
Role of Neuropilin-1 in Diabetic Nephropathy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
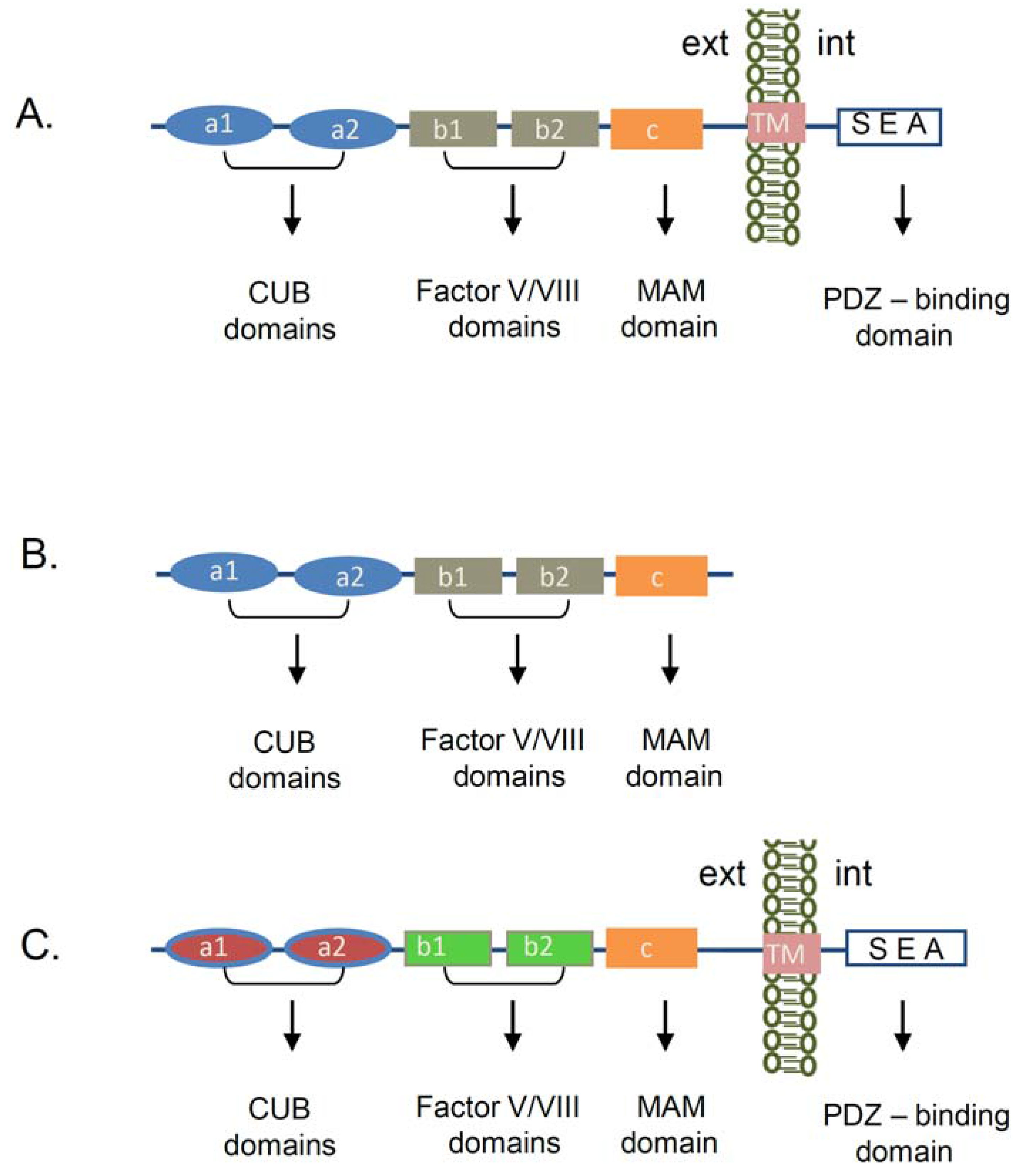
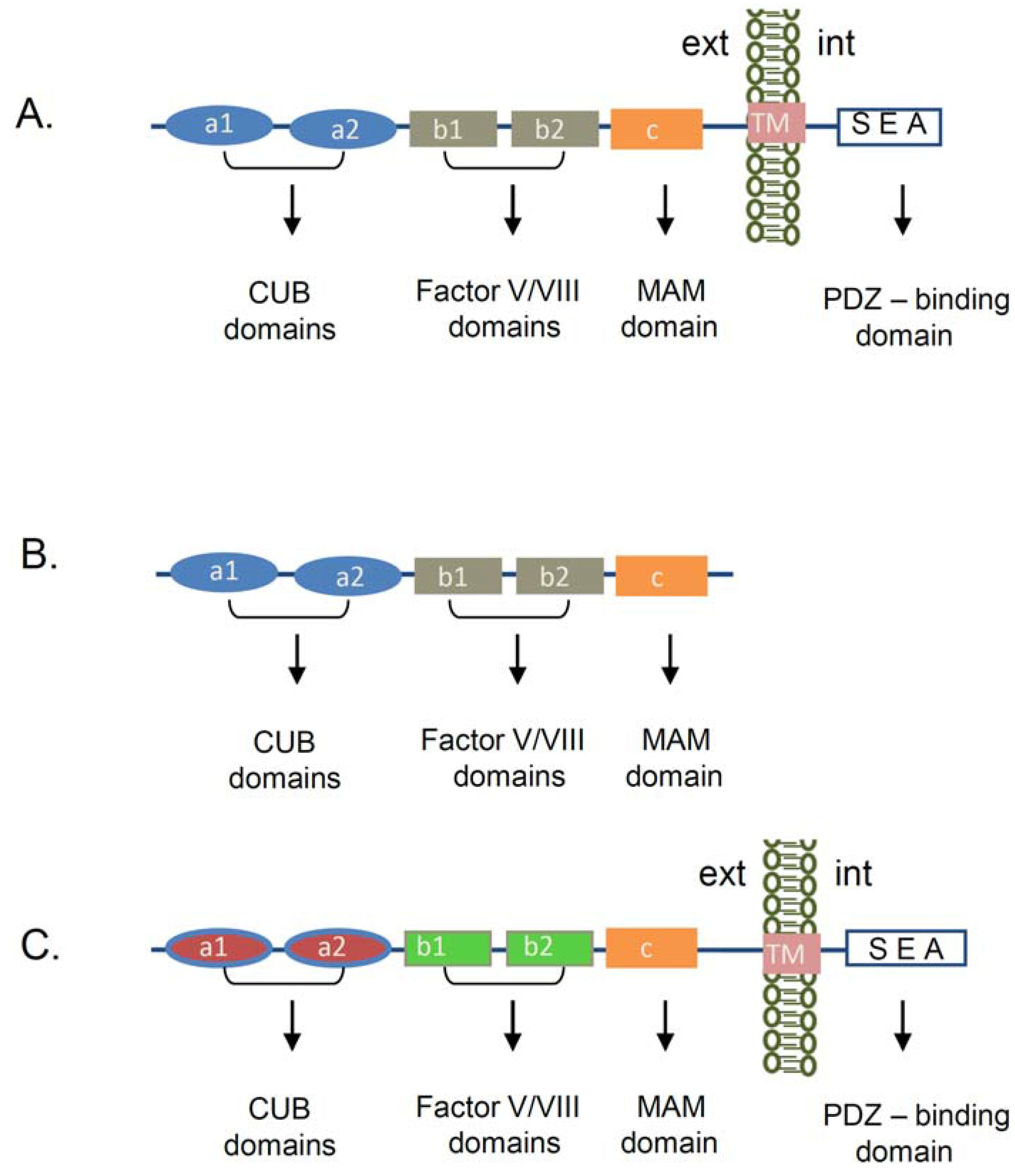
2. Neuropilin-1 Structure
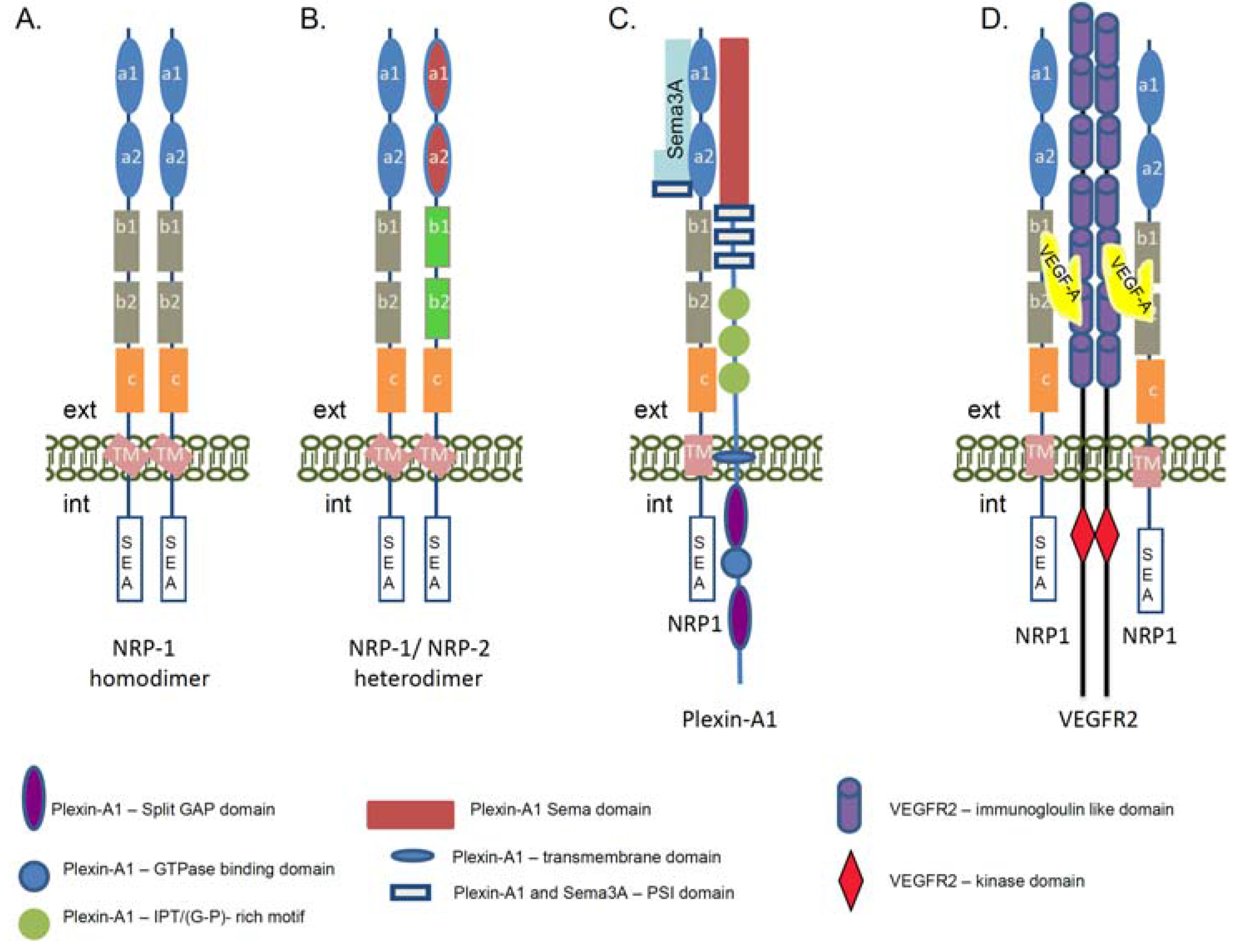
3. Neuropilin-1 Ligands and Signaling Co-Receptors
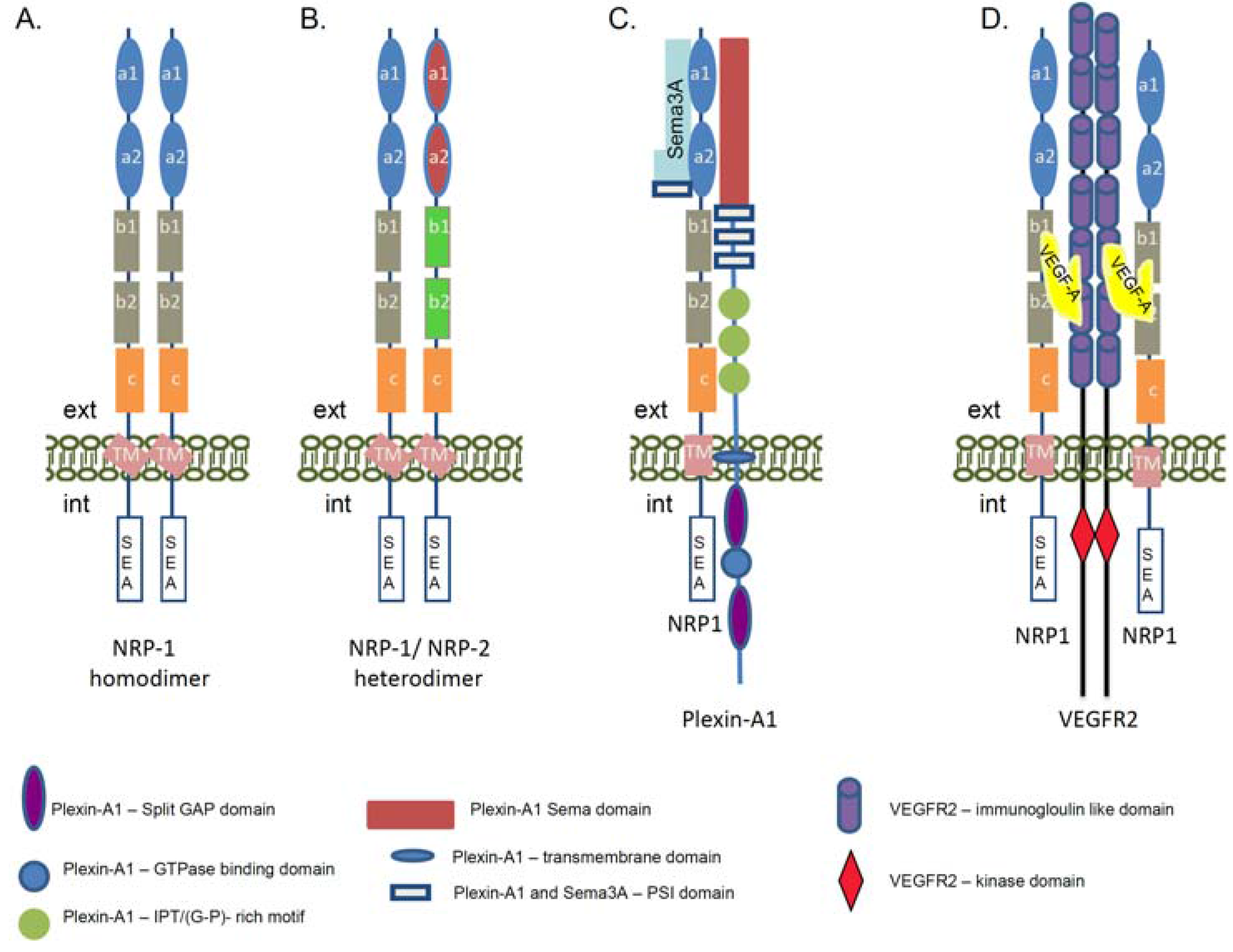
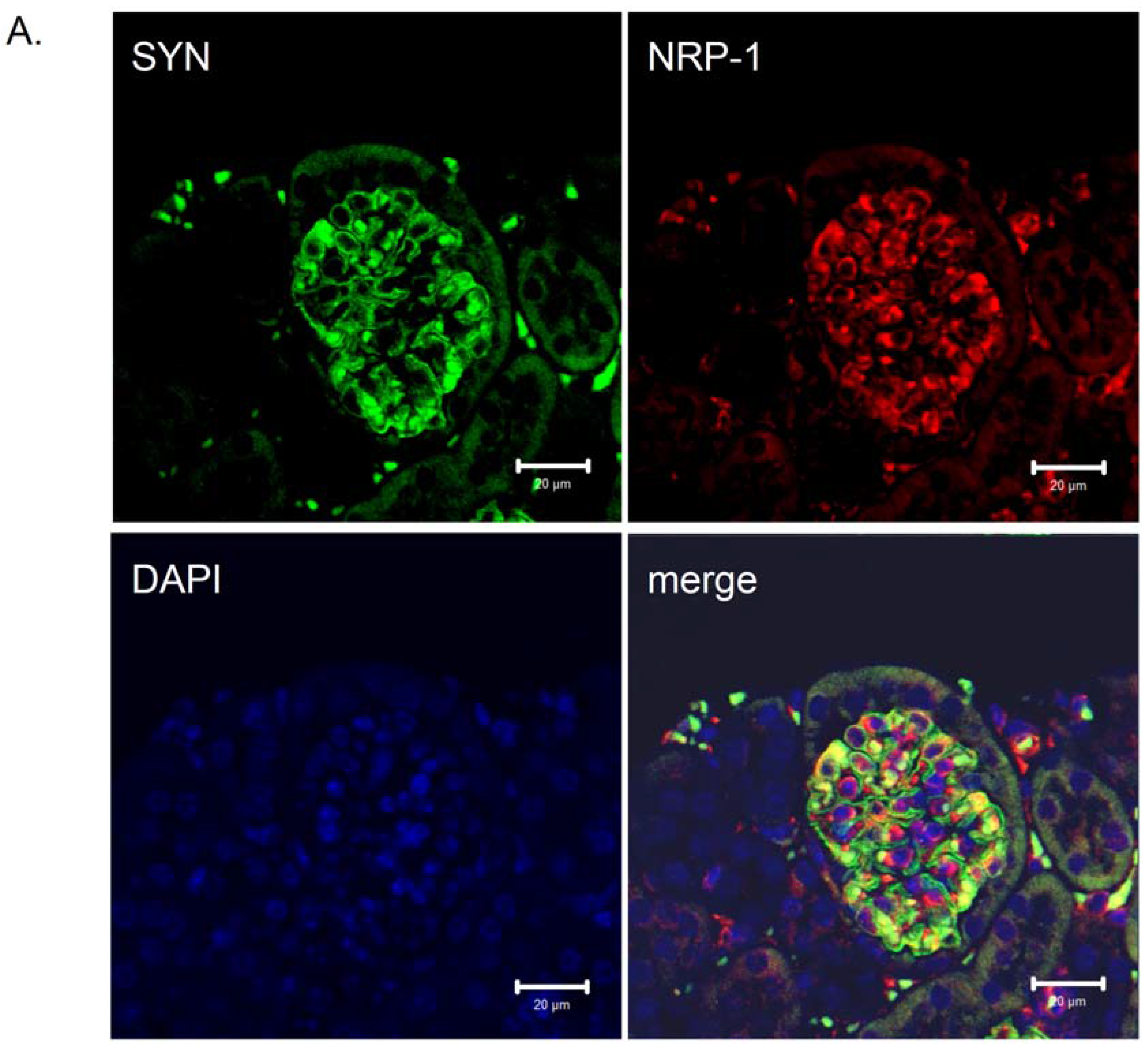
4. NRP-1 Expression in Diabetes and Diabetic Nephropathy
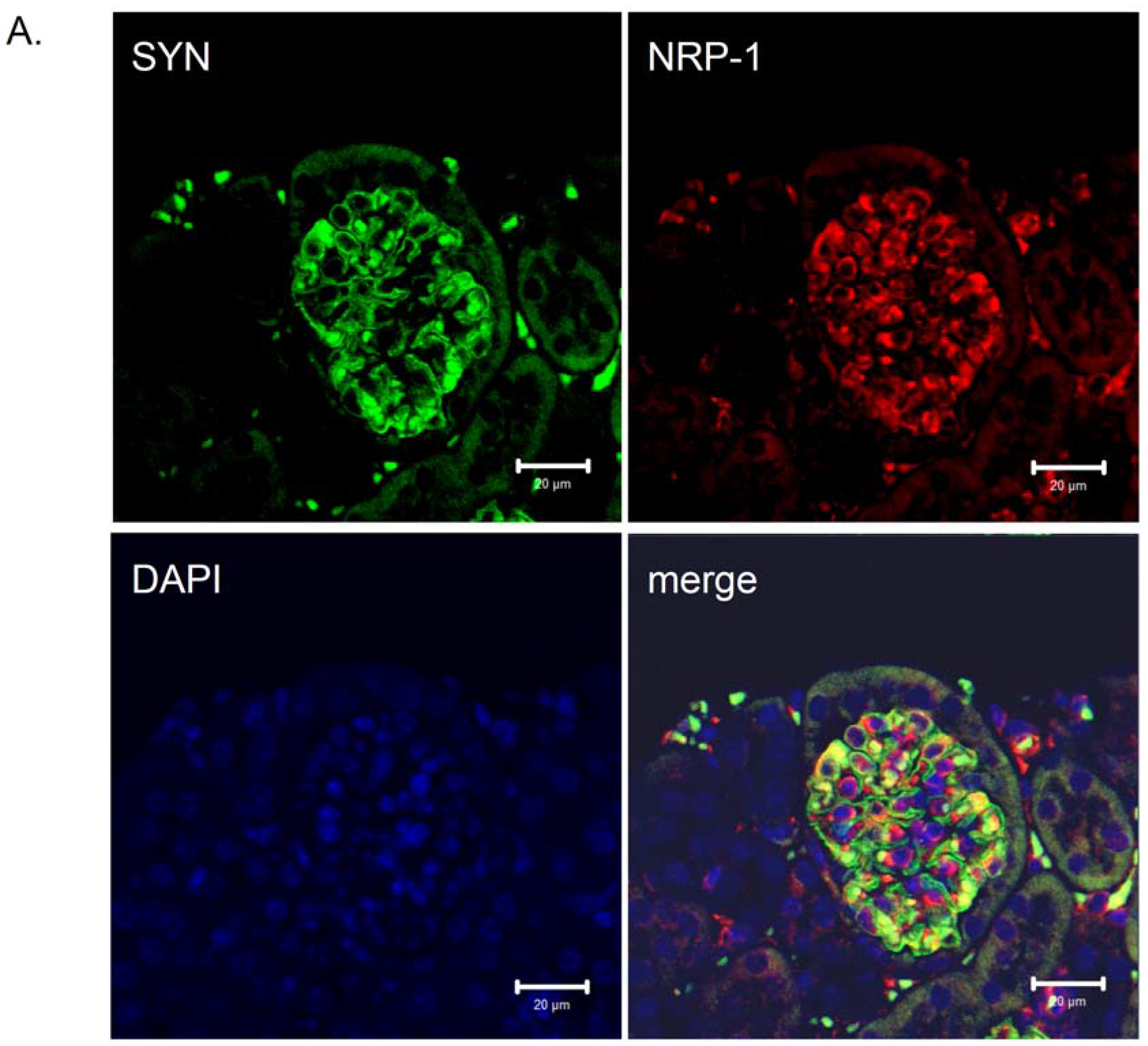
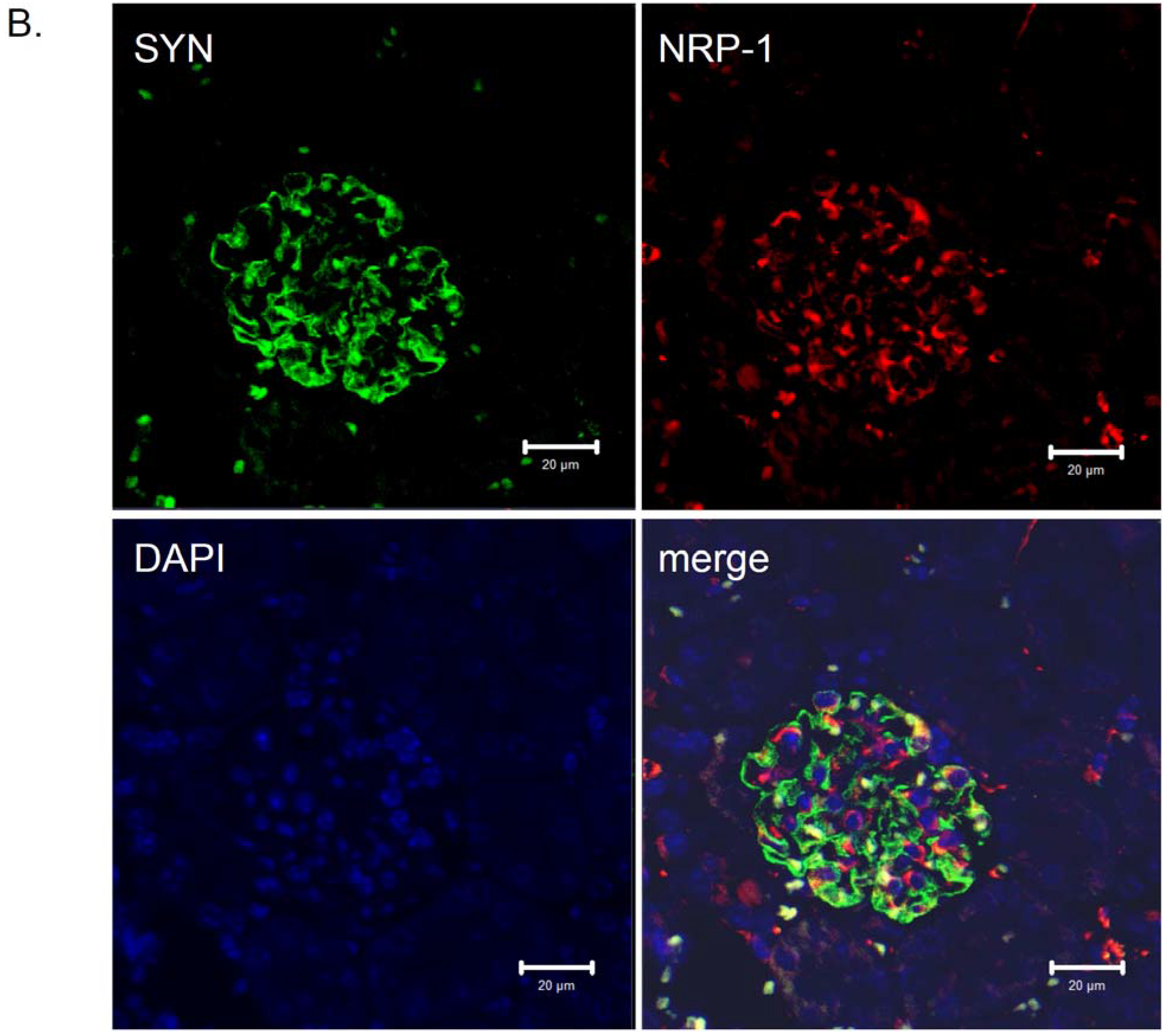
5. NRP-1 Is Implicated in Podocyte Adhesion and/or Migration
6. NRP-1 Ligand Sema3A and Its Function in DN
7. NRP-1 Ligand VEGF-A and Its Function in DN
8. Linking Neuropilin-1 to the Slit Diaphragm Proteins via CD2AP/NRP-1 Complex?
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wolf, G.; Ziyadeh, F.N. Cellular and molecular mechanisms of proteinuria in diabetic nephropathy. Nephron Physiol. 2007, 106, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Arora, M.K.; Singh, U.K. Molecular mechanisms in the pathogenesis of diabetic nephropathy: An update. Vasc. Pharmacol. 2013, 58, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Raptis, A.E.; Viberti, G. Pathogenesis of diabetic nephropathy. Exp. Clin. Endocrinol. Diabetes 2001, 109 (Suppl. S2), 424–437. [Google Scholar] [CrossRef] [PubMed]
- Bohlender, J.M.; Franke, S.; Stein, G.; Wolf, G. Advanced glycation end products and the kidney. Am. J. Physiol. Ren. Physiol. 2005, 289, F645–F659. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.; Franke, S.; Ruster, C.; Wolf, G. Advanced glycation end-products and the kidney. Eur. J. Clin. Investig. 2010, 40, 742–755. [Google Scholar] [CrossRef] [PubMed]
- Heidland, A.; Sebekova, K.; Schinzel, R. Advanced glycation end products and the progressive course of renal disease. Am. J. Kidney Dis. 2001, 38 (Suppl. S1), 100–106. [Google Scholar] [CrossRef]
- Wolf, G. Angiotensin II as a renal growth factor. Contrib. Nephrol. 2001, 135, 92–110. [Google Scholar] [PubMed]
- Chen, S.; Wolf, G.; Ziyadeh, F.N. The renin-angiotensin system in diabetic nephropathy. Contrib. Nephrol. 2001, 135, 212–221. [Google Scholar] [PubMed]
- Tsuchida, K.; Cronin, B.; Sharma, K. Novel aspects of transforming growth factor-beta in diabetic kidney disease. Nephron 2002, 92, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lee, J.S.; Iglesias-de la Cruz, M.C.; Wang, A.; Izquierdo-Lahuerta, A.; Gandhi, N.K.; Danesh, F.R.; Wolf, G.; Ziyadeh, F.N. Angiotensin ii stimulates alpha3(iv) collagen production in mouse podocytes via tgf-beta and vegf signalling: Implications for diabetic glomerulopathy. Nephrol. Dial. Transplant. 2005, 20, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Kwak, S.J.; Jung, D.S.; Kim, J.J.; Yoo, T.H.; Ryu, D.R.; Han, S.H.; Choi, H.Y.; Lee, J.E.; Moon, S.J.; et al. Podocyte biology in diabetic nephropathy. Kidney Int. Suppl. 2007, S36–S42. [Google Scholar] [CrossRef] [PubMed]
- Pellet-Many, C.; Frankel, P.; Jia, H.; Zachary, I. Neuropilins: Structure, function and role in disease. Biochem. J. 2008, 411, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Takagi, S.; Kasuya, Y.; Shimizu, M.; Matsuura, T.; Tsuboi, M.; Kawakami, A.; Fujisawa, H. Expression of a cell adhesion molecule, neuropilin, in the developing chick nervous system. Dev. Biol. 1995, 170, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chedotal, A.; He, Z.; Goodman, C.S.; Tessier-Lavigne, M. Neuropilin-2, a novel member of the neuropilin family, is a high affinity receptor for the semaphorins sema e and sema iv but not sema iii. Neuron 1997, 19, 547–559. [Google Scholar] [CrossRef]
- Robert, B.; Zhao, X.; Abrahamson, D.R. Coexpression of neuropilin-1, flk1, and vegf(164) in developing and mature mouse kidney glomeruli. Am. J. Physiol. Ren. Physiol. 2000, 279, F275–F282. [Google Scholar]
- Harper, S.J.; Xing, C.Y.; Whittle, C.; Parry, R.; Gillatt, D.; Peat, D.; Mathieson, P.W. Expression of neuropilin-1 by human glomerular epithelial cells in vitro and in vivo. Clin. Sci. (Lond.) 2001, 101, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Bondeva, T.; Ruster, C.; Franke, S.; Hammerschmid, E.; Klagsbrun, M.; Cohen, C.D.; Wolf, G. Advanced glycation end-products suppress neuropilin-1 expression in podocytes. Kidney Int. 2009, 75, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Villegas, G.; Tufro, A. Ontogeny of semaphorins 3a and 3f and their receptors neuropilins 1 and 2 in the kidney. Mech. Dev. 2002, 119 (Suppl. S1), 149–153. [Google Scholar] [CrossRef]
- Takagi, S.; Hirata, T.; Agata, K.; Mochii, M.; Eguchi, G.; Fujisawa, H. The a5 antigen, a candidate for the neuronal recognition molecule, has homologies to complement components and coagulation factors. Neuron 1991, 7, 295–307. [Google Scholar] [CrossRef]
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent semaphorin iii. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Kolodkin, A.L.; Levengood, D.V.; Rowe, E.G.; Tai, Y.T.; Giger, R.J.; Ginty, D.D. Neuropilin is a semaphorin iii receptor. Cell 1997, 90, 753–762. [Google Scholar] [CrossRef]
- Reza, J.N.; Gavazzi, I.; Cohen, J. Neuropilin-1 is expressed on adult mammalian dorsal root ganglion neurons and mediates semaphorin3a/collapsin-1-induced growth cone collapse by small diameter sensory afferents. Mol. Cell. Neurosci. 1999, 14, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhao, H.; Liao, Y.; Takashima, S.; Asano, Y.; Shintani, Y.; Hori, M.; Kitakaze, M. Higher mortality in heterozygous neuropilin-1 mice after cardiac pressure overload. Biochem. Biophys. Res. Commun. 2008, 370, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Soker, S.; Takashima, S.; Miao, H.Q.; Neufeld, G.; Klagsbrun, M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 1998, 92, 735–745. [Google Scholar] [CrossRef]
- Miao, H.Q.; Lee, P.; Lin, H.; Soker, S.; Klagsbrun, M. Neuropilin-1 expression by tumor cells promotes tumor angiogenesis and progression. FASEB J. 2000, 14, 2532–2539. [Google Scholar] [CrossRef] [PubMed]
- Tordjman, R.; Ortega, N.; Coulombel, L.; Plouet, J.; Romeo, P.H.; Lemarchandel, V. Neuropilin-1 is expressed on bone marrow stromal cells: A novel interaction with hematopoietic cells? Blood 1999, 94, 2301–2309. [Google Scholar] [PubMed]
- Glinka, Y.; Prud’homme, G.J. Neuropilin-1 is a receptor for transforming growth factor beta-1, activates its latent form, and promotes regulatory t cell activity. J. Leukoc. Biol. 2008, 84, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Sarris, M.; Andersen, K.G.; Randow, F.; Mayr, L.; Betz, A.G. Neuropilin-1 expression on regulatory t cells enhances their interactions with dendritic cells during antigen recognition. Immunity 2008, 28, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Kardos, J.; Gal, P.; Szilagyi, L.; Thielens, N.M.; Szilagyi, K.; Lorincz, Z.; Kulcsar, P.; Graf, L.; Arlaud, G.J.; Zavodszky, P. The role of the individual domains in the structure and function of the catalytic region of a modular serine protease, c1r. J. Immunol. 2001, 167, 5202–5208. [Google Scholar] [CrossRef] [PubMed]
- Delgadillo-Reynoso, M.G.; Rollo, D.R.; Hursh, D.A.; Raff, R.A. Structural analysis of the uegf gene in the sea urchin strongylocentrotus purpuratus reveals more similarity to vertebrate than to invertebrate genes with egf-like repeats. J. Mol. Evol. 1989, 29, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Hartigan, N.; Garrigue-Antar, L.; Kadler, K.E. Bone morphogenetic protein-1 (bmp-1). Identification of the minimal domain structure for procollagen c-proteinase activity. J. Biol. Chem. 2003, 278, 18045–18049. [Google Scholar] [CrossRef] [PubMed]
- Orchard-Webb, D.J.; Lee, T.C.; Cook, G.P.M.; Blair, G.E. Cub domain containing protein 1 (cdcp1) modulates adhesion and motility in colon cancer cells. BMC Cancer 2014, 14, 754. [Google Scholar] [CrossRef] [PubMed]
- Paavola, K.J.; Sidik, H.; Zuchero, J.B.; Eckart, M.; Talbot, W.S. Type iv collagen is an activating ligand for the adhesion g protein-coupled receptor gpr126. Sci. Signal. 2014, 7, ra76. [Google Scholar] [CrossRef] [PubMed]
- Appleton, B.A.; Wu, P.; Maloney, J.; Yin, J.; Liang, W.C.; Stawicki, S.; Mortara, K.; Bowman, K.K.; Elliott, J.M.; Desmarais, W.; et al. Structural studies of neuropilin/antibody complexes provide insights into semaphorin and vegf binding. EMBO J. 2007, 26, 4902–4912. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.W.; Xu, P.; Guo, H.F.; Vander Kooi, C.W. Mechanism of selective vegf-a binding by neuropilin-1 reveals a basis for specific ligand inhibition. PLoS ONE 2012, 7, e49177. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, G.P.; Haun, R.S.; Herzog, C.; Shah, S.V. Meprin a metalloproteinase and its role in acute kidney injury. Am. J. Physiol. Ren. Physiol. 2013, 304, F1150–F1158. [Google Scholar] [CrossRef] [PubMed]
- Aricescu, A.R.; Hon, W.C.; Siebold, C.; Lu, W.; van der Merwe, P.A.; Jones, E.Y. Molecular analysis of receptor protein tyrosine phosphatase mu-mediated cell adhesion. EMBO J. 2006, 25, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Cismasiu, V.B.; Denes, S.A.; Reilander, H.; Michel, H.; Szedlacsek, S.E. The mam (meprin/a5-protein/ptpmu) domain is a homophilic binding site promoting the lateral dimerization of receptor-like protein-tyrosine phosphatase mu. J. Biol. Chem. 2004, 279, 26922–26931. [Google Scholar] [CrossRef] [PubMed]
- Roth, L.; Nasarre, C.; Dirrig-Grosch, S.; Aunis, D.; Cremel, G.; Hubert, P.; Bagnard, D. Transmembrane domain interactions control biological functions of neuropilin-1. Mol. Biol. Cell 2008, 19, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Reed, R.R. Cloning and characterization of neuropilin-1-interacting protein: A psd-95/dlg/zo-1 domain-containing protein that interacts with the cytoplasmic domain of neuropilin-1. J. Neurosci. 1999, 19, 6519–6527. [Google Scholar] [PubMed]
- Parker, M.W.; Guo, H.F.; Li, X.; Linkugel, A.D.; Vander Kooi, C.W. Function of members of the neuropilin family as essential pleiotropic cell surface receptors. Biochemistry 2012, 51, 9437–9446. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, Q.; Ruhrberg, C. Neuropilin, you gotta let me know: Should i stay or should i go? Cell Adhes. Migr. 2010, 4, 61–66. [Google Scholar] [CrossRef]
- Nakamura, F.; Tanaka, M.; Takahashi, T.; Kalb, R.G.; Strittmatter, S.M. Neuropilin-1 extracellular domains mediate semaphorin d/iii-induced growth cone collapse. Neuron 1998, 21, 1093–1100. [Google Scholar] [CrossRef]
- Giger, R.J.; Urquhart, E.R.; Gillespie, S.K.; Levengood, D.V.; Ginty, D.D.; Kolodkin, A.L. Neuropilin-2 is a receptor for semaphorin iv: Insight into the structural basis of receptor function and specificity. Neuron 1998, 21, 1079–1092. [Google Scholar] [CrossRef]
- Rohm, B.; Ottemeyer, A.; Lohrum, M.; Puschel, A.W. Plexin/neuropilin complexes mediate repulsion by the axonal guidance signal semaphorin 3a. Mech. Dev. 2000, 93, 95–104. [Google Scholar] [CrossRef]
- Takahashi, T.; Fournier, A.; Nakamura, F.; Wang, L.H.; Murakami, Y.; Kalb, R.G.; Fujisawa, H.; Strittmatter, S.M. Plexin-neuropilin-1 complexes form functional semaphorin-3a receptors. Cell 1999, 99, 59–69. [Google Scholar] [CrossRef]
- Song, H.; Ming, G.; He, Z.; Lehmann, M.; McKerracher, L.; Tessier-Lavigne, M.; Poo, M. Conversion of neuronal growth cone responses from repulsion to attraction by cyclic nucleotides. Science 1998, 281, 1515–1518. [Google Scholar] [CrossRef] [PubMed]
- Lampropoulou, A.; Ruhrberg, C. Neuropilin regulation of angiogenesis. Biochem. Soc. Trans. 2014, 42, 1623–1628. [Google Scholar] [CrossRef] [PubMed]
- Lanahan, A.; Zhang, X.; Fantin, A.; Zhuang, Z.; Rivera-Molina, F.; Speichinge, K.; Prahst, C.; Zhang, J.; Wang, Y.; Davis, G.; et al. The neuropilin 1 cytoplasmic domain is required for vegf-a-dependent arteriogenesis. Dev. Cell 2013, 25, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N. Vascular endothelial growth factor receptors: Molecular mechanisms of activation and therapeutic potentials. Exp. Eye Res. 2006, 83, 1005–1016. [Google Scholar] [CrossRef] [PubMed]
- Soker, S.; Miao, H.Q.; Nomi, M.; Takashima, S.; Klagsbrun, M. Vegf165 mediates formation of complexes containing vegfr-2 and neuropilin-1 that enhance vegf165-receptor binding. J. Cell. Biochem. 2002, 85, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Herzog, B.; Pellet-Many, C.; Britton, G.; Hartzoulakis, B.; Zachary, I.C. Vegf binding to nrp1 is essential for vegf stimulation of endothelial cell migration, complex formation between nrp1 and vegfr2, and signaling via fak tyr407 phosphorylation. Mol. Biol. Cell 2011, 22, 2766–2776. [Google Scholar] [CrossRef] [PubMed]
- Zachary, I.C. How neuropilin-1 regulates receptor tyrosine kinase signalling: The knowns and known unknowns. Biochem. Soc. Trans. 2011, 39, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Klagsbrun, M.; Takashima, S.; Mamluk, R. The role of neuropilin in vascular and tumor biology. Adv. Exp. Med. Biol. 2002, 515, 33–48. [Google Scholar] [PubMed]
- Jones, E.A.; Yuan, L.; Breant, C.; Watts, R.J.; Eichmann, A. Separating genetic and hemodynamic defects in neuropilin 1 knockout embryos. Development 2008, 135, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Prud’homme, G.J.; Glinka, Y. Neuropilins are multifunctional coreceptors involved in tumor initiation, growth, metastasis and immunity. Oncotarget 2012, 3, 921–939. [Google Scholar] [PubMed]
- Kumanogoh, A.; Kikutani, H. Immunological functions of the neuropilins and plexins as receptors for semaphorins. Nat. Rev. Immunol. 2013, 13, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Pellet-Many, C.; Frankel, P.; Evans, I.M.; Herzog, B.M.; Junemann-Ramirez, M.; Zachary, I.C. Neuropilin-1 mediates pdgf stimulation of vascular smooth muscle cell migration and signalling via p130cas. Biochem. J. 2011, 435, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Glinka, Y.; Stoilova, S.; Mohammed, N.; Prud’homme, G.J. Neuropilin-1 exerts co-receptor function for tgf-beta-1 on the membrane of cancer cells and enhances responses to both latent and active tgf-beta. Carcinogenesis 2011, 32, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.G.; Shuttleworth, C.A.; Kielty, C.M. Platelet-derived growth factor receptors regulate mesenchymal stem cell fate: Implications for neovascularization. Expert Opin. Biol. Ther. 2010, 10, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Teesalu, T.; Sugahara, K.N.; Kotamraju, V.R.; Ruoslahti, E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proc. Natl. Acad. Sci. USA 2009, 106, 16157–16162. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Mukhopadhyay, D.; Xu, X. C terminus of rgs-gaip-interacting protein conveys neuropilin-1-mediated signaling during angiogenesis. FASEB J. 2006, 20, 1513–1515. [Google Scholar] [CrossRef] [PubMed]
- Naccache, S.N.; Hasson, T. Myosin vi altered at threonine 406 stabilizes actin filaments in vivo. Cell Motil. Cytoskeleton 2006, 63, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Seerapu, H.R.; Borthakur, S.; Kong, N.; Agrawal, S.; Drazba, J.; Vasanji, A.; Fantin, A.; Ruhrberg, C.; Buck, M.; Horowitz, A. The cytoplasmic domain of neuropilin-1 regulates focal adhesion turnover. FEBS Lett. 2013, 587, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Villegas, G.; Teichman, J.; Mundel, P.; Tufro, A. Autocrine class 3 semaphorin system regulates slit diaphragm proteins and podocyte survival. Kidney Int. 2006, 69, 1564–1569. [Google Scholar] [CrossRef] [PubMed]
- Bondeva, T.; Wolf, G. Advanced glycation end products suppress neuropilin-1 expression in podocytes by a reduction in sp1-dependent transcriptional activity. Am. J. Nephrol. 2009, 30, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, I.; Ruster, C.; Franke, S.; Liebisch, M.; Wolf, G. Erythropoietin ameliorates podocyte injury in advanced diabetic nephropathy in the db/db mouse. Am. J. Physiol. Ren. Physiol. 2013, 305, F911–F918. [Google Scholar] [CrossRef] [PubMed]
- Wendt, T.M.; Tanji, N.; Guo, J.; Kislinger, T.R.; Qu, W.; Lu, Y.; Bucciarelli, L.G.; Rong, L.L.; Moser, B.; Markowitz, G.S.; et al. Rage drives the development of glomerulosclerosis and implicates podocyte activation in the pathogenesis of diabetic nephropathy. Am. J. Pathol. 2003, 162, 1123–1137. [Google Scholar] [CrossRef]
- Rossignol, M.; Pouyssegur, J.; Klagsbrun, M. Characterization of the neuropilin-1 promoter; gene expression is mediated by the transcription factor sp1. J. Cell. Biochem. 2003, 88, 744–757. [Google Scholar] [CrossRef] [PubMed]
- Schramek, H.; Sarkozi, R.; Lauterberg, C.; Kronbichler, A.; Pirklbauer, M.; Albrecht, R.; Noppert, S.J.; Perco, P.; Rudnicki, M.; Strutz, F.M.; et al. Neuropilin-1 and neuropilin-2 are differentially expressed in human proteinuric nephropathies and cytokine-stimulated proximal tubular cells. Lab. Investig. 2009, 89, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.W.; Bennett, P.H.; Nelson, R.G. Podocyte number predicts long-term urinary albumin excretion in pima indians with type ii diabetes and microalbuminuria. Diabetologia 1999, 42, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Pagtalunan, M.E.; Miller, P.L.; Jumping-Eagle, S.; Nelson, R.G.; Myers, B.D.; Rennke, H.G.; Coplon, N.S.; Sun, L.; Meyer, T.W. Podocyte loss and progressive glomerular injury in type ii diabetes. J. Clin. Investig. 1997, 99, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Ushiyama, C.; Suzuki, S.; Hara, M.; Shimad, N.; Ebihara, I.; Koide, H. Urinary excretion of podocytes in patients with diabetic nephropathy. Nephrol. Dial. Transplant. 2000, 15, 1379–1383. [Google Scholar] [CrossRef] [PubMed]
- Petermann, A.T.; Pippin, J.; Krofft, R.; Blonski, M.; Griffin, S.; Durvasula, R.; Shankland, S.J. Viable podocytes detach in experimental diabetic nephropathy: Potential mechanism underlying glomerulosclerosis. Nephron Exp. Nephrol. 2004, 98. [Google Scholar] [CrossRef] [PubMed]
- Bondeva, T.; Wojciech, S.; Wolf, G. Advanced glycation end products inhibit adhesion ability of differentiated podocytes in a neuropilin-1-dependent manner. Am. J. Physiol. Ren. Physiol. 2011, 301. [Google Scholar] [CrossRef] [PubMed]
- Valdembri, D.; Caswell, P.T.; Anderson, K.I.; Schwarz, J.P.; Konig, I.; Astanina, E.; Caccavari, F.; Norman, J.C.; Humphries, M.J.; Bussolino, F.; et al. Neuropilin-1/gipc1 signaling regulates alpha5beta1 integrin traffic and function in endothelial cells. PLoS Biol. 2009, 7, e25. [Google Scholar] [CrossRef] [PubMed]
- Staton, C.A. Class 3 semaphorins and their receptors in physiological and pathological angiogenesis. Biochem. Soc. Trans. 2011, 39, 1565–1570. [Google Scholar] [CrossRef] [PubMed]
- Tufro, A.; Teichman, J.; Woda, C.; Villegas, G. Semaphorin3a inhibits ureteric bud branching morphogenesis. Mech. Dev. 2008, 125, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Reidy, K.J.; Villegas, G.; Teichman, J.; Veron, D.; Shen, W.; Jimenez, J.; Thomas, D.; Tufro, A. Semaphorin3a regulates endothelial cell number and podocyte differentiation during glomerular development. Development 2009, 136, 3979–3989. [Google Scholar] [CrossRef] [PubMed]
- Reidy, K.J.; Aggarwal, P.K.; Jimenez, J.J.; Thomas, D.B.; Veron, D.; Tufro, A. Excess podocyte semaphorin-3a leads to glomerular disease involving plexina1-nephrin interaction. Am. J. Pathol. 2013, 183, 1156–1168. [Google Scholar] [CrossRef] [PubMed]
- Reidy, K.; Tufro, A. Semaphorins in kidney development and disease: Modulators of ureteric bud branching, vascular morphogenesis, and podocyte-endothelial crosstalk. Pediatr. Nephrol. 2011, 26, 1407–1412. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, P.K.; Veron, D.; Thomas, D.B.; Siegel, D.; Moeckel, G.; Kashgarian, M.; Tufro, A. Semaphorin3a promotes advanced diabetic nephropathy. Diabetes 2015, 64, 1743–1759. [Google Scholar] [CrossRef] [PubMed]
- Eremina, V.; Baelde, H.J.; Quaggin, S.E. Role of the vegf—A signaling pathway in the glomerulus: Evidence for crosstalk between components of the glomerular filtration barrier. Nephron Physiol. 2007, 106, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Eremina, V.; Sood, M.; Haigh, J.; Nagy, A.; Lajoie, G.; Ferrara, N.; Gerber, H.P.; Kikkawa, Y.; Miner, J.H.; Quaggin, S.E. Glomerular-specific alterations of vegf-a expression lead to distinct congenital and acquired renal diseases. J. Clin. Investig. 2003, 111, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.R.; Saleem, M.A.; Mathieson, P.W.; Bates, D.O.; Harper, S.J. Vascular endothelial growth factor and nephrin interact and reduce apoptosis in human podocytes. Am. J. Physiol. Ren. Physiol. 2005, 288, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Foster, R.R.; Hole, R.; Anderson, K.; Satchell, S.C.; Coward, R.J.; Mathieson, P.W.; Gillatt, D.A.; Saleem, M.A.; Bates, D.O.; Harper, S.J. Functional evidence that vascular endothelial growth factor may act as an autocrine factor on human podocytes. Am. J. Physiol. Ren. Physiol. 2003, 284, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Guan, F.; Villegas, G.; Teichman, J.; Mundel, P.; Tufro, A. Autocrine vegf-a system in podocytes regulates podocin and its interaction with cd2ap. Am. J. Physiol. Ren. Physiol. 2006, 291, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Karnovsky, M.J.; Ainsworth, S.K. The structural basis of glomerular filtration. Adv. Nephrol. Necker Hosp. 1972, 2, 35–60. [Google Scholar] [PubMed]
- Rodewald, R.; Karnovsky, M.J. Porous substructure of the glomerular slit diaphragm in the rat and mouse. J. Cell Biol. 1974, 60, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Patrakka, J.; Tryggvason, K. Nephrin—A unique structural and signaling protein of the kidney filter. Trends Mol. Med. 2007, 13, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, V.; Ljungberg, P.; Wartiovaara, J.; Lenkkeri, U.; Kestila, M.; Jalanko, H.; Holmberg, C.; Tryggvason, K. Nephrin is specifically located at the slit diaphragm of glomerular podocytes. Proc. Natl. Acad. Sci. USA 1999, 96, 7962–7967. [Google Scholar] [CrossRef] [PubMed]
- Khoshnoodi, J.; Sigmundsson, K.; Ofverstedt, L.G.; Skoglund, U.; Obrink, B.; Wartiovaara, J.; Tryggvason, K. Nephrin promotes cell-cell adhesion through homophilic interactions. Am. J. Pathol. 2003, 163, 2337–2346. [Google Scholar] [CrossRef]
- Schnabel, E.; Anderson, J.M.; Farquhar, M.G. The tight junction protein zo-1 is concentrated along slit diaphragms of the glomerular epithelium. J. Cell Biol. 1990, 111, 1255–1263. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.G.; Ryu, D.R.; Yoo, T.H.; Jung, D.S.; Kim, J.J.; Kim, H.J.; Choi, H.Y.; Kim, J.S.; Adler, S.G.; Natarajan, R.; et al. P-cadherin is decreased in diabetic glomeruli and in glucose-stimulated podocytes in vivo and in vitro studies. Nephrol. Dial. Transplant. 2005, 20, 524–531. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Reiser, J.; Kriz, W.; Kretzler, M.; Mundel, P. The glomerular slit diaphragm is a modified adherens junction. J. Am. Soc. Nephrol. 2000, 11, 1–8. [Google Scholar] [PubMed]
- Ciani, L.; Patel, A.; Allen, N.D.; ffrench-Constant, C. Mice lacking the giant protocadherin mfat1 exhibit renal slit junction abnormalities and a partially penetrant cyclopia and anophthalmia phenotype. Mol. Cell. Biol. 2003, 23, 3575–3582. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Yaoita, E.; Kurihara, H.; Shimizu, F.; Sakai, T.; Kobayashi, T.; Ohshiro, K.; Kawachi, H.; Okada, H.; Suzuki, H.; et al. Fat is a component of glomerular slit diaphragms. Kidney Int. 2001, 59, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Donoviel, D.B.; Freed, D.D.; Vogel, H.; Potter, D.G.; Hawkins, E.; Barrish, J.P.; Mathur, B.N.; Turner, C.A.; Geske, R.; Montgomery, C.A.; et al. Proteinuria and perinatal lethality in mice lacking neph1, a novel protein with homology to nephrin. Mol. Cell. Biol. 2001, 21, 4829–4836. [Google Scholar] [CrossRef] [PubMed]
- Shih, N.Y.; Li, J.; Cotran, R.; Mundel, P.; Miner, J.H.; Shaw, A.S. Cd2ap localizes to the slit diaphragm and binds to nephrin via a novel c-terminal domain. Am. J. Pathol. 2001, 159, 2303–2308. [Google Scholar] [CrossRef]
- Wolf, G.; Stahl, R.A. CD2-associated protein and glomerular disease. Lancet 2003, 362, 1746–1748. [Google Scholar] [CrossRef]
- Shih, N.Y.; Li, J.; Karpitskii, V.; Nguyen, A.; Dustin, M.L.; Kanagawa, O.; Miner, J.H.; Shaw, A.S. Congenital nephrotic syndrome in mice lacking cd2-associated protein. Science 1999, 286, 312–315. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bondeva, T.; Wolf, G. Role of Neuropilin-1 in Diabetic Nephropathy. J. Clin. Med. 2015, 4, 1293-1311. https://doi.org/10.3390/jcm4061293
Bondeva T, Wolf G. Role of Neuropilin-1 in Diabetic Nephropathy. Journal of Clinical Medicine. 2015; 4(6):1293-1311. https://doi.org/10.3390/jcm4061293
Chicago/Turabian StyleBondeva, Tzvetanka, and Gunter Wolf. 2015. "Role of Neuropilin-1 in Diabetic Nephropathy" Journal of Clinical Medicine 4, no. 6: 1293-1311. https://doi.org/10.3390/jcm4061293
APA StyleBondeva, T., & Wolf, G. (2015). Role of Neuropilin-1 in Diabetic Nephropathy. Journal of Clinical Medicine, 4(6), 1293-1311. https://doi.org/10.3390/jcm4061293