DNA Fragmentation in Viable and Non-Viable Spermatozoa Discriminates Fertile and Subfertile Subjects with Similar Accuracy



 ,
, 
Abstract
1. Introduction
2. Material and Methods
2.1. Subjects and Sample Collection and Preparation
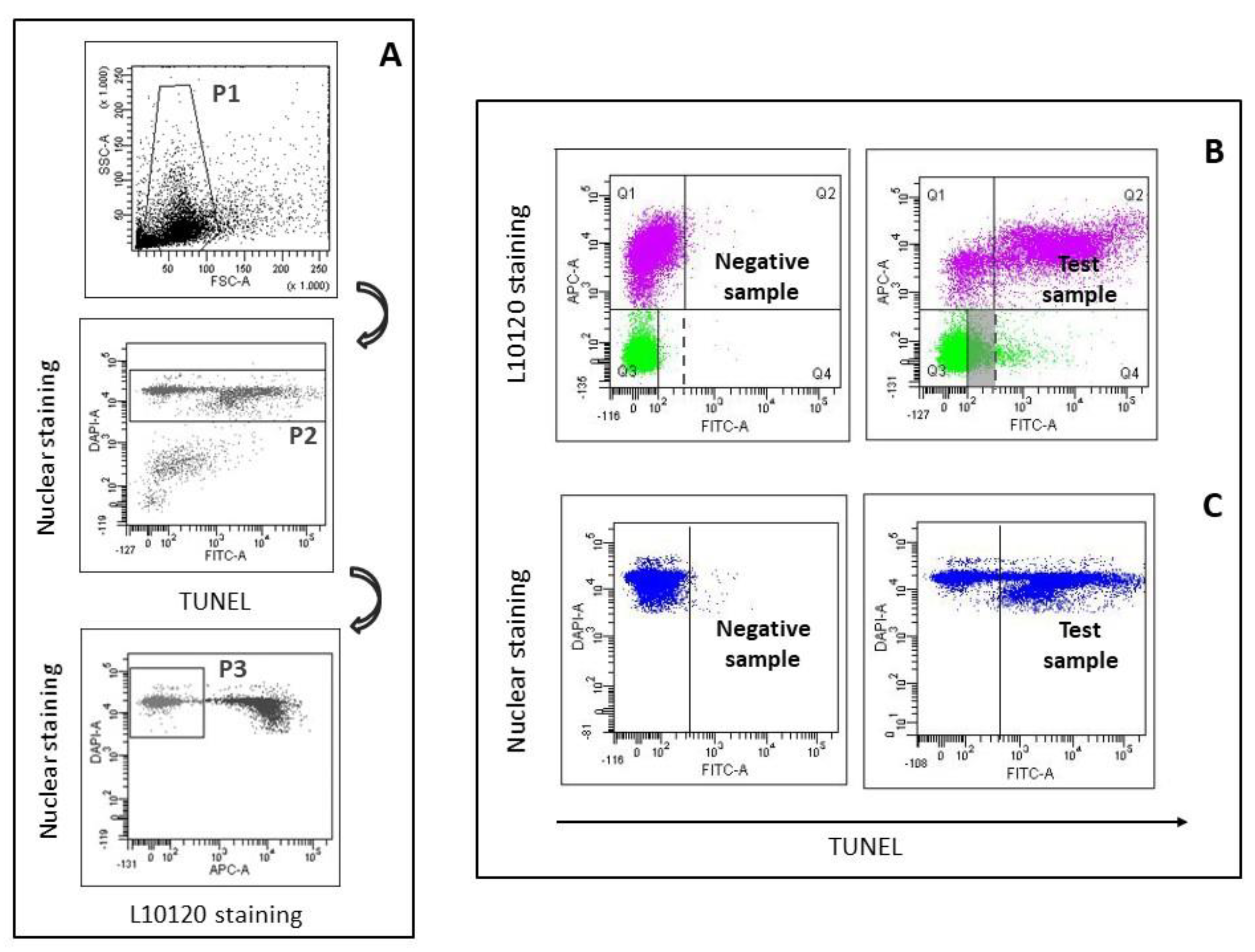
2.2. LiveTUNEL
2.3. Flow Cytometric Analyses
2.4. Statistical Analysis
3. Results
3.1. LiveTUNEL
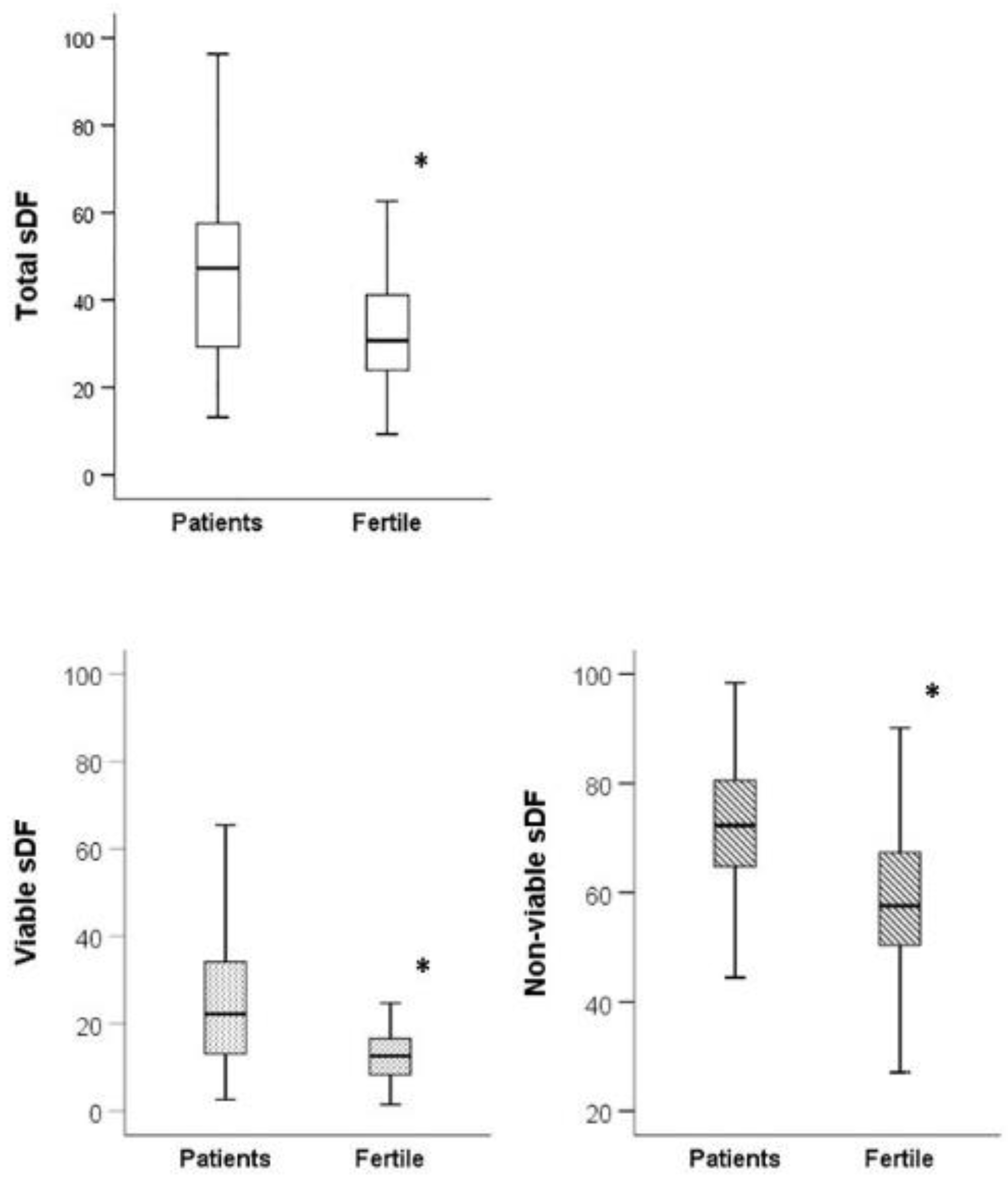
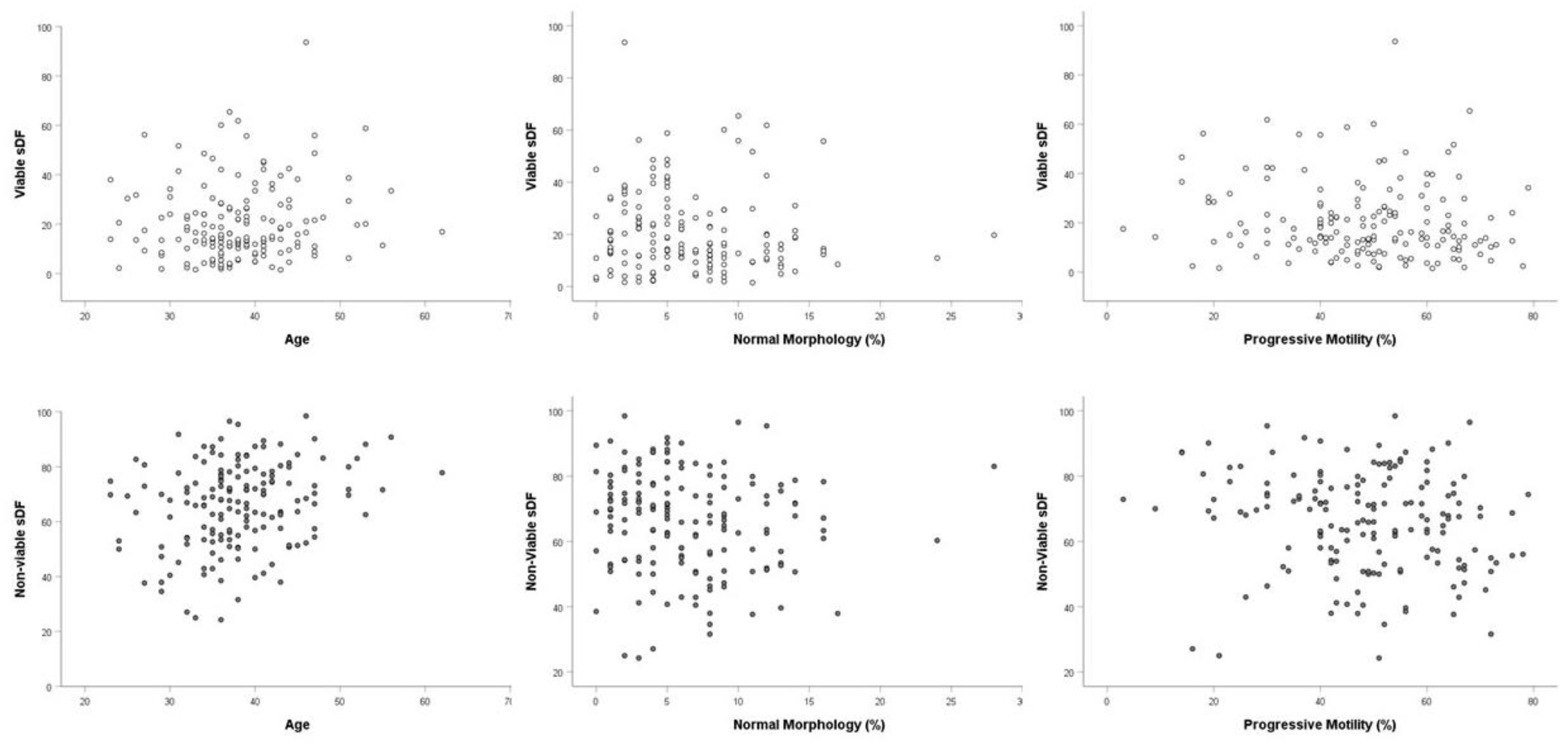
3.2. sDF in Fertile Men and in Patients
3.3. SDF as a Predictor of Fertile versus Subfertile Status
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Levine, H.; Jørgensen, N.; Martino-Andrade, A.; Mendiola, J.; Weksler-Derri, D.; Mindlis, I.; Pinotti, R.; Swan, S.H. Temporal trends in sperm count: A systematic review and meta-regression analysis. Hum. Reprod. Update 2017, 23, 646–659. [Google Scholar] [CrossRef] [PubMed]
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jørgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male Reproductive Disorders and Fertility Trends: Influences of Environment and Genetic Susceptibility. Physiol. Rev. 2016, 96, 55–97. [Google Scholar] [CrossRef] [PubMed]
- Carré, J.; Gatimel, N.; Moreau, J.; Parinaud, J.; Léandri, R. Does air pollution play a role in infertility? A systematic review. Environ. Health 2017, 16, 82. [Google Scholar] [CrossRef] [PubMed]
- Ilacqua, A.; Izzo, G.; Emerenziani, G.P.; Baldari, C.; Aversa, A. Lifestyle and fertility: The influence of stress and quality of life on male fertility. Reprod. Biol. Endocrinol. 2018, 16, 115. [Google Scholar] [CrossRef] [PubMed]
- Hayden, R.P.; Flannigan, R.; Schlegel, P.N. The Role of Lifestyle in Male Infertility: Diet, Physical Activity, and Body Habitus. Curr. Urol. Rep. 2018, 19, 56. [Google Scholar] [CrossRef] [PubMed]
- Muratori, M.; Marchiani, S.; Tamburrino, L.; Baldi, E. Sperm DNA Fragmentation: Mechanisms of Origin. Adv. Exp. Med. Biol. 2019, 1166, 75–85. [Google Scholar]
- Muratori, M.; Tamburrino, L.; Marchiani, S.; Cambi, M.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E. Investigation on the Origin of Sperm DNA Fragmentation: Role of Apoptosis, Immaturity and Oxidative Stress. Mol. Med. 2015, 21, 109–122. [Google Scholar] [CrossRef]
- Evenson, D.P.; Jost, L.K.; Marshall, D.; Zinaman, M.J.; Clegg, E.; Purvis, K.; de Angelis, P.; Claussen, O.P. Utility of the sperm chromatin structure assay as a diagnostic and prognostic tool in the human fertility clinic. Hum. Reprod. 1999, 14, 1039–1049. [Google Scholar] [CrossRef]
- Giwercman, A.; Lindstedt, L.; Larsson, M.; Bungum, M.; Spano, M.; Levine, R.J.; Rylander, L. Sperm chromatin structure assay as an independent predictor of fertility in vivo: A case-control study. Int. J. Androl. 2010, 33, e221–e227. [Google Scholar] [CrossRef]
- Muratori, M.; Marchiani, S.; Tamburrino, L.; Cambi, M.; Lotti, F.; Natali, I.; Filimberti, E.; Noci, I.; Forti, G.; Maggi, M.; et al. DNA fragmentation in brighter sperm predicts male fertility independently from age and semen parameters. Fertil. Steril. 2015, 104, 582–590.e4. [Google Scholar] [CrossRef]
- Borini, A.; Tarozzi, N.; Bizzaro, D.; Bonu, M.A.; Fava, L.; Flamigni, C.; Coticchio, G. Sperm DNA fragmentation: Paternal effect on early post-implantation embryo development in ART. Hum. Reprod. 2006, 21, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.; Gallos, I.D.; Conner, S.J.; Rajkhowa, M.; Miller, D.; Lewis, S.; Kirkman-Brown, J.; Coomarasamy, A. The effect of sperm DNA fragmentation on miscarriage rates: A systematic review and meta-analysis. Hum. Reprod. 2012, 27, 2908–2917. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Taskin, O.; Albert, A.; Bedaiwy, M.A. Association between sperm DNA fragmentation and idiopathic recurrent pregnancy loss: A systematic review and meta-analysis. Reprod. Biomed. Online 2019, 38, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Cairo Consensus Workshop Group. The current status and future of andrology: A consensus report from the Cairo workshop group. Andrology 2020, 8, 27–52. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, L.A.; De Iuliis, G.N.; Aitken, R.J. The TUNEL assay consistently underestimates DNA damage in human spermatozoa and is influenced by DNA compaction and cell vitality: Development of an improved methodology. Int. J. Androl. 2011, 34, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Drevet, J.R.; Aitken, R.J. Oxidative Damage to Sperm DNA: Attack and Defense. Adv. Exp. Med. Biol. 2019, 1166, 107–117. [Google Scholar]
- Adewoyin, M.; Ibrahim, M.; Roszaman, R.; Isa, M.L.M.; Alewi, N.A.M.; Rafa, A.A.A.; Anuar, M.N.N. Male Infertility: The Effect of Natural Antioxidants and Phytocompounds on Seminal Oxidative Stress. Reprod. Biomed. Online 2012, 25, 300–306. [Google Scholar] [CrossRef]
- Kesari, K.K.; Agarwal, A.; Henkel, R. Radiations and male fertility. Reprod. Biol. Endocrinol. 2018, 16, 118. [Google Scholar] [CrossRef]
- La Vignera, S.; Condorelli, R.; Vicari, E.; D’Agata, R.; Calogero, A.E. Diabetes mellitus and sperm parameters. J. Androl. 2012, 33, 145–153. [Google Scholar] [CrossRef]
- Agarwal, A.; Sharma, R.K.; Desai, N.R.; Prabakaran, S.; Tavares, A.; Sabanegh, E. Role of oxidative stress in pathogenesis of varicocele and infertility. Urology 2009, 73, 461–469. [Google Scholar] [CrossRef]
- Belloc, S.; Hazout, A.; Zini, A.; Merviel, P.; Cabry, R.; Chahine, H.; Copin, H.; Benkhalifa, M. How to overcome male infertility after 40: Influence of paternal age on fertility. Maturitas 2014, 78, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Fatehi, A.N.; Bevers, M.M.; Schoevers, E.; Roelen, B.A.; Colenbrander, B.; Gadella, B.M. DNA damage in bovine sperm does not block fertilization and early embryonic development but induces apoptosis after the first cleavages. J. Androl. 2006, 27, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Borges, E., Jr.; Zanetti, B.F.; Setti, A.S.; Braga, D.P.A.F.; Provenza, R.R.; Iaconelli, A., Jr. Sperm DNA fragmentation is correlated with poor embryo development, lower implantation rate, and higher miscarriage rate in reproductive cycles of non-male factor infertility. Fertil. Steril. 2019, 112, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Alvarez Sedó, C.; Bilinski, M.; Lorenzi, D.; Uriondo, H.; Noblía, F.; Longobucco, V.; Lagar, E.V.; Nodar, F. Effect of sperm DNA fragmentation on embryo development: Clinical and biological aspects. JBRA Assist. Reprod. 2017, 21, 343–350. [Google Scholar]
- World Health Organization. Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO Press: Geneva, Switzerland, 2010. [Google Scholar]
- Muratori, M.; Tarozzi, N.; Carpentiero, F.; Danti, S.; Perrone, F.M.; Cambi, M.; Casini, A.; Azzari, C.; Boni, L.; Maggi, M.; et al. Sperm selection with density gradient centrifugation and swim up: Effect on DNA fragmentation in viable spermatozoa. Sci. Rep. 2019, 9, 7492. [Google Scholar] [CrossRef]
- Muratori, M.; Tamburrino, L.; Tocci, V.; Costantino, A.; Marchiani, S.; Giachini, C.; Laface, I.; Krausz, C.; Meriggiola, M.C.; Forti, G.; et al. Small variations in crucial steps of TUNEL assay coupled to flow cytometry greatly affect measures of sperm DNA fragmentation. J. Androl. 2010, 31, 336–345. [Google Scholar] [CrossRef]
- Muratori, M.; Marchiani, S.; Tamburrino, L.; Tocci, V.; Failli, P.; Forti, G.; Baldi, E. Nuclear staining identifies two populations of human sperm with different DNA fragmentation extent and relationship with semen parameters. Hum. Reprod. 2008, 23, 1035–1043. [Google Scholar] [CrossRef]
- Marchiani, S.; Tamburrino, L.; Maoggi, A.; Vannelli, G.B.; Forti, G.; Baldi, E.; Muratori, M. Characterization of M540 bodies in human semen: Evidence that they are apoptotic bodies. Mol. Hum. Reprod. 2007, 13, 621–631. [Google Scholar] [CrossRef]
- Aitken, R.J.; De Iuliis, G.N.; Finnie, J.M.; Hedges, A.; McLachlan, R.I. Analysis of the relationships between oxidative stress, DNA damage and sperm vitality in a patient population: Development of diagnostic criteria. Hum. Reprod. 2010, 25, 2415–2426. [Google Scholar] [CrossRef]
- Muratori, M.; Forti, G.; Baldi, E. M540 bodies interfere with TUNEL analyses in human semen samples. Hum. Reprod. 2011, 26, 729. [Google Scholar] [CrossRef][Green Version]
- Agarwal, A.; Rana, M.; Qiu, E.; AlBunni, H.; Bui, A.D.; Henkel, R. Role of oxidative stress, infection and inflammation in male infertility. Andrologia 2018, 50, e13126. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Encinas, A.; García-Peiró, A.; Ribas-Maynou, J.; Abad, C.; Amengual, M.J.; Navarro, J.; Benet, J. Characterization of Nuclease Activity in Human Seminal Plasma and its relationship to Semen Parameters, Sperm DNA Fragmentation and Male Infertility. J. Urol. 2016, 195, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Arroyo, F.; Gosálvez, J. Dynamic assessment of human sperm DNA damage I: The effect of seminal plasma-sperm co-incubation after ejaculation. Int. Urol. Nephrol. 2018, 50, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Muratori, M.; Maggi, M.; Spinelli, S.; Filimberti, E.; Forti, G.; Baldi, E. Spontaneous DNA fragmentation in swim-up selected human spermatozoa during long term incubation. J. Androl. 2003, 24, 253–262. [Google Scholar] [CrossRef]
- Eisenberg, M.L.; Meldrum, D. Effects of age on fertility and sexual function. Fertil. Steril. 2017, 107, 301–304. [Google Scholar] [CrossRef]
- Evenson, D.P.; Larson, K.J.; Jost, L.K. Sperm Chromatin Structure Assay: Its clinical use for detecting sperm DNA fragmentation in male infertility and comparisons with other techniques. J. Androl. 2002, 23, 25–43. [Google Scholar] [CrossRef]
- Aitken, R.J.; Bronson, R.; Smith, T.B.; De Iuliis, G.N. The source and significance of DNA damage in human spermatozoa; a commentary on diagnostic strategies and straw man fallacies. Mol. Hum. Reprod. 2013, 19, 475–485. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Muratori, M.; De Geyter, M.; De Geyter, C. TUNEL labeling with BrdUTP/anti-BrdUTP greatly underestimates the level of sperm DNA fragmentation in semen evaluation. PLoS ONE 2017, 12, e0181802. [Google Scholar] [CrossRef]
- Marchiani, S.; Tamburrino, L.; Giuliano, L.; Nosi, D.; Sarli, V.; Gandini, L.; Piomboni, P.; Belmonte, G.; Forti, G.; Baldi, E.; et al. Sumo1-ylation of human spermatozoa and its relationship with semen quality. Int. J. Androl. 2011, 34, 581–593. [Google Scholar] [CrossRef]
- Baccetti, B.; Collodel, G.; Piomboni, P. Apoptosis in human ejaculated sperm cells (notulae seminologicae 9). J. Submicrosc. Cytol. Pathol. 1996, 28, 587–596. [Google Scholar]
- Sergerie, M.; Laforest, G.; Bujan, L.; Bissonnette, F.; Bleau, G. Sperm DNA fragmentation: Threshold value in male fertility. Hum. Reprod. 2005, 20, 3446–3451. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Maynou, J.; García-Peiró, A.; Fernández-Encinas, A.; Abad, C.; Amengual, M.J.; Prada, E.; Navarro, J.; Benet, J. Comprehensive analysis of sperm DNA fragmentation by five different assays: TUNEL assay, SCSA, SCD test and alkaline and neutral Comet assay. Andrology 2013, 1, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.; Aitken, J.R.; Conner, S.J.; Iuliis, G.D.; Evenson, D.P.; Henkel, R.; Giwercman, A.; Gharagozloo, P. The impact of sperm DNA damage in assisted conception and beyond: Recent advances in diagnosis and treatment. Reprod. Biomed. Online 2013, 27, 325–337. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| All Subjects | Patients | Fertile Men | p# | |
|---|---|---|---|---|
| Number of Subjects | 162 | 91 | 71 | |
| Age (y) | 38.00 (34.00–42.00) | 38.00 (35.00–42.00) | 36.00 (34.00–42.00) | 0.121 |
| Sperm Number (millions/ejaculate) | 175.75 (106.35–271.80) | 153.00 (67.84–266.60) | 185.49 (114.40–285.00) | 0.133 |
| Concentration (106/mL) | 52.50 (33.00–102.20) | 46.00 (22.00–88.00) | 62.00 (38.60–121.50) | 0.067 |
| Total Motility (%) | 60.00 (51.75–70.00) | 57.00 (47.00–64.00) | 65.00 (57.00–74.00) | <0.001 |
| Progressive Motility (%) | 50.00 (40.00-60.00) | 45.00 (33.00–54.00) | 54.00 (47.00–64.00) | <0.001 |
| Morphology (%) | 5.00 (3.00–9.00) | 5.00 (2.00–8.00) | 7.00 (4.00–9.00) | <0.001 |
| Abstinence (d) | 4.00 (3.00–4.25) | 4.00 (3.00–5.00) | 3.00 (3.00–4.00) | 0.012 |
| Semen Volume (ml) | 3.10 (2.48–4.60) | 3.20 (2.50–4.60) | 3.10 (2.30–4.50) | 0.125 |
| Variable | Total sDF | Viable sDF | Non−Viable sDF |
|---|---|---|---|
| r | r | r | |
| p | p | p | |
| Age, y | 0.189 * | 0.086 | 0.219 * |
| 0.016 | 0.278 | 0.005 | |
| Sperm Number (×106/ejaculate) | −0.061 | −0.088 | 0.022 |
| 0.444 | 0.264 | 0.785 | |
| Concentration (106/mL) | −0.03 | −0.034 | 0.064 |
| 0.705 | 0.67 | 0.418 | |
| Total Motility (%) | −0.277 * | −0.12 | −0.202 * |
| 0 | 0.128 | 0.01 | |
| Progressive Motility (%) | −0.282 * | −0.136 | −0.183 * |
| 0 | 0.084 | 0.02 | |
| Morphology (%) | −0.256 * | −0.101 | −0.159 * |
| 0.001 | 0.202 | 0.044 | |
| Abstinence, d | 0.203 * | 0.074 | 0.231 * |
| 0.01 | 0.352 | 0.003 |
| AUC (95% CI) | SE | p-Value | |
|---|---|---|---|
| Total sDF | 0.696 (0.615–0.776) | 0.041 | <0.001 |
| Viable sDF | 0.718 (0.640–0.797) | 0.040 | <0.001 |
| Non-viable sDF | 0.760 (0.685–0.835 | 0.038 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muratori, M.; Pellegrino, G.; Mangone, G.; Azzari, C.; Lotti, F.; Tarozzi, N.; Boni, L.; Borini, A.; Maggi, M.; Baldi, E. DNA Fragmentation in Viable and Non-Viable Spermatozoa Discriminates Fertile and Subfertile Subjects with Similar Accuracy. J. Clin. Med. 2020, 9, 1341. https://doi.org/10.3390/jcm9051341
Muratori M, Pellegrino G, Mangone G, Azzari C, Lotti F, Tarozzi N, Boni L, Borini A, Maggi M, Baldi E. DNA Fragmentation in Viable and Non-Viable Spermatozoa Discriminates Fertile and Subfertile Subjects with Similar Accuracy. Journal of Clinical Medicine. 2020; 9(5):1341. https://doi.org/10.3390/jcm9051341
Chicago/Turabian StyleMuratori, Monica, Giulia Pellegrino, Giusi Mangone, Chiara Azzari, Francesco Lotti, Nicoletta Tarozzi, Luca Boni, Andrea Borini, Mario Maggi, and Elisabetta Baldi. 2020. "DNA Fragmentation in Viable and Non-Viable Spermatozoa Discriminates Fertile and Subfertile Subjects with Similar Accuracy" Journal of Clinical Medicine 9, no. 5: 1341. https://doi.org/10.3390/jcm9051341
APA StyleMuratori, M., Pellegrino, G., Mangone, G., Azzari, C., Lotti, F., Tarozzi, N., Boni, L., Borini, A., Maggi, M., & Baldi, E. (2020). DNA Fragmentation in Viable and Non-Viable Spermatozoa Discriminates Fertile and Subfertile Subjects with Similar Accuracy. Journal of Clinical Medicine, 9(5), 1341. https://doi.org/10.3390/jcm9051341