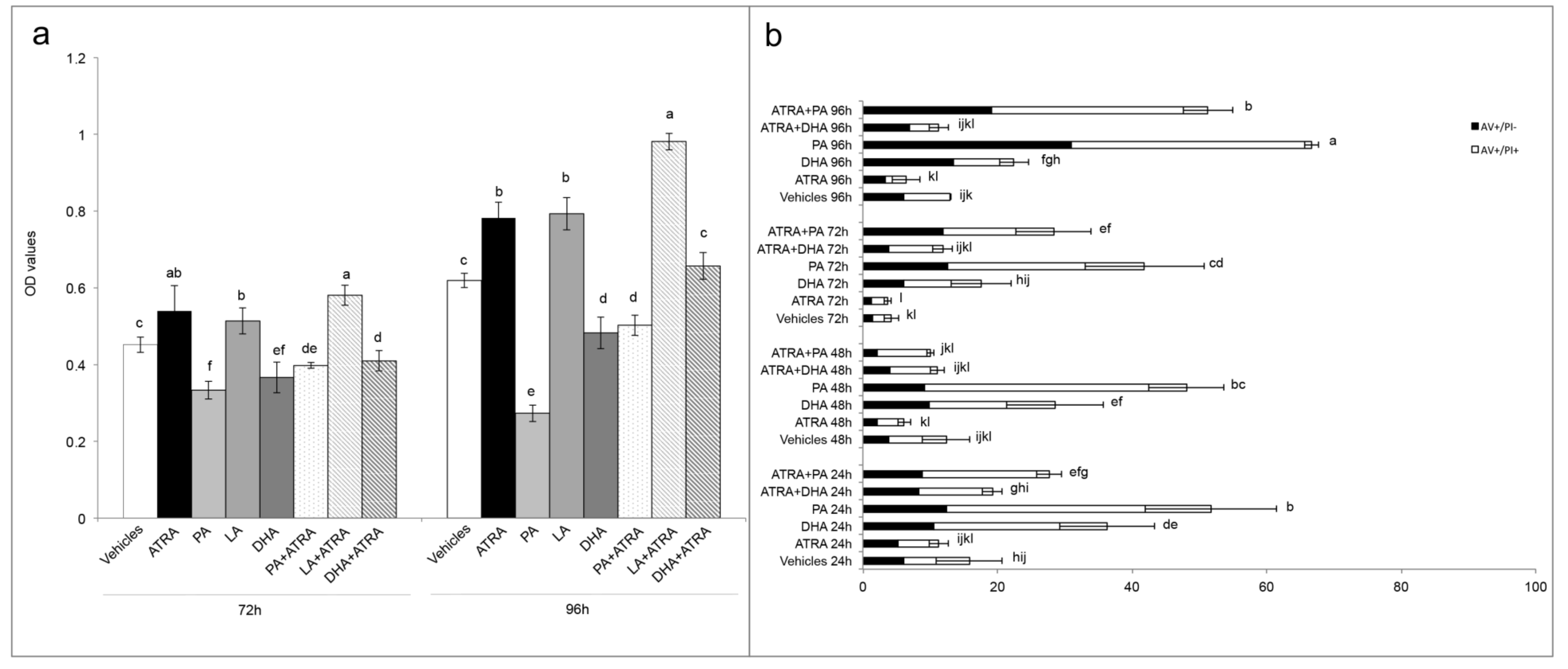
Finally, with the aim to better understand the molecular mechanism of ATRA in relation to FABP5-PPARβ/δ pathway, and in order to find an antagonist that inhibits the ATRA proliferative action, the roles of LCFA was investigated as ATRA co-treatment. Indeed, in the paper of Levi and colleagues [
13] was shown that saturated LCFA, in particular PA, is able to displace ATRA from FABP5, inhibiting its delivery to PPARβ/δ, and blocking FABP5 in the cytoplasm. In this way all the ATRA molecules in the cytosol can bind CRABP-II/RARs exerting its classical/genomic activity [
13]. Conversely, in the same paper, the authors stated that unsaturated LCFAs, in particular LA, could act similarly to ATRA, binding FABP5, triggering its translocation to the nucleus, and inducing PPARβ/δ activation. However, in the present paper we found that this is an incorrect generalization. Indeed, among the unsaturated LCFAs, DHA was not tested in the paper of Levi and co-workers. In particular, we found similar data for PA and LA, or rather we found that PA reduced viability when administered as co-treatment with ATRA, while LA potentiated the ATRA proliferative action (
Figure 5). However, the unsaturated DHA gave different results in comparison to LA, inhibiting the proliferative action of ATRA and giving viability percentages similar to control with vehicles (
Figure 5a). The cytotoxic action of DHA was also evident in the DHA single treatment, considering that it induced a significant viability decrease, and significant percentages of apoptosis (
Figure 5b). Although a previous paper of Lin and colleagues [
23] investigated the combined effect of ATRA and unsaturated LCFA, ALA, EPA, and DHA, on ER- ATRA resistant breast cancer cell lines, they used huge LCFA and ATRA concentrations (80 µM and 20 µM, respectively), and attributed the cytotoxic action to a synergistic action between unsaturated LCFA and ATRA. In the present paper, the reversion of ATRA proliferative action was obtained with the same concentration of DHA and ATRA (10 µM). Moreover, from the data it is possible to speculate that ATRA continues to carry out its proliferative action; however, this is counteracted by DHA co-treatment. Anyway, further investigations are needed to understand the molecular pathways implicated in these effects.

 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




