Fibroblast Growth Factor Family in the Progression of Prostate Cancer



Abstract
1. Introduction
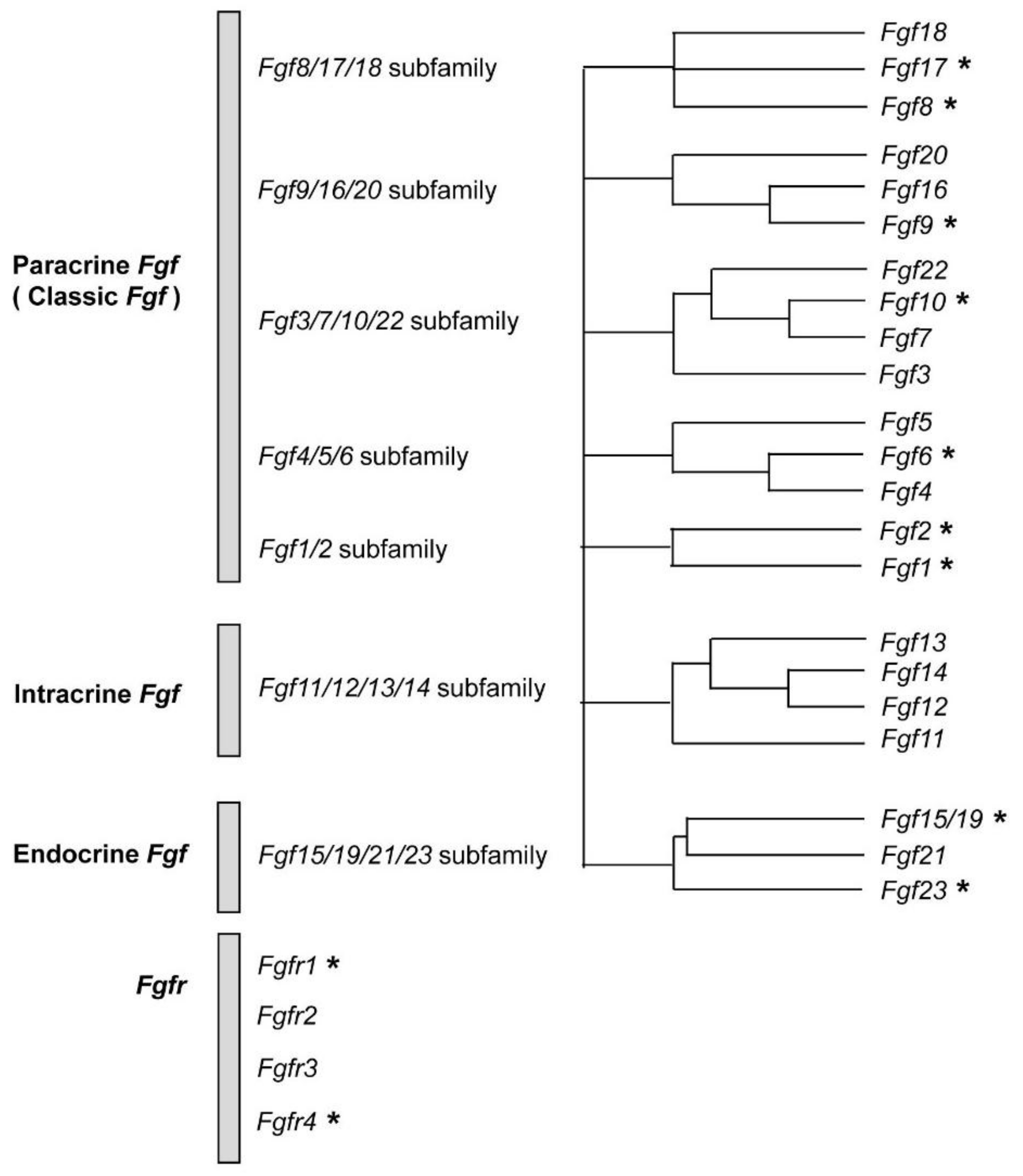
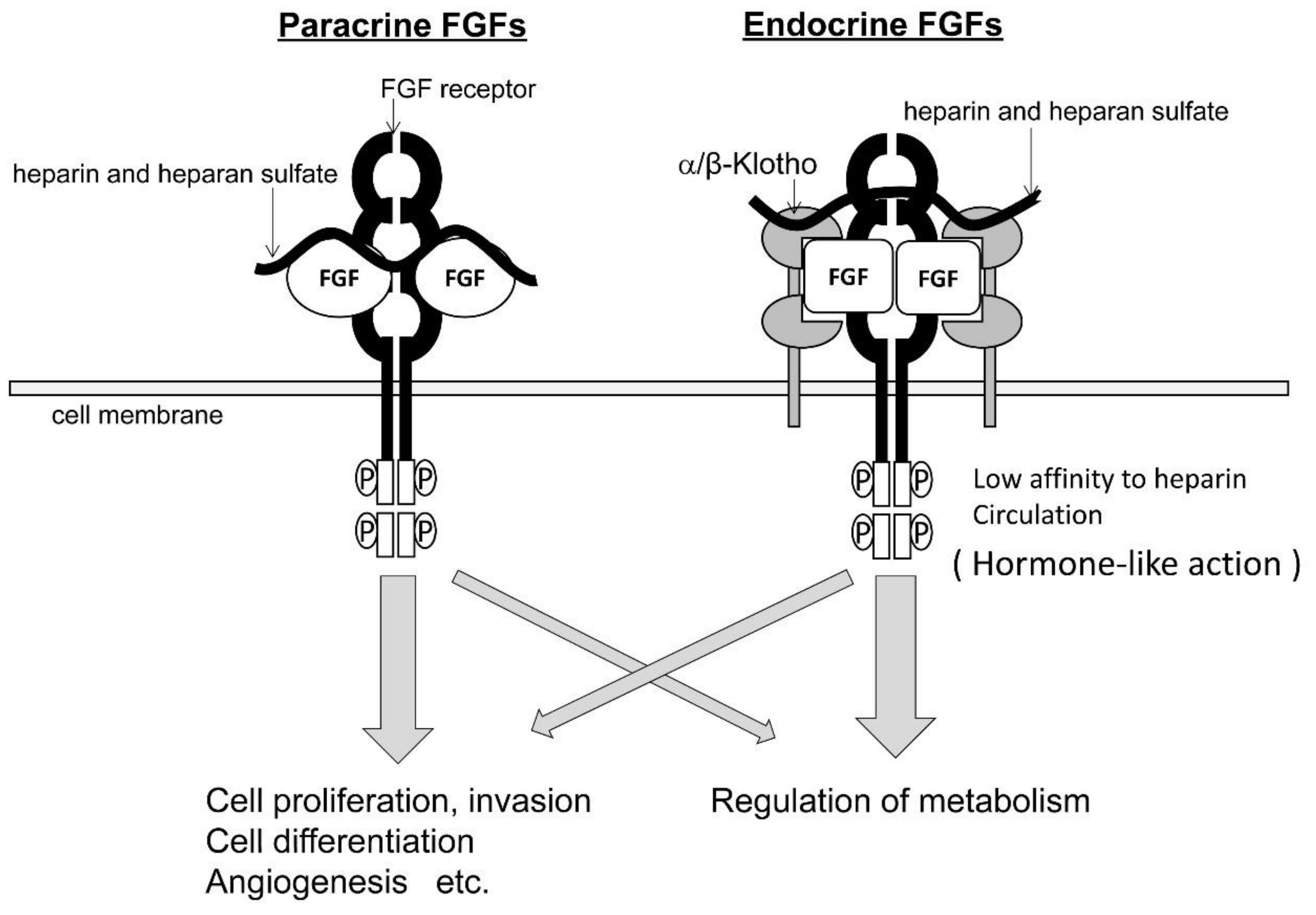
2. FGFs and FGFRs
3. FGF-FGFR Signaling in Epithelial-Stromal Interaction in Prostate Tissue
4. Effects of the Restoration of FGFR2IIIb in Prostate Cancer Cells
5. Involvement of FGFs in the Development and Progression of Prostate Cancer
5.1. Paracrine FGFs
5.2. Endocrine FGFs
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Corn, P.G.; Wang, F.; McKeehan, W.L.; Navone, N. Targeting fibroblast growth factor pathways in prostate cancer. Clin. Cancer. Res. 2013, 19, 5856–5866. [Google Scholar] [CrossRef] [PubMed]
- Wesche, J.; Haglund, K.; Haugsten, E.M. Fibroblast growth factors and their receptors in cancer. Biochem. J. 2011, 437, 199–213. [Google Scholar] [CrossRef]
- Itoh, N.; Ornitz, D.M. Fibroblast growth factors: From molecular evolution to roles in development, metabolism and disease. J. Biochem. 2011, 149, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Degirolamo, C.; Sabbà, C.; Moschetta, A. Therapeutic potential of the endocrine fibroblast growth factors FGF19, FGF21 and FGF23. Nat. Rev. Drug Discov. 2016, 15, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.C.; Wang, M.; Blackmore, C.; Desnoyers, L.R. Liver-specific activities of FGF19 require Klotho beta. J. Biol. Chem. 2007, 282, 27277–27284. [Google Scholar] [CrossRef] [PubMed]
- Kurosu, H.; Choi, M.; Ogawa, Y.; Dickson, A.S.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Rosenblatt, K.P.; Kliewer, S.A.; Kuro-o, M. Tissue-specific expression of betaKlotho and fibroblast growth factor (FGF) receptor isoforms determines metabolic activity of FGF19 and FGF21. J. Biol. Chem. 2007, 282, 26687–26695. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.H.; Holcomb, I.; Deuel, B.; Dowd, P.; Huang, A.; Vagts, A.; Foster, J.; Liang, J.; Brush, J.; Gu, Q.; et al. FGF-19, a novel fibroblast growth factor with unique specificity for FGFR4. Cytokine 1999, 11, 729–735. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kurosu, H.; Yamamoto, M.; Nandi, A.; Rosenblatt, K.P.; Goetz, R.; Eliseenkova, A.V.; Mohammadi, M.; Kuro-o, M. β-Klotho is required for metabolic activity of fibroblast growth factor 21. Proc. Natl. Acad. Sci. USA 2007, 104, 7432–7437. [Google Scholar] [CrossRef]
- Ding, X.; Boney-Montoya, J.; Owen, B.M.; Bookout, A.L.; Coate, K.C.; Mangelsdorf, D.J.; Kliewer, S.A. β-Klotho is required for fibroblast growth factor 21 effects on growth and metabolism. Cell Metab. 2012, 16, 387–393. [Google Scholar] [CrossRef]
- Kolek, O.I.; Hines, E.R.; Jones, M.D.; LeSueur, L.K.; Lipko, M.A.; Kiela, P.R.; Collins, J.F.; Haussler, M.R.; Ghishan, F.K. 1alpha, 25-Dihydroxyvitamin D3 upregulates FGF23 gene expression in bone: The final link in a renal-gastrointestinal-skeletal axis that controls phosphate transport. Am. J. Physiol. Gastrointest. Liver. Physiol. 2005, 289, G1036–G1042. [Google Scholar] [CrossRef]
- Yu, X.; Sabbagh, Y.; Davis, S.I.; Demay, M.B.; White, K.E. Genetic dissection of phosphate- and vitamin D-mediated regulation of circulating Fgf23 concentrations. Bone 2005, 36, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.K.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor specificity of the fibroblast growth factor family. The complete mammalian FGF family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Wang, F. FGF signaling in prostate development, tissue homoeostasis and tumorigenesis. Biosci. Rep. 2010, 30, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Cotton, L.M.; O’Bryan, M.K.; Hinton, B.T. Cellular signaling by fibroblast growth factors (FGFs) and their receptors (FGFRs) in male reproduction. Endocr. Rev. 2008, 29, 193–216. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hamana, T.; Liu, J.; Wang, C.; An, L.; You, P.; Chang, J.Y.; Xu, J.; Jin, C.; Zhang, Z.; et al. Type 2 Fibroblast Growth Factor Receptor Signaling Preserves Stemness and Prevents Differentiation of Prostate Stem Cells from the Basal Compartment. J. Biol. Chem. 2015, 290, 17753–17761. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liu, G.; Zhang, Y.; Hu, Y.P.; Yu, K.; Lin, C.; McKeehan, K.; Xuan, J.W.; Ornitz, D.M.; Shen, M.M.; et al. Fibroblast growth factor receptor 2 tyrosine kinase is required for prostatic morphogenesis and the acquisition of strict androgen dependency for adult tissue homeostasis. Development 2007, 134, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, A.; Yasumoto, H.; Usui, T. Hormone Refractory Prostate Cancer and Fibroblast Growth Factor Receptor. Breast Cancer 1999, 6, 320–324. [Google Scholar] [CrossRef]
- Carstens, R.P.; Eaton, J.V.; Krigman, H.R.; Walther, P.J.; Garcia-Blanco, M.A. Alternative splicing of fibroblast growth factor receptor 2 (FGF-R2) in human prostate cancer. Oncogene 1997, 15, 3059–3065. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Lin, Y.; Lan, Y.; Lin, C.; Xuan, J.W.; Shen, M.M.; McKeehan, W.L.; Greenberg, N.M.; Wang, F. Role of epithelial cell fibroblast growth factor receptor substrate 2alpha in prostate development, regeneration and tumorigenesis. Development 2008, 135, 775–784. [Google Scholar] [CrossRef]
- Liu, J.; You, P.; Chen, G.; Fu, X.; Zeng, X.; Wang, C.; Huang, Y.; An, L.; Wan, X.; Navone, N.; et al. Hyperactivated FRS2α-mediated signaling in prostate cancer cells promotes tumor angiogenesis and predicts poor clinical outcome of patients. Oncogene 2016, 35, 1750–1759. [Google Scholar] [CrossRef]
- Lu, W.; Luo, Y.; Kan, M.; McKeehan, W.L. Fibroblast growth factor-10. A second candidate stromal to epithelial cell andromedin in prostate. J. Biol. Chem. 1999, 274, 12827–12834. [Google Scholar] [CrossRef]
- Feng, S.; Wang, F.; Matsubara, A.; Kan, M.; McKeehan, W.L. Fibroblast growth factor receptor 2 limits and receptor 1 accelerates tumorigenicity of prostate epithelial cells. Cancer Res. 1997, 57, 5369–5378. [Google Scholar] [PubMed]
- Turner, N.; Grose, R. Fibroblast growth factor signaling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Ingram, L.; Kim, S.; Beharry, Z.; Cooper, J.A.; Cai, H. Paracrine Fibroblast Growth Factor Initiates Oncogenic Synergy with Epithelial FGFR/Src Transformation in Prostate Tumor Progression. Neoplasia 2018, 20, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.; Wang, J.; Karatas, O.F.; Feng, S.; Zhang, Y.; Creighton, C.J.; Ittmann, M. Fibroblast growth factor receptor signaling plays a key role in transformation induced by the TMPRSS2/ERG fusion gene and decreased PTEN. Oncotarget 2018, 9, 14456–14471. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Saribekyan, G.; Liao, C.P.; Cohen, M.B.; Roy-Burman, P. Cooperation between FGF8b overexpression and PTEN deficiency in prostate tumorigenesis. Cancer Res. 2006, 66, 2188–2194. [Google Scholar] [CrossRef]
- Ronca, R.; Tamma, R.; Coltrini, D.; Ruggieri, S.; Presta, M.; Ribatti, D. Fibroblast growth factor modulates mast cell recruitment in a murine model of prostate cancer. Oncotarget 2017, 8, 82583–82592. [Google Scholar] [CrossRef]
- Liu, J.; Chen, G.; Liu, Z.; Liu, S.; Cai, Z.; You, P.; Ke, Y.; Lai, L.; Huang, Y.; Gao, H.; et al. Aberrant FGFR tyrosine kinase signaling enhances the Warburg effect by reprogramming LDH isoform expression and activity in prostate cancer. Cancer Res. 2018, 78, 4459–4470. [Google Scholar] [CrossRef]
- Wang, C.; Ke, Y.; Liu, S.; Pan, S.; Liu, Z.; Zhang, H.; Fan, Z.; Zhou, C.; Liu, J.; Wang, F. Ectopic fibroblast growth factor receptor 1 promotes inflammation by promoting nuclear factor-κB signaling in prostate cancer cells. J. Biol. Chem. 2018, 293, 14839–14849. [Google Scholar] [CrossRef]
- Acevedo, V.D.; Gangula, R.D.; Freeman, K.W.; Li, R.; Zhang, Y.; Wang, F.; Ayala, G.E.; Peterson, L.E.; Ittmann, M.; Spencer, D.M. Inducible FGFR-1 activation leads to irreversible prostate adenocarcinoma and an epithelial-to-mesenchymal transition. Cancer Cell. 2007, 12, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Huang, Y.; Jin, C.; Luo, Y.; Okamoto, T.; Wang, F.; McKeehan, W.L. FGFR1 abrogates inhibitory effect of androgen receptor concurrent with induction of androgen-receptor variants in androgen receptor-negative prostate tumor epithelial cells. Prostate 2011, 71, 1691–1700. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, A.; Kan, M.; Feng, S.; McKeehan, W.L. Inhibition of growth of malignant rat prostate tumor cells by restoration of fibroblast growth factor receptor 2. Cancer Res 1998, 58, 1509–1514. [Google Scholar] [PubMed]
- Yasumoto, H.; Matsubara, A.; Mutaguchi, K.; Usui, T.; McKeehan, W.L. Restoration of fibroblast growth factor receptor2 suppresses growth and tumorigenicity of malignant human prostate carcinoma PC-3 cells. Prostate 2004, 61, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, A.; Teishima, J.; Mirkhat, S.; Yasumoto, H.; Mochizuki, H.; Seki, M.; Mutaguchi, K.; Mckeehan, W.L.; Usui, T. Restoration of FGF receptor type 2 enhances radiosensitivity of hormone-refractory human prostate carcinoma PC-3 cells. Anticancer Res. 2008, 28, 2141–2146. [Google Scholar] [PubMed]
- Shoji, K.; Teishima, J.; Hayashi, T.; Ohara, S.; Mckeehan, W.L.; Matsubara, A. Restoration of fibroblast growth factor receptor 2IIIb enhances the chemosensitivity of human prostate cancer cells. Oncol. Rep. 2014, 32, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ronca, R.; Giacomini, A.; Di Salle, E.; Coltrini, D.; Pagano, K.; Ragona, L.; Matarazzo, S.; Rezzola, S.; Maiolo, D.; Torrella, R.; et al. Long-Pentraxin 3 derivative as a small-molecule FGF trap for cancer therapy. Cancer Cell. 2015, 28, 225–239. [Google Scholar] [CrossRef]
- Memarzadeh, S.; Xin, L.; Mulholland, D.J.; Mansukhani, A.; Wu, H.; Teitell, M.A.; Witte, O.N. Enhanced paracrine FGF10 expression promotes formation of multifocal prostate adenocarcinoma and an increase in epithelial androgen receptor. Cancer Cell. 2007, 12, 572–585. [Google Scholar] [CrossRef]
- Pecqueux, C.; Arslan, A.; Heller, M.; Falkenstein, M.; Kaczorowski, A.; Tolstov, Y.; Sültmann, H.; Grüllich, C.; Herpel, E.; Duensing, A. FGF-2 is a driving force for chromosomal instability and a stromal factor associated with adverse clinico-pathological features in prostate cancer. Urol. Oncol. 2018, 36, e15–e365. [Google Scholar] [CrossRef]
- Cuevas, R.; Korzeniewski, N.; Tolstov, Y.; Hohenfellner, M.; Duensing, S. FGF-2 disrupts mitotic stability in prostate cancer through the intracellular trafficking protein CEP57. Cancer Res. 2013, 73, 1400–1410. [Google Scholar] [CrossRef]
- Meng, X.; Vander Ark, A.; Daft, P.; Woodford, E.; Wang, J.; Madaj, Z.; Li, X. Loss of TGF-β signaling in osteoblasts increases basic-FGF and promotes prostate cancer bone metastasis. Cancer Lett. 2018, 418, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Wang, F.; Wu, X.; Yu, C.; Luo, Y.; McKeehan, W.L. Directionally specific paracrine communication mediated by epithelial FGF9 to stromal FGFR3 in two-compartment premalignant prostate tumors. Cancer Res. 2004, 64, 4555–4562. [Google Scholar] [CrossRef] [PubMed]
- Reuss, B.; Hertel, M.; Werner, S.; Unsicker, K. Fibroblast growth factors-5 and -9 distinctly regulate expression and function of the gap junction protein connexin43 in cultured astroglial cells from different brain regions. Glia 2000, 30, 231–241. [Google Scholar] [CrossRef]
- Lum, M.; Turbic, A.; Mitrovic, B.; Turnley, A.M.J. Fibroblast growth factor-9 inhibits astrocyte differentiation of adult mouse neural progenitor cells. Neurosci. Res. 2009, 87, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Cohen, R.I.; Chandross, K.J. Fibroblast growth factor-9 modulates the expression of myelin related proteins and multiple fibroblast growth factor receptors in developing oligodendrocytes. J. Neurosci. Res. 2000, 61, 273–287. [Google Scholar] [CrossRef]
- Harada, M.; Murakami, H.; Okawa, A.; Okimoto, N.; Hiraoka, S.; Nakahara, T.; Akasaka, R.; Shiraishi, Y.; Futatsugi, N.; Mizutani-Koseki, Y.; et al. FGF9 monomer/ dimer equilibrium regulates extracellular matrix affinity and tissue diffusion. Nat. Genet. 2009, 41, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Kalinina, J.; Byron, S.A.; Makarenkova, H.P.; Olsen, S.K.; Eliseenkova, A.V.; Larochelle, W.J.; Dhanabal, M.; Blais, S.; Ornitz, D.M.; Day, L.A.; et al. Homodimerization controls the fibroblast growth factor 9 subfamily’s receptor binding and heparan sulfate-dependent diffusion in the extracellular matrix. Mol. Cell. Biol. 2009, 29, 4663–4678. [Google Scholar] [CrossRef]
- Miyagi, N.; Kato, S.; Terasaki, M.; Aoki, T.; Sugita, Y.; Yamaguchi, M.; Shigemori, M.; Morimatsu, M. Fibroblast growth factor-9 (glia-activating factor) stimulates proliferation and production of glial fibrillary acidic protein in human gliomas either in the presence or in the absence of the endogenous growth factor expression. Oncol. Rep. 1999, 6, 87–92. [Google Scholar] [CrossRef]
- Hendrix, N.D.; Wu, R.; Kuick, R.; Schwartz, D.R.; Fearon, E.R.; Cho, K.R. Fibroblast growth factor 9 has oncogenic activity and is a downstream target of Wnt signaling in ovarian endometrioid adenocarcinomas. Cancer Res. 2006, 66, 1354–1362. [Google Scholar] [CrossRef]
- Wang, C.K.; Chang, H.; Chen, P.H.; Chang, J.T.; Kuo, Y.C.; Ko, J.L.; Lin, P. Aryl hydrocarbon receptor activation and overexpression upregulated fibroblast growth factor-9 in human lung adenocarcinomas. Int. J. Cancer. 2009, 125, 807–815. [Google Scholar] [CrossRef]
- Huang, Y.; Jin, C.; Hamana, T.; Liu, J.; Wang, C.; An, L.; McKeehan, W.L.; Wang, F. Overexpression of FGF9 in prostate epithelial cells augments reactive stroma formation and promotes prostate cancer progression. Int. J. Biol. Sci. 2015, 11, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Teishima, J.; Yano, S.; Shoji, K.; Hayashi, T.; Goto, K.; Kitano, H.; Oka, K.; Nagamatsu, H.; Matsubara, A. Accumulation of FGF9 in prostate cancer correlates with epithelial-to-mesenchymal transition and induction of VEGF-A expression. Anticancer Res. 2014, 34, 695–700. [Google Scholar]
- Teishima, J.; Shoji, K.; Hayashi, T.; Miyamoto, K.; Ohara, S.; Matsubara, A. Relationship between the localization of fibroblast growth factor 9 in prostate cancer cells and postoperative recurrence. Prostate Cancer Prostatic Dis. 2012, 15, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Li, ZG.; Mathew, P.; Yang, J.; Starbuck, M.W.; Zurita, A.J.; Liu, J.; Sikes, C.; Multani, A.S.; Efstathiou, E.; Lopez, A.; et al. Androgen receptor-negative human prostate cancer cells induce osteogenesis in mice through FGF9-mediated mechanisms. J. Clin. Investig. 2008, 118, 2697–2710. [Google Scholar] [CrossRef] [PubMed]
- Bluemn, E.G.; Coleman, I.M.; Lucas, J.M.; Coleman, R.T.; Hernandez-Lopez, S.; Tharakan, R.; Bianchi-Frias, D.; Dumpit, R.F.; Kaipainen, A.; Corella, A.N.; et al. Androgen Receptor Pathway-Independent Prostate Cancer Is Sustained through FGF Signaling. Cancer Cell. 2017, 32, 474–489. [Google Scholar] [CrossRef] [PubMed]
- Nicholes, K.; Guillet, S.; Tomlinson, E.; Hillan, K.; Wright, B.; Frantz, G.D.; Pham, T.A.; Dillard-Telm, L.; Tsai, S.P.; Stephan, J.P.; et al. A mouse model of hepatocellular carcinoma: Ectopic expression of fibroblast growth factor 19 in skeletal muscle of transgenic mice. Am. J. Pathol. 2002, 160, 2295–2307. [Google Scholar] [CrossRef]
- Alvarez-Sola, G.; Uriarte, I.; Latasa, M.U.; Urtasun, R.; Bárcena-Varela, M.; Elizalde, M.; Jiménez, M.; Rodriguez-Ortigosa, C.M.; Corrales, F.J.; Fernández-Barrena, M.G.; et al. Fibroblast growth factor 15/19 in hepatocarcinogenesis. Dig. Dis. 2017, 35, 158–165. [Google Scholar] [CrossRef]
- Desnoyers, L.R.; Pai, R.; Ferrando, R.E.; Hötzel, K.; Le, T.; Ross, J.; Carano, R.; D’Souza, A.; Qing, J.; Mohtashemi, I.; et al. Targeting FGF19 inhibits tumor growth in colon cancer xenograft and FGF19 transgenic hepatocellular carcinoma models. Oncogene 2008, 27, 85–97. [Google Scholar] [CrossRef]
- Pai, R.; Dunlap, D.; Qing, J.; Mohtashemi, I.; Hotzel, K.; French, D.M. Inhibition of fibroblast growth factor 19 reduces tumor growth by modulating beta-catenin signaling. Cancer Res. 2008, 68, 5086–5095. [Google Scholar] [CrossRef]
- Nagamatsu, H.; Teishima, J.; Goto, K.; Shikuma, H.; Kitano, H.; Shoji, K.; Inoue, S.; Matsubara, A. FGF19 promotes progression of prostate cancer. Prostate 2015, 75, 1092–1101. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, H.; Ding, S.; Qi, S.; Liu, S.; Sun, D.; Dong, W.; Yin, L.; Li, M.; Zhao, X.; et al. βKlotho inhibits androgen/androgen receptor‑associated epithelial‑mesenchymal transition in prostate cancer through inactivation of ERK1/2 signaling. Oncol. Rep. 2018, 40, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Dakhova, O.; Creighton, C.J.; Ittmann, M. Endocrine fibroblast growth factor FGF19 promotes prostate cancer progression. Cancer Res. 2013, 73, 2551–2562. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wang, J.; Zhang, Y.; Creighton, C.J.; Ittmann, M. FGF23 promotes prostate cancer progression. Oncotarget 2015, 6, 17291–17301. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Up-Regulation | Down-Regulation | Increase | Decrease | |
|---|---|---|---|---|
| FGF19 | ||||
| Glycogen synthesis | Bile acid synthesis | Extrahepatic cholestasis | IBD | |
| Gluconeogenesis | Chronic hemodialysis | NAFLD | ||
| Primary bile acid malabsorption | ||||
| Obesity | ||||
| FGF21 | ||||
| Hepatic fatty acid oxidation | Ovulation | Type 2 diabetes | Anorexia | |
| Ketogenesis | Growth hormone signaling | Metabolic syndrome | Nervosa | |
| Glucogenesis | NAFLD | |||
| Thermogenesis | Coronary heart disease | |||
| WAT browing | ||||
| Growth hormone resistance | ||||
| Weight loss | ||||
| Ovulation | ||||
| FGF23 | ||||
| Calcium secretion | Renal phosphate absorption | ADHR | Hemodialysis | |
| Life span | Bone and renal calcium reabsorption | XLH rickets | Familial tumoral calcinosis | |
| Vitamin D synthesis | TIO | |||
| PTH secretion | Cardiac hypertrophy |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teishima, J.; Hayashi, T.; Nagamatsu, H.; Shoji, K.; Shikuma, H.; Yamanaka, R.; Sekino, Y.; Goto, K.; Inoue, S.; Matsubara, A. Fibroblast Growth Factor Family in the Progression of Prostate Cancer. J. Clin. Med. 2019, 8, 183. https://doi.org/10.3390/jcm8020183
Teishima J, Hayashi T, Nagamatsu H, Shoji K, Shikuma H, Yamanaka R, Sekino Y, Goto K, Inoue S, Matsubara A. Fibroblast Growth Factor Family in the Progression of Prostate Cancer. Journal of Clinical Medicine. 2019; 8(2):183. https://doi.org/10.3390/jcm8020183
Chicago/Turabian StyleTeishima, Jun, Tetsutaro Hayashi, Hirotaka Nagamatsu, Koichi Shoji, Hiroyuki Shikuma, Ryoken Yamanaka, Yohei Sekino, Keisuke Goto, Shogo Inoue, and Akio Matsubara. 2019. "Fibroblast Growth Factor Family in the Progression of Prostate Cancer" Journal of Clinical Medicine 8, no. 2: 183. https://doi.org/10.3390/jcm8020183
APA StyleTeishima, J., Hayashi, T., Nagamatsu, H., Shoji, K., Shikuma, H., Yamanaka, R., Sekino, Y., Goto, K., Inoue, S., & Matsubara, A. (2019). Fibroblast Growth Factor Family in the Progression of Prostate Cancer. Journal of Clinical Medicine, 8(2), 183. https://doi.org/10.3390/jcm8020183