Bevacizumab Induces Upregulation of Keratin 3 and VEGFA in Human Limbal Epithelial Cells in Vitro


Abstract
1. Introduction
2. Materials and Methods
2.1. Culturing of 3T3 Mouse Fibroblasts
2.2. Primary Human Limbal Epithelial Cell Harvesting and Maintenance
2.3. Treatment with Bevacizumab
2.4. Immunocytofluorescence
2.5. Colony-Forming Efficiency Assay
2.6. Cell Metabolic Activity
2.7. Scratch Wound Assay
2.8. RT-PCR
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Bevacizumab Does Not Affect Limbal Epithelial Cell Proliferation While It Decelerates Scratch Wound Healing
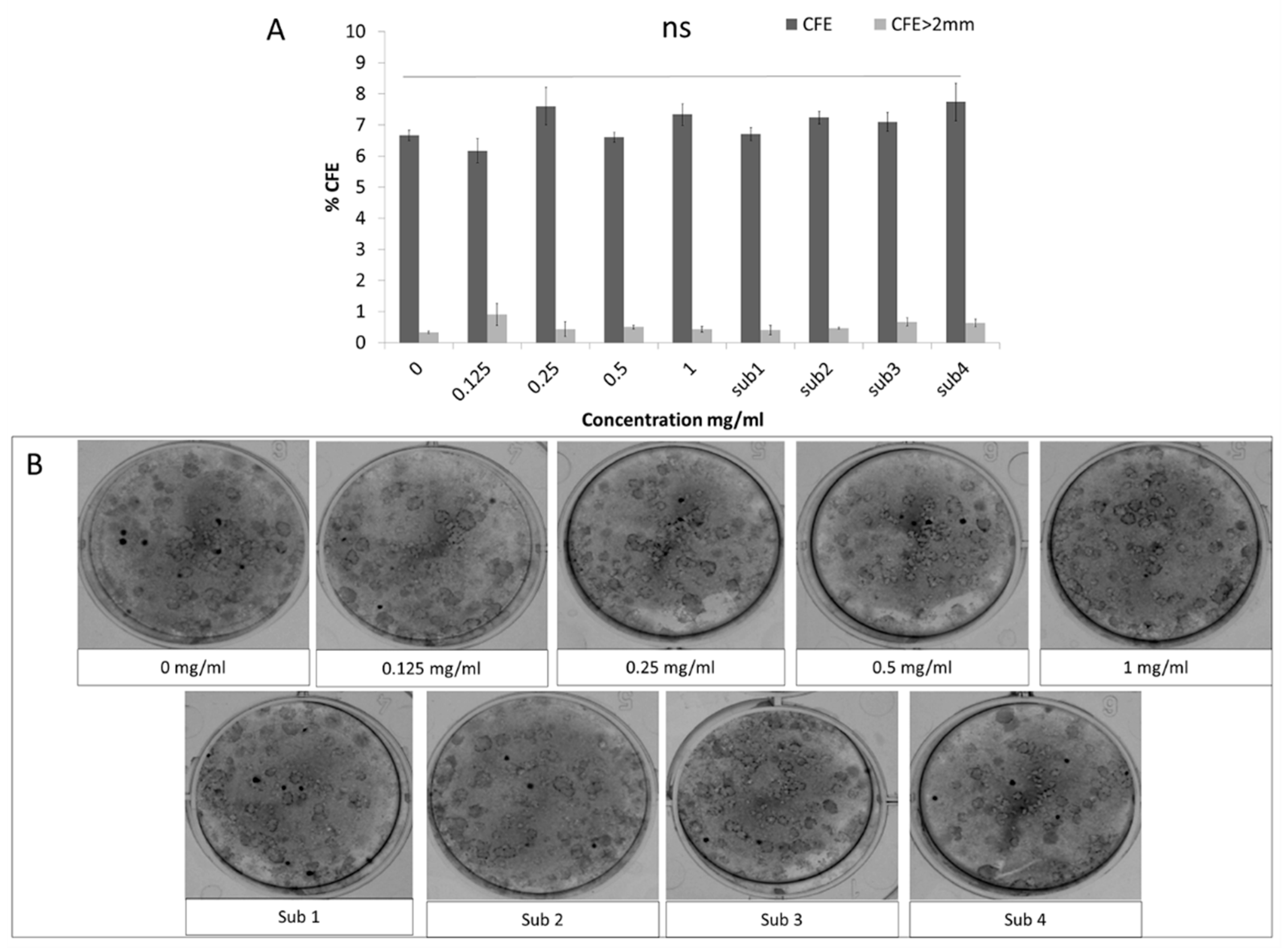
3.2. Colony Forming Efficiency (CFE) and Putative Limbal Epithelial Stem Cell Marker Expression Remains Unaffected by Bevacizumab While the Differentiation Marker Keratin 3 is Upregulated
3.3. VEGFA Expression of Limbal Epithelial Cells Is Upregulated in Response to Treatment with Bevacizumab
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bock, F.; Maruyama, K.; Regenfuss, B.; Hos, D.; Steven, P.; Heindl, L.M.; Cursiefen, C. Novel anti(lymph)angiogenic treatment strategies for corneal and ocular surface diseases. Prog. Retin. Eye Res. 2013, 34, 89–124. [Google Scholar] [CrossRef] [PubMed]
- Cursiefen, C.; Chen, L.; Saint-Geniez, M.; Hamrah, P.; Jin, Y.; Rashid, S.; Pytowski, B.; Persaud, K.; Wu, Y.; Streilein, J.W.; et al. Nonvascular VEGF receptor 3 expression by corneal epithelium maintains avascularity and vision. Proc. Natl. Acad. Sci. USA 2006, 103, 11405–11410. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Alatza, A.; Gilfillan, J.; Harris, A.R.; Levis, H.J.; Schrader, S.; Vernon, A.; Daniels, J.T. In sickness and in health: Corneal epithelial stem cell biology, pathology and therapy. Exp. Eye Res. 2010, 90, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Daniels, J.T. Biological principals and clinical potentials of limbal epithelial stem cells. Cell Tissue Res. 2008, 331, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Dua, H.S.; Joseph, A.; Shanmuganathan, V.A.; Jones, R.E. Stem cell differentiation and the effects of deficiency. Eye 2003, 17, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Sacchetti, M.; Lambiase, A.; Cortes, M.; Sgrulletta, R.; Bonini, S.; Merlo, D.; Bonini, S. Clinical and cytological findings in limbal stem cell deficiency. Graefe’s Arch. Clin. Exp. Ophthalmol./Albrecht von Graefes Archiv fur Klinische und Experimentelle Ophthalmologie 2005, 243, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Martin, R. Corneal conjunctivalisation in long-standing contact lens wearers. Clin. Exp. Optom. 2007, 90, 26–30. [Google Scholar] [CrossRef]
- Shortt, A.J.; Bunce, C.; Levis, H.J.; Blows, P.; Dore, C.J.; Vernon, A.; Secker, G.A.; Tuft, S.J.; Daniels, J.T. Three-year outcomes of cultured limbal epithelial allografts in aniridia and Stevens-Johnson syndrome evaluated using the Clinical Outcome Assessment in Surgical Trials assessment tool. Stem Cells Transl. Med. 2014, 3, 265–275. [Google Scholar] [CrossRef]
- Koch, S.; Tugues, S.; Li, X.; Gualandi, L.; Claesson-Welsh, L. Signal transduction by vascular endothelial growth factor receptors. Biochem. J. 2011, 437, 169–183. [Google Scholar] [CrossRef]
- Cursiefen, C.; Chen, L.; Borges, L.P.; Jackson, D.; Cao, J.; Radziejewski, C.; D’Amore, P.A.; Dana, M.R.; Wiegand, S.J.; Streilein, J.W. VEGF-A stimulates lymphangiogenesis and hemangiogenesis in inflammatory neovascularization via macrophage recruitment. J. Clin. Investig. 2004, 113, 1040–1050. [Google Scholar] [CrossRef]
- Ambati, B.K.; Nozaki, M.; Singh, N.; Takeda, A.; Jani, P.D.; Suthar, T.; Albuquerque, R.J.; Richter, E.; Sakurai, E.; Newcomb, M.T.; et al. Corneal avascularity is due to soluble VEGF receptor-1. Nature 2006, 443, 993–997. [Google Scholar] [CrossRef] [PubMed]
- Pavlakovic, H.; Becker, J.; Albuquerque, R.; Wilting, J.; Ambati, J. Soluble VEGFR-2: An antilymphangiogenic variant of VEGF receptors. Ann. N. Y. Acad. Sci. 2010, 1207 (Suppl. 1), E7–E15. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.H.; Gabison, E.E.; Kato, T.; Azar, D.T. Corneal neovascularization. Curr. Opin. Ophthalmol. 2001, 12, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Koenig, Y.; Bock, F.; Horn, F.; Kruse, F.; Straub, K.; Cursiefen, C. Short- and long-term safety profile and efficacy of topical Bevacizumab (Avastin) eye drops against corneal neovascularization. Graefe’s Arch. Clin. Exp. Ophthalmol./Albrecht von Graefes Archiv fur Klinische und Experimentelle Ophthalmologie 2009, 247, 1375–1382. [Google Scholar] [CrossRef]
- Koenig, Y.; Bock, F.; Kruse, F.E.; Stock, K.; Cursiefen, C. Angioregressive pretreatment of mature corneal blood vessels before keratoplasty: Fine-needle vessel coagulation combined with anti-VEGFs. Cornea 2012, 31, 887–892. [Google Scholar] [CrossRef]
- Ferrara, N.; Hillan, K.J.; Novotny, W. Bevacizumab (Avastin), a humanized anti-VEGF monoclonal antibody for cancer therapy. Biochem. Biophys. Res. Commun. 2005, 333, 328–335. [Google Scholar] [CrossRef]
- Grisanti, S.; Ziemssen, F. Bevacizumab: Off-label use in ophthalmology. Indian J. Ophthalmol. 2007, 55, 417–420. [Google Scholar] [CrossRef]
- Chalam, K.V.; Agarwal, S.; Brar, V.S.; Murthy, R.K.; Sharma, R.K. Evaluation of cytotoxic effects of Bevacizumab on human corneal cells. Cornea 2009, 28, 328–333. [Google Scholar] [CrossRef]
- Lin, C.T.; Hu, F.R.; Kuo, K.T.; Chen, Y.M.; Chu, H.S.; Lin, Y.H.; Chen, W.L. The different effects of early and late Bevacizumab (Avastin) injection on inhibiting corneal neovascularization and conjunctivalization in rabbit limbal insufficiency. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6277–6285. [Google Scholar] [CrossRef]
- Krizova, D.; Vokrojova, M.; Liehneova, K.; Studeny, P. Treatment of Corneal Neovascularization Using Anti-VEGF Bevacizumab. J. Ophthalmol. 2014, 2014, 178132. [Google Scholar] [CrossRef]
- Chang, J.H.; Garg, N.K.; Lunde, E.; Han, K.Y.; Jain, S.; Azar, D.T. Corneal neovascularization: An anti-VEGF therapy review. Surv. Ophthalmol. 2012, 57, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Cavallini, G.M.; Pellegrini, G.; Volante, V.; Ducange, P.; De Maria, M.; Torlai, G.; Benatti, C.; Forlini, M. Chemical injury treated with autologous limbal epithelial stem cell transplantation and subconjunctival Bevacizumab. Clin. Ophthalmol. 2014, 8, 1671–1673. [Google Scholar] [CrossRef] [PubMed]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal stem-cell therapy and long-term corneal regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, G.; Lambiase, A.; Macaluso, C.; Pocobelli, A.; Deng, S.; Cavallini, G.M.; Esteki, R.; Rama, P. From discovery to approval of an advanced therapy medicinal product-containing stem cells, in the EU. Regener. Med. 2016, 11, 407–420. [Google Scholar] [CrossRef]
- Luthra, S.; Sharma, A.; Dong, J.; Neekhra, A.; Gramajo, A.L.; Seigel, G.M.; Kenney, M.C.; Kuppermann, B.D. Effect of Bevacizumab (Avastinâ„¢) on mitochondrial function of in vitro retinal pigment epithelial, neurosensory retinal and microvascular endothelial cells. Indian J. Ophthalmol. 2013, 61, 705–710. [Google Scholar] [CrossRef]
- Malik, D.; Tarek, M.; Caceres del Carpio, J.; Ramirez, C.; Boyer, D.; Kenney, M.C.; Kuppermann, B.D. Safety profiles of anti-VEGF drugs: Bevacizumab, ranibizumab, aflibercept and ziv-aflibercept on human retinal pigment epithelium cells in culture. Br. J. Ophthalmol. 2014, 98, i11–i16. [Google Scholar] [CrossRef]
- Li, D.Q.; Chen, Z.; Song, X.J.; de Paiva, C.S.; Kim, H.S.; Pflugfelder, S.C. Partial enrichment of a population of human limbal epithelial cells with putative stem cell properties based on collagen type IV adhesiveness. Exp. Eye Res. 2005, 80, 581–590. [Google Scholar] [CrossRef]
- Di Iorio, E.; Barbaro, V.; Ruzza, A.; Ponzin, D.; Pellegrini, G.; De Luca, M. Isoforms of DeltaNp63 and the migration of ocular limbal cells in human corneal regeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 9523–9528. [Google Scholar] [CrossRef]
- Schermer, A.; Galvin, S.; Sun, T.T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef]
- Notara, M.; Shortt, A.J.; Galatowicz, G.; Calder, V.; Daniels, J.T. IL6 and the human limbal stem cell niche: A mediator of epithelial-stromal interaction. Stem Cell Res. 2010, 5, 188–200. [Google Scholar] [CrossRef]
- Notara, M.; Shortt, A.J.; O’Callaghan, A.R.; Daniels, J.T. The impact of age on the physical and cellular properties of the human limbal stem cell niche. Age 2013, 35, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Bock, F.; Onderka, J.; Rummelt, C.; Dietrich, T.; Bachmann, B.; Kruse, F.E.; Schlotzer-Schrehardt, U.; Cursiefen, C. Safety profile of topical VEGF neutralization at the cornea. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2095–2102. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Luthra, S.; Narayanan, R.; Marques, L.E.; Chwa, M.; Kim, D.W.; Dong, J.; Seigel, G.M.; Neekhra, A.; Gramajo, A.L.; Brown, D.J.; et al. Evaluation of in vitro effects of Bevacizumab (Avastin) on retinal pigment epithelial, neurosensory retinal, and microvascular endothelial cells. Retina 2006, 26, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, M.S.; Yoeruek, E.; Sierra, A.; Wallenfels-Thilo, B.; Schraermeyer, U.; Spitzer, B.; Bartz-Schmidt, K.U.; Szurman, P. Comparative antiproliferative and cytotoxic profile of Bevacizumab (Avastin), pegaptanib (Macugen) and ranibizumab (Lucentis) on different ocular cells. Graefe’s Arch. Clin. Exp. Ophthalmol./Albrecht von Graefes Archiv fur Klinische und Experimentelle Ophthalmologie 2007, 245, 1837–1842. [Google Scholar] [CrossRef]
- Rusovici, R.; Sakhalkar, M.; Chalam, K.V. Evaluation of cytotoxicity of Bevacizumab on VEGF-enriched corneal endothelial cells. Mol. Vis. 2011, 17, 3339–3346. [Google Scholar]
- Kim, E.K.; Kang, S.W.; Kim, J.Y.; Min, K.; Kim, T.I. Modulation of Bevacizumab-induced toxicity for cultured human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3922–3931. [Google Scholar] [CrossRef]
- Colombres, G.A.; Gramajo, A.L.; Arrambide, M.P.; Juarez, S.M.; Arevalo, J.F.; Bar, J.; Juarez, C.P.; Luna, J.D. Delayed Corneal Epithelial Healing after Intravitreal Bevacizumab: A Clinical and Experimental Study. J. Ophthalmic Vis. Res. 2011, 6, 18–25. [Google Scholar]
- Kim, E.C.; Lee, W.S.; Kim, M.S. The inhibitory effects of Bevacizumab eye drops on NGF expression and corneal wound healing in rats. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4569–4573. [Google Scholar] [CrossRef]
- Kim, E.C.; Ryu, H.W.; Lee, H.J.; Kim, M.S. Bevacizumab eye drops delay corneal epithelial wound healing and increase the stromal response to epithelial injury in rats. Clin. Exp. Ophthalmol. 2013, 41, 694–701. [Google Scholar] [CrossRef]
- Kim, S.W.; Ha, B.J.; Kim, E.K.; Tchah, H.; Kim, T.I. The effect of topical Bevacizumab on corneal neovascularization. Ophthalmology 2008, 115, e33–e38. [Google Scholar] [CrossRef]
- Kim, T.I.; Chung, J.L.; Hong, J.P.; Min, K.; Seo, K.Y.; Kim, E.K. Bevacizumab application delays epithelial healing in rabbit cornea. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4653–4659. [Google Scholar] [CrossRef] [PubMed]
- van Setten, G.B. Vascular endothelial growth factor (VEGF) in normal human corneal epithelium: Detection and physiological importance. Acta Ophthalmol. Scand. 1997, 75, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Thanos, M.; Pauklin, M.; Steuhl, K.P.; Meller, D. Ocular surface reconstruction with cultivated limbal epithelium in a patient with unilateral stem cell deficiency caused by Epidermolysis bullosa dystrophica hallopeau-Siemens. Cornea 2010, 29, 462–464. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense Primer | Antisense Primer |
|---|---|---|
| Keratin 3 | GGCAGAGATCGAGGGTGTC | GTCATCCTTCGCCTGCTGTAG |
| ΔNP63α | GGAAAACAATGCCCAGACTC(ΔN) | ATGATGAACAGCCCAACCTC(α-termini) |
| Integrin β1 | AGTGAATGGGAACAACGAGGTC | CAATTCCAGCAACCACACCA |
| VEGFA | ACAGGTACAGGGATGAGGACAC | AAGCAGGTGAGAGTAAGCGAAG |
| VEGFC | GCCTGTGAATGTACAGAAAGTCC | AATATGAAGGGACACAACGACAC |
| VEGFD | CCGCCATCCATACTCAATTATC | CCATAGCATGTCAATAGGACAGAG |
| VEGFR1 | CTACCACTCCCTTGAACACGA | GGTCCACTCCTTACACGACAA |
| VEGFR2 | ACCTCACCTGTTTCCTGTATGG | GACTGATTCCTGCTGTGTTGTC |
| VEGFR3 | CTCAAAGTCTCTCACGAACACG | GGTACATGCCAACGACACAG |
| TBP (housekeeping) | GTTGGTGGGTGAGCACAAG | AGGAGCCAAGAGTGAAGAACAG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Notara, M.; Lentzsch, A.; Clahsen, T.; Behboudifard, S.; Braun, G.; Cursiefen, C. Bevacizumab Induces Upregulation of Keratin 3 and VEGFA in Human Limbal Epithelial Cells in Vitro. J. Clin. Med. 2019, 8, 1925. https://doi.org/10.3390/jcm8111925
Notara M, Lentzsch A, Clahsen T, Behboudifard S, Braun G, Cursiefen C. Bevacizumab Induces Upregulation of Keratin 3 and VEGFA in Human Limbal Epithelial Cells in Vitro. Journal of Clinical Medicine. 2019; 8(11):1925. https://doi.org/10.3390/jcm8111925
Chicago/Turabian StyleNotara, Maria, Anna Lentzsch, Thomas Clahsen, Sara Behboudifard, Gabriele Braun, and Claus Cursiefen. 2019. "Bevacizumab Induces Upregulation of Keratin 3 and VEGFA in Human Limbal Epithelial Cells in Vitro" Journal of Clinical Medicine 8, no. 11: 1925. https://doi.org/10.3390/jcm8111925
APA StyleNotara, M., Lentzsch, A., Clahsen, T., Behboudifard, S., Braun, G., & Cursiefen, C. (2019). Bevacizumab Induces Upregulation of Keratin 3 and VEGFA in Human Limbal Epithelial Cells in Vitro. Journal of Clinical Medicine, 8(11), 1925. https://doi.org/10.3390/jcm8111925