Peptidyl Arginine Deiminase Type 4 Gene Promoter Hypo-Methylation in Rheumatoid Arthritis


Abstract
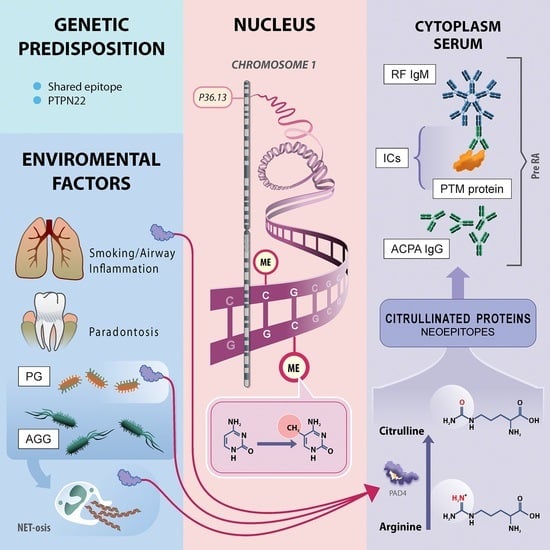
1. Introduction
2. Experimental Section
2.1. Patients
2.2. DNA Extraction and Methylation
2.3. Detection of ACPA and Anti-PAD4
2.4. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aletaha, D.; Alasti, F.; Smolen, J.S. Rheumatoid factor, not antibodies against citrullinated proteins, is associated with baseline disease activity in rheumatoid arthritis clinical trials. Arthritis Res. Ther. 2015, 17, 229. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham Iii, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.L.; Hunt, L.; Hensor, E.M.; Emery, P. Enriching case selection for imminent RA: The use of anti-CCP antibodies in individuals with new non-specific musculoskeletal symptoms—A cohort study. Ann. Rheum. Dis. 2016, 75, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- Sebbag, M.; Chapuy-Regaud, S.; Auger, I.; Petit-Texeira, E.; Clavel, C.; Nogueira, L.; Vincent, C.; Cornélis, F.; Roudier, J.; Serre, G. Clinical and pathophysiological significance of the autoimmune response to citrullinated proteins in rheumatoid arthritis. Jt. Bone Spine 2004, 71, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Yamada, R.; Chang, X.; Tokuhiro, S.; Sawada, T.; Suzuki, M.; Nagasaki, M.; Nakayama-Hamada, M.; Kawaida, R.; Ono, M. Functional haplotypes of PADI4, encoding citrullinating enzyme peptidylarginine deiminase 4, are associated with rheumatoid arthritis. Nat. Genet. 2003, 34, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.; Lugonja, B.; Jimmy Ytterberg, A.; Zubarev, R.A.; Creese, A.J.; Pearson, M.J.; Grant, M.M.; Milward, M.; Lundberg, K.; Buckley, C.D. Release of active peptidyl arginine deiminases by neutrophils can explain production of extracellular citrullinated autoantigens in rheumatoid arthritis synovial fluid. Arthritis Rheumatol. 2015, 67, 3135–3145. [Google Scholar] [CrossRef]
- Delgado-Vega, A.; Sánchez, E.; Löfgren, S.; Castillejo-López, C.; Alarcón-Riquelme, M.E. Recent findings on genetics of systemic autoimmune diseases. Curr. Opin. Immunol. 2010, 22, 698–705. [Google Scholar] [CrossRef]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res. 2019, 47, D745–D751. [Google Scholar] [CrossRef]
- Darrah, E.; Giles, J.T.; Ols, M.L.; Bull, H.G.; Andrade, F.; Rosen, A. Erosive rheumatoid arthritis is associated with antibodies that activate PAD4 by increasing calcium sensitivity. Sci. Transl. Med. 2013, 5, 186ra165. [Google Scholar] [CrossRef] [PubMed]
- Arita, K.; Hashimoto, H.; Shimizu, T.; Nakashima, K.; Yamada, M.; Sato, M. Structural basis for Ca2+-induced activation of human PAD4. Nat. Struct. Mol. Biol. 2004, 11, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Seaman, A.; Darrah, E.; Infantino, M.; Meacci, F.; Manfredi, M.; Benucci, M.; Mahler, M. Anti-peptidyl-arginine deaminase 3 (PAD3) antibodies as a promising marker to measure joint damage in patients with rheumatoid arthritis. Autoimmun. Rev. 2016, 15, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; van de Stadt, L.A.; Levarht, E.N.; Huizinga, T.W.; Hamann, D.; van Schaardenburg, D.; Toes, R.E.; Trouw, L.A. Anti-carbamylated protein (anti-CarP) antibodies precede the onset of rheumatoid arthritis. Ann. Rheum. Dis. 2014, 73, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Majka, D.S.; Deane, K.D.; Parrish, L.A.; Lazar, A.A.; Barn, A.E.; Walker, C.W.; Rubertone, M.V.; Gilliland, W.R.; Norris, J.M.; Holers, V.M. Duration of preclinical rheumatoid arthritis-related autoantibody positivity increases in subjects with older age at time of disease diagnosis. Ann. Rheum. Dis. 2008, 67, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Sokolove, J.; Johnson, D.S.; Lahey, L.J.; Wagner, C.A.; Cheng, D.; Thiele, G.M.; Michaud, K.; Sayles, H.; Reimold, A.M.; Caplan, L.; et al. Rheumatoid factor as a potentiator of anti-citrullinated protein antibody-mediated inflammation in rheumatoid arthritis. Arthritis Rheumatol. 2014, 66, 813–821. [Google Scholar] [CrossRef]
- Oppermann, U. Why is epigenetics important in understanding the pathogenesis of inflammatory musculoskeletal diseases. Arthritis Res. Ther. 2013, 15, 209. [Google Scholar] [CrossRef]
- Liu, Y.; Aryee, M.J.; Padyukov, L.; Fallin, M.D.; Hesselberg, E.; Runarsson, A.; Reinius, L.; Acevedo, N.; Taub, M.; Ronninger, M.; et al. Epigenome-wide association data implicate DNA methylation as an intermediary of genetic risk in rheumatoid arthritis. Nat. Biotechnol. 2013, 31, 142–147. [Google Scholar] [CrossRef]
- Padyukov, L.; Seielstad, M.; Ong, R.; Ding, B.; Ronnelid, J.; Seddighzadeh, M.; Alfredsson, L.; Klareskog, L. Epidemiological Investigation of Rheumatoid Arthritis study g. A genome-wide association study suggests contrasting associations in ACPA-positive versus ACPA-negative rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 259–265. [Google Scholar] [CrossRef]
- Xu, X.Y.; Wang, M.M.; Xiao, C.S.; LI, X.F.; Wang, L.Y. The study of genome DNA methylation in rheumatoid arthritis. Chin. J. Rheumatol. 2006, 10, 462–465. [Google Scholar]
- Zhu, H.; Wu, L.-F.; Mo, X.-B.; Lu, X.; Tang, H.; Zhu, X.-W.; Xia, W.; Guo, Y.-F.; Wang, M.-J.; Zeng, K.-Q.; et al. Rheumatoid arthritis–associated DNA methylation sites in peripheral blood mononuclear cells. Ann. Rheum. Dis. 2019, 78, 36–42. [Google Scholar] [CrossRef]
- Fransen, J.; van Riel, P.L. The Disease Activity Score and the EULAR response criteria. Clin. Exp. Rheumatol. 2005, 23, S93–S99. [Google Scholar] [CrossRef]
- Dreos, R.; Ambrosini, G.; Périer, R.C.; Bucher, P. The Eukaryotic Promoter Database: Expansion of EPDnew and new promoter analysis tools. Nucleic Acids Res. 2014, 43, D92–D96. [Google Scholar] [CrossRef] [PubMed]
- Davidović, R.S.; Božović, A.M.; Mandušić, V.L.; Krajnović, M.M. Methylation-specific PCR: Four steps in primer design. Cent. Eur. J. Biol. 2014, 9, 1127–1139. [Google Scholar] [CrossRef]
- Menigatti, M.; Truninger, K.; Gebbers, J.; Marbet, U.; Marra, G.; Schär, P. Normal colorectal mucosa exhibits sex-and segment-specific susceptibility to DNA methylation at the hMLH1 and MGMT promoters. Oncogene 2009, 28, 899–909. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R. Epigenetics in autoimmune diseases: Pathogenesis and prospects for therapy. Autoimmun. Rev. 2015, 14, 854–863. [Google Scholar] [CrossRef]
- Viatte, S.; Plant, D.; Raychaudhuri, S. Genetics and epigenetics of rheumatoid arthritis. Nat. Rev. Rheumatol. 2013, 9, 141–153. [Google Scholar] [CrossRef]
- Plant, D.; Webster, A.; Nair, N.; Oliver, J.; Smith, S.L.; Eyre, S.; Hyrich, K.L.; Wilson, A.G.; Morgan, A.W.; Isaacs, J.D.J.A.; et al. Differential methylation as a biomarker of response to etanercept in patients with rheumatoid arthritis. Arthritis Rheumatol. 2016, 68, 1353–1360. [Google Scholar] [CrossRef]
- Nair, N.; Plant, D.; Verstappen, S.M.; Isaacs, J.D.; Morgan, A.W.; Hyrich, K.L.; Barton, A.; Wilson, A.G.; the MATURA investigators. Differential DNA methylation correlates with response to methotrexate in rheumatoid arthritis. Rheumatology 2020, 59, 1364–1371. [Google Scholar] [CrossRef]
- Kolarz, B.; Majdan, M. Epigenetic aspects of rheumatoid arthritis: Contribution of non-coding RNAs. Semin. Arthritis Rheum. 2017, 46, 724–731. [Google Scholar] [CrossRef]
- Meng, W.; Zhu, Z.; Jiang, X.; Too, C.L.; Uebe, S.; Jagodic, M.; Kockum, I.; Murad, S.; Ferrucci, L.; Alfredsson, L. DNA methylation mediates genotype and smoking interaction in the development of anti-citrullinated peptide antibody-positive rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 71. [Google Scholar] [CrossRef] [PubMed]
- Wegner, N.; Wait, R.; Sroka, A.; Eick, S.; Nguyen, K.A.; Lundberg, K.; Kinloch, A.; Culshaw, S.; Potempa, J.; Venables, P.J. Peptidylarginine deiminase from Porphyromonas gingivalis citrullinates human fibrinogen and α-enolase: Implications for autoimmunity in rheumatoid arthritis. Arthritis Rheum. 2010, 62, 2662–2672. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Millán, I.; Darrah, E.; Westfall, A.O.; Mikuls, T.R.; Reynolds, R.J.; Danila, M.I.; Curtis, J.R.; Rosen, A.; Bridges, S.L. Association of anti-peptidyl arginine deiminase antibodies with radiographic severity of rheumatoid arthritis in African Americans. Arthritis Res. Ther. 2016, 18, 241. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, E.H.; Pollmann, S.; Gilboe, I.M.; Van Der Heijde, D.; Landewé, R.; Ødegård, S.; Kvien, T.K.; Molberg, Ø. Serum IgG antibodies to peptidylarginine deiminase 4 in rheumatoid arthritis and associations with disease severity. Ann. Rheum. Dis. 2008, 67, 414–417. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, Y.; He, J.; Jia, R.; Li, Z. Prevalence and significance of anti-peptidylarginine deiminase 4 antibodies in rheumatoid arthritis. J. Rheumatol. 2008, 35, 969–974. [Google Scholar]
- Martinez-Prat, L.; Lucia, D.; Ibarra, C.; Mahler, M.; Dervieux, T. Antibodies targeting protein-arginine deiminase 4 (PAD4) demonstrate diagnostic value in rheumatoid arthritis. Ann. Rheum. Dis. 2019, 78, 434–436. [Google Scholar] [CrossRef]
- Guderud, K.; Mæhlen, M.T.; Nordang, G.B.N.; Viken, M.K.; Andreassen, B.K.; Molberg, Ø.; Flåm, S.T.; Lie, B.A. Lack of association among peptidyl arginine deiminase type 4 autoantibodies, PADI4 polymorphisms, and clinical characteristics in rheumatoid arthritis. J. Rheumatol. 2018, 45, 1211–1219. [Google Scholar] [CrossRef]
- Halvorsen, E.H.; Haavardsholm, E.A.; Pollmann, S.; Boonen, A.; van der Heijde, D.; Kvien, T.K.; Molberg, Ø. Serum IgG antibodies to peptidylarginine deiminase 4 predict radiographic progression in patients with rheumatoid arthritis treated with tumour necrosis factor-α blocking agents. Ann. Rheum. Dis. 2009, 68, 249–252. [Google Scholar] [CrossRef]
- Paul, B.J.; Kandy, H.I.; Krishnan, V. Pre-rheumatoid arthritis and its prevention. Eur. J. Rheumatol. 2017, 4, 161–165. [Google Scholar] [CrossRef]
- Makrygiannakis, D.; Hermansson, M.; Ulfgren, A.K.; Nicholas, A.P.; Zendman, A.J.; Eklund, A.; Grunewald, J.; Skold, C.M.; Klareskog, L.; Catrina, A.I. Smoking increases peptidylarginine deiminase 2 enzyme expression in human lungs and increases citrullination in BAL cells. Ann. Rheum. Dis. 2008, 67, 1488–1492. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Totaro, M.C.; Cattani, P.; Ria, F.; Tolusso, B.; Gremese, E.; Fedele, A.L.; D’Onghia, S.; Marchetti, S.; Di Sante, G.; Canestri, S. Porphyromonas gingivalis and the pathogenesis of rheumatoid arthritis: Analysis of various compartments including the synovial tissue. Arthritis Res. Ther. 2013, 15, R66. [Google Scholar] [CrossRef] [PubMed]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T. Aggregatibacter actinomycetemcomitans–induced hypercitrullination links periodontal infection to autoimmunity in rheumatoid arthritis. Sci. Transl. Med. 2016, 8, 369ra176. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, D.; Butcher, J.; Culshaw, S. Periodontitis, Porphyromonas, and the pathogenesis of rheumatoid arthritis. Mucosal Immunol. 2012, 5, 112–120. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | RA | HC |
|---|---|---|
| N = 125 | N = 30 | |
| Age; mean (SD) | 52.2 (12.3) | 53.2 (8.1) |
| Females; n (%) | 103 (82.4) | 23 (76.7) |
| Disease duration [years]; n (SD) | 11.69 (9.3) | n/a |
| Rheumatoid Factor positive; n (%), | 87 (69.6) | none |
| anti-PAD4 positive; n (%) | 67 (53.6) | 2 (6.7) |
| ACPA positive; n (%) | 104 (83.2) | none |
| ESR; mean (SD) | 31.5 (24.4) | 14.6 (9.2) |
| CRP [mg/dL]; mean (SD) | 13.96 (25.28) | 1.37 (1.71) |
| VAS PGA; mean (SD) | 27.1 (27.2) | n/a |
| VAS PhGA; mean (SD) | 21.7 (20.6) | n/a |
| Treatment | n/a | |
| At least Methotrexate, n (%) | 90 (72) | |
| At least Biologics, n (%) | 45 (35.2) | |
| At least Steroids, n (%) | 73 (58.4) | |
| Single drug therapy, n (%) | 45 (36) | |
| Metotrexate, n (%) | 23 (18.4) | |
| Biologics, n (%) | 5 (4) | |
| Steroids, n (%) | 17 (13.6) | |
| Double drug therapy, n (%) | 50 (40) | |
| At least Methotrexate + Steroids, n (%) | 51 (40.8) | |
| At least Methotrexate + Biologics, n (%) | 36 (28.8) | |
| At least Steroids + Biologics, n (%) | 25 (20) | |
| Methotrexate + Steroids, n (%) | 31 (24.8) | |
| Methotrexate + Biologics, n (%) | 15 (12) | |
| Steroids + Biologics, n (%) | 4 (3.2) | |
| Triple drug therapy, n (%) | 21 (16.8) | |
| No therapy, n (%) | 9 (7.2) |
| RA Severe DAS28 > 5.1 (n = 34; 27.2%) | RA Moderate DAS28 > 3.2–5.1 (n = 46; 36.8%) | RA Low DAS28 > 2.6–3.2 (n = 19; 15.2%) | RA Remission DAS28 ≤ 2.6 (n = 26; 20.8%) | RA Overall n = 125 | Healthy Control n = 30 | |
|---|---|---|---|---|---|---|
| PADI4 Metylation [fold change] (methylated sequences) * | 1.19 [0.59–1.9] | 1.06 [0.54–2.96] | 1.66 [1.01–3.22] | 2.53 [1.47–4.45] | 1.46 [0.68–3.07] | 5.17 [2.77–8.74] |
| anti-PAD4 [U/mL] * | 731.03 [477.98–1288.1] | 716.11 [400.85–1250.8] | 589.57 [306.08–2380.2] | 455.54 [271.26–1104.9] | 681.39 [339.36–1288.1] | 293.25 [234.39–389.12] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolarz, B.; Ciesla, M.; Dryglewska, M.; Majdan, M. Peptidyl Arginine Deiminase Type 4 Gene Promoter Hypo-Methylation in Rheumatoid Arthritis. J. Clin. Med. 2020, 9, 2049. https://doi.org/10.3390/jcm9072049
Kolarz B, Ciesla M, Dryglewska M, Majdan M. Peptidyl Arginine Deiminase Type 4 Gene Promoter Hypo-Methylation in Rheumatoid Arthritis. Journal of Clinical Medicine. 2020; 9(7):2049. https://doi.org/10.3390/jcm9072049
Chicago/Turabian StyleKolarz, Bogdan, Marek Ciesla, Magdalena Dryglewska, and Maria Majdan. 2020. "Peptidyl Arginine Deiminase Type 4 Gene Promoter Hypo-Methylation in Rheumatoid Arthritis" Journal of Clinical Medicine 9, no. 7: 2049. https://doi.org/10.3390/jcm9072049
APA StyleKolarz, B., Ciesla, M., Dryglewska, M., & Majdan, M. (2020). Peptidyl Arginine Deiminase Type 4 Gene Promoter Hypo-Methylation in Rheumatoid Arthritis. Journal of Clinical Medicine, 9(7), 2049. https://doi.org/10.3390/jcm9072049