
Pyodermatitis–Pyostomatitis Vegetans: The Role of Langerin Deficiency in Disease Pathogenesis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Research
2.2. Inclusion and Exclusion Criteria
2.3. Data Extraction and Quality Assessment
2.4. Immunohistochemistry
2.5. Data Analysis
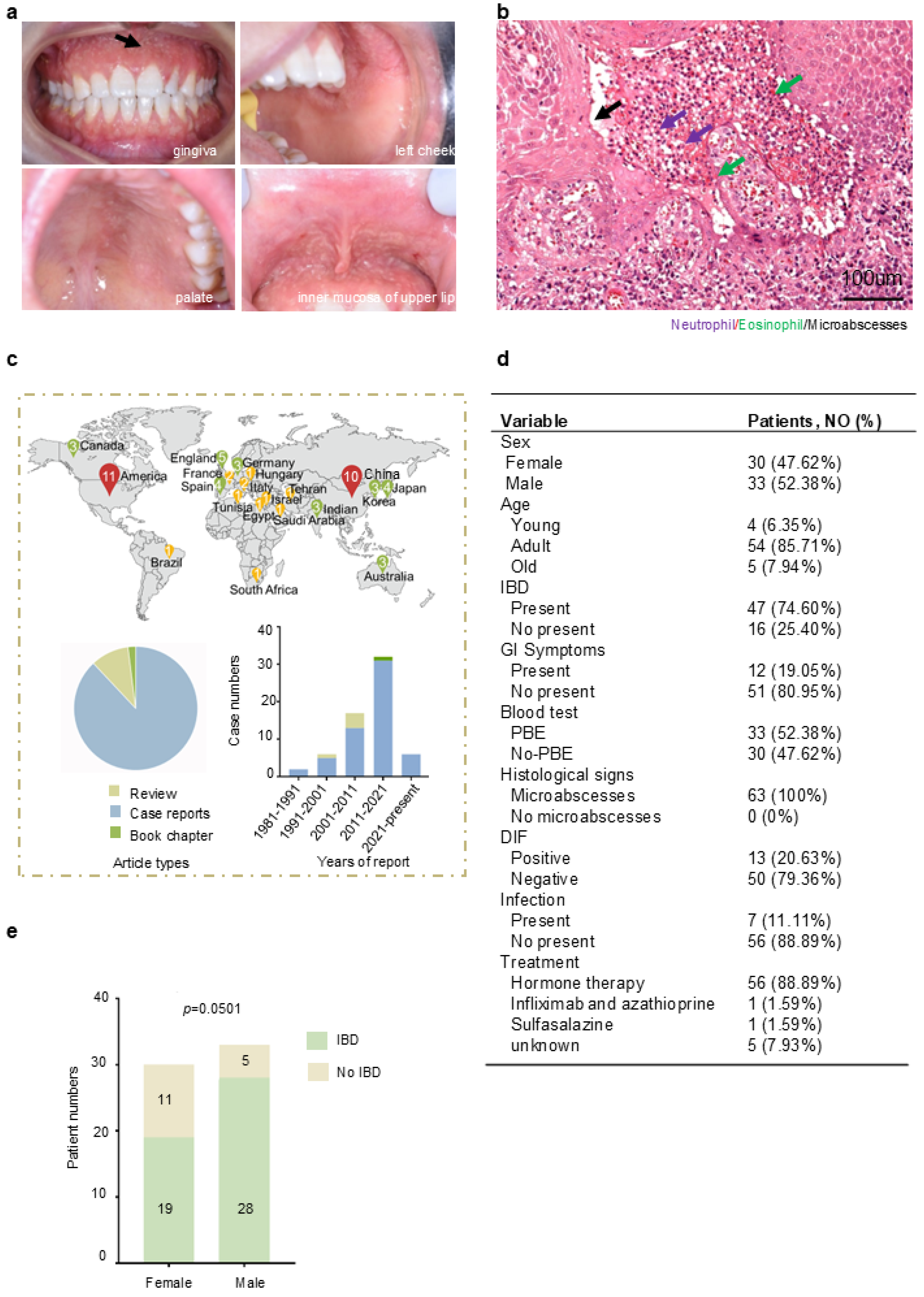
3. Summary from Literature Research
3.1. Current Status of Clinical Features, Pathology, and Treatment of PPV
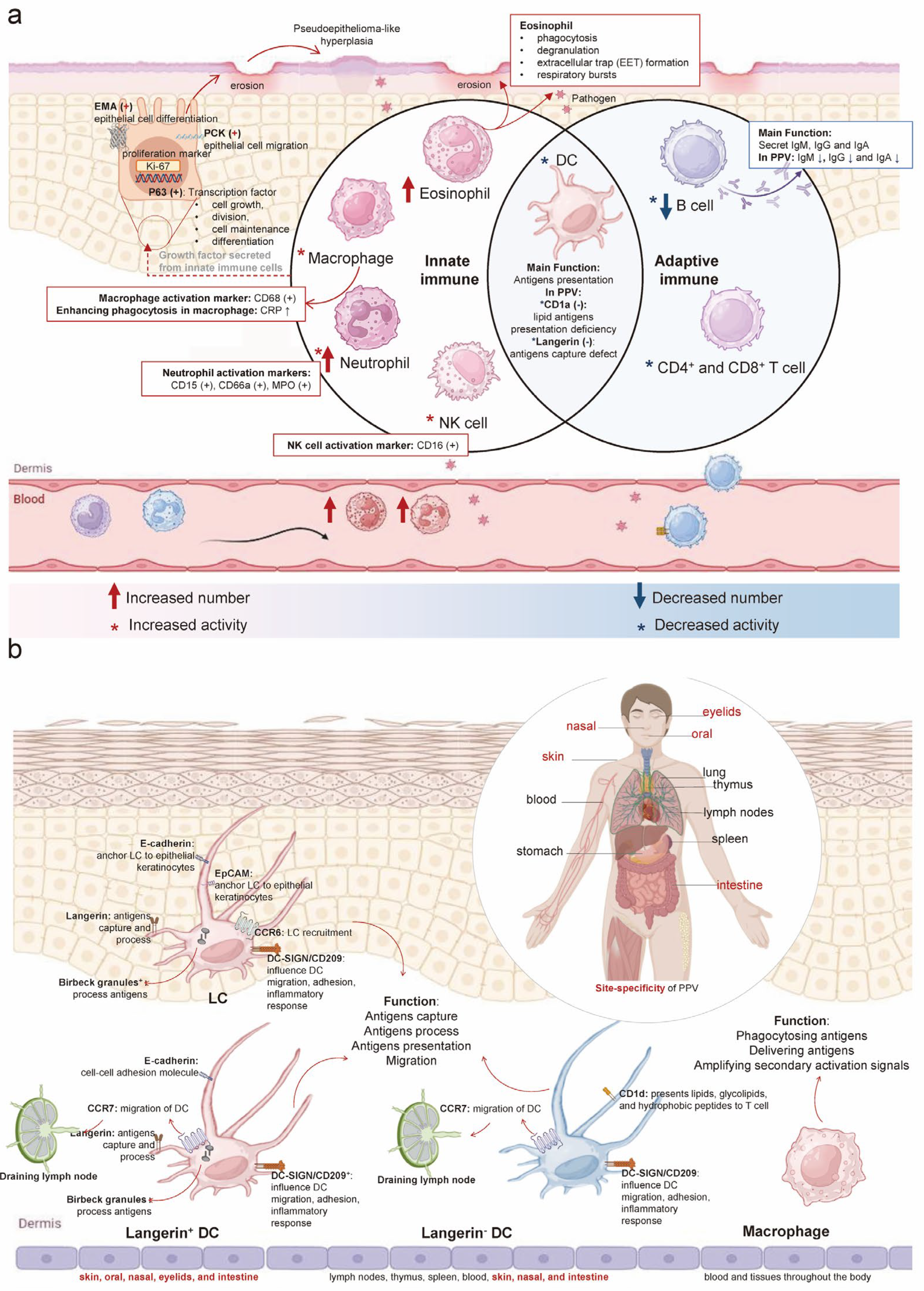
3.2. Enhancement of Innate Immunity and Weakening of Adaptive Immunity in PPV
4. Hypothesis from Literature Research
4.1. PPV Is an Inflammatory Disease Mediated by Langerin Deficiency
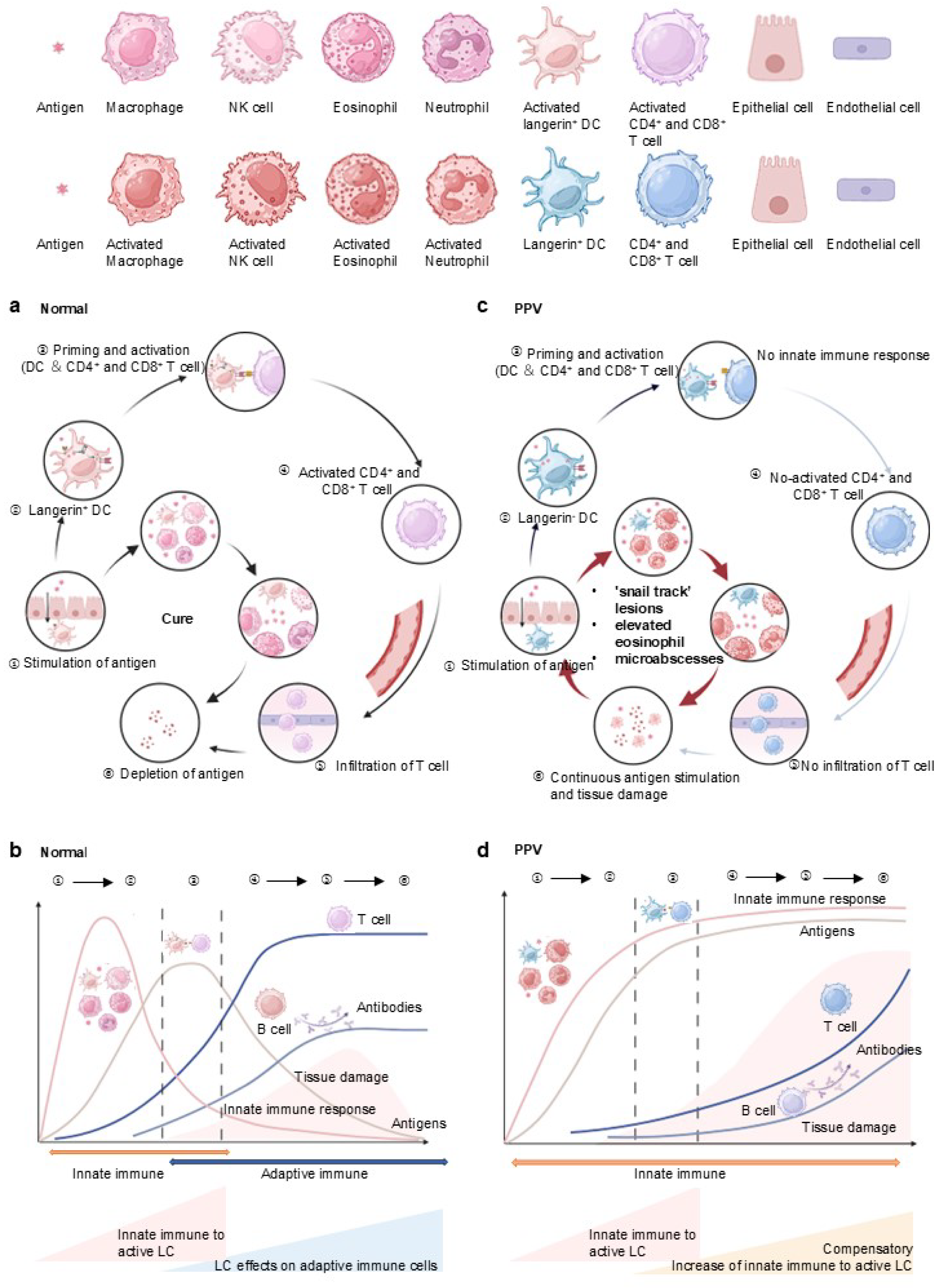
4.2. Enhanced Innate and Inactive Adaptive Immune Response Due to Langerin Deficiency in PPV
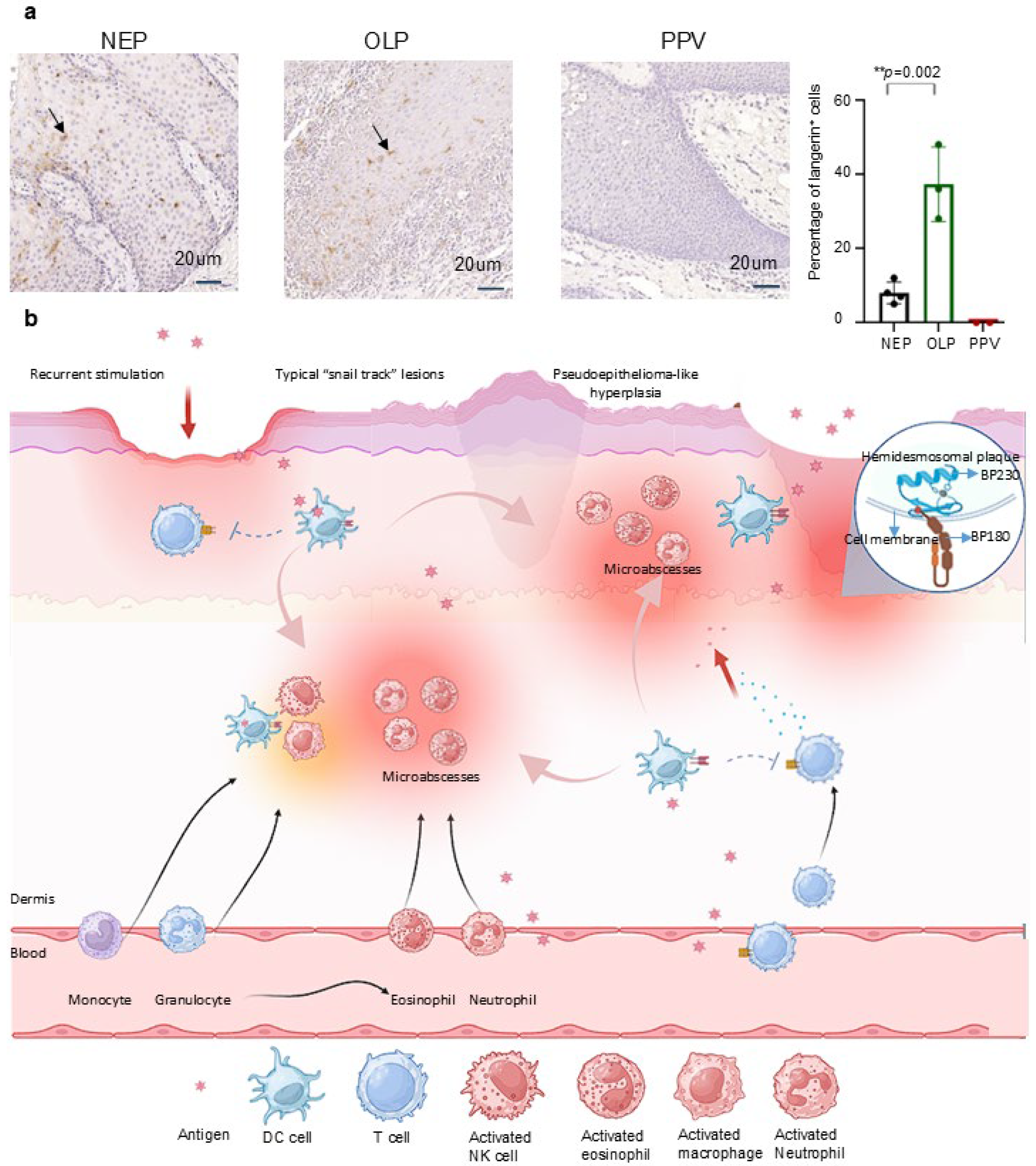
4.3. Immunohistochemical Staining Revealed Langerin Negative in PPV
4.4. The Deficiency of Langerin Interprets the PPV Clinical Pathological Manifestations
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gheisari, M.; Zerehpoosh, F.B.; Zaresharifi, S. Pyodermatitis-pyostomatitis vegetans: A case report and review of literature. Dermatol. Online J. 2020, 26, 5. [Google Scholar] [CrossRef]
- Fde, A.M.; Rosa, D.J.; Carvalho, M.T.; Castañon, M.C. Pyodermatitis-pyostomatitis vegetans: Case report and review of medical literature. An. Bras. Dermatol. 2011, 86, S137–S140. [Google Scholar]
- De Giorgi, V.; Trane, L.; Silvestri, F.; Venturi, F.; Zuccaro, B.; Scarfì, F. Pyodermatitis-Pyostomatitis Vegetans in Active Inflammatory Bowel Disease. Am. J. Gastroenterol. 2021, 116, 2163. [Google Scholar] [CrossRef]
- Ficarra, G.; Baroni, G.; Massi, D. Pyostomatitis vegetans: Cellular immune profile and expression of IL-6, IL-8 and TNF-alpha. Head Neck Pathol. 2010, 4, 1–9. [Google Scholar] [CrossRef]
- Ahn, B.K.; Kim, S.C. Pyodermatitis-pyostomatitis vegetans with circulating autoantibodies to bullous pemphigoid antigen 230. J. Am. Acad. Dermatol. 2004, 50, 785–788. [Google Scholar] [CrossRef]
- Kawahira, H.; Higashi, Y.; Matsuoka, A.; Fujii, K.; Ishii, N.; Hashimoto, T.; Kanekura, T. Pyodermatitis vegetans with antibodies to bullous pemphigoid antigen 180. J. Dermatol. 2017, 44, 1417–1418. [Google Scholar] [CrossRef]
- Storwick, G.S.; Prihoda, M.B.; Fulton, R.J.; Wood, W.S. Pyodermatitis-pyostomatitis vegetans: A specific marker for inflammatory bowel disease. J. Am. Acad. Dermatol. 1994, 31, 336–341. [Google Scholar] [CrossRef]
- Mehravaran, M.; Kemény, L.; Husz, S.; Korom, I.; Kiss, M.; Dobozy, A. Pyodermatitis-pyostomatitis vegetans. Br. J. Dermatol. 1997, 137, 266–269. [Google Scholar] [CrossRef]
- Naish, J.M.; Batchvarov, B.D.; Lawoyin, V.L. A case of ulcerative colitis and pyostomatitis vegetans in an African. Gut 1970, 11, 38–40. [Google Scholar] [CrossRef]
- Femiano, F.; Lanza, A.; Buonaiuto, C.; Perillo, L.; Dell’Ermo, A.; Cirillo, N. Pyostomatitis vegetans: A review of the literature. Med. Oral Patol. Oral Cir. Bucal 2009, 14, E114–E117. [Google Scholar]
- Gara, S.; Souissi, A.; Mokni, M. Pyodermatitis Pyostomatitis Vegetans. JAMA Dermatol. 2020, 156, 335. [Google Scholar] [CrossRef] [PubMed]
- Chrcanovic, B.R.; Martins-Chaves, R.R.; Pontes, F.S.C.; Fonseca, F.P.; Pontes, H.A.R.; Gomez, R.S. Pyodermatitis-pyostomatitis vegetans: A case report and systematic review focusing on oral involvement. Oral Maxillofac. Surg. 2024, 28, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Z.; Feng, S. Low-dose sulfasalazine in a case of pyodermatitis-pyostomatitis vegetans. J. Am. Acad. Dermatol. 2020, 82, e149–e150. [Google Scholar] [CrossRef]
- Fasoulas, K.; Kafalis, N.; Soufleris, K. Azathioprine-Induced Tremor: A Rare, Dose-Dependent Side Effect of Azathioprine in a Patient With Active Crohn Disease. Inflamm. Bowel Dis. 2020, 26, e72. [Google Scholar] [CrossRef]
- Dziąbowska-Grabias, K.; Sztanke, M.; Zając, P.; Celejewski, M.; Kurek, K.; Szkutnicki, S.; Korga, P.; Bulikowski, W.; Sztanke, K. Antioxidant Therapy in Inflammatory Bowel Diseases. Antioxidants 2021, 10, 412. [Google Scholar] [CrossRef]
- Iborra, M.; Beltrán, B.; Bastida, G.; Aguas, M.; Nos, P. Infliximab and adalimumab-induced psoriasis in Crohn’s disease: A paradoxical side effect. J. Crohn’s Colitis 2011, 5, 157–161. [Google Scholar] [CrossRef]
- Dodd, E.M.; Howard, J.R.; Dulaney, E.D.; Rosenthal, S.I.; Wanna, M.R.; Farah, R.S. Pyodermatitis-pyostomatitis vegetans associated with asymptomatic inflammatory bowel disease. Int. J. Dermatol. 2017, 56, 1457–1459. [Google Scholar] [CrossRef]
- Holness, C.L.; Simmons, D.L. Molecular cloning of CD68, a human macrophage marker related to lysosomal glycoproteins. Blood 1993, 81, 1607–1613. [Google Scholar] [CrossRef]
- Salazar, J.; Martínez, M.S.; Chávez-Castillo, M.; Núñez, V.; Añez, R.; Torres, Y.; Toledo, A.; Chacín, M.; Silva, C.; Pacheco, E.; et al. C-Reactive Protein: An In-Depth Look into Structure, Function, and Regulation. Int. Sch. Res. Not. 2014, 2014, 653045. [Google Scholar] [CrossRef]
- Ji, S.R.; Zhang, S.H.; Chang, Y.; Li, H.Y.; Wang, M.Y.; Lv, J.M.; Zhu, L.; Tang, P.M.K.; Wu, Y. C-Reactive Protein: The Most Familiar Stranger. J. Immunol. 2023, 210, 699–707. [Google Scholar] [CrossRef]
- Chen, Q.; Zeng, X. Case Based Oral Mucosal Diseases; Springer: Singapore, 2018. [Google Scholar]
- Radonjic-Hösli, S.; Simon, H.U. Eosinophils. Chem. Immunol. Allergy 2014, 100, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Gigon, L.; Fettrelet, T.; Yousefi, S.; Simon, D.; Simon, H.U. Eosinophils from A to Z. Allergy 2023, 78, 1810–1846. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- Ndlovu, L.N.; Peetluk, L.; Moodley, S.; Nhamoyebonde, S.; Ngoepe, A.T.; Mazibuko, M.; Khan, K.; Karim, F.; Pym, A.S.; Maruri, F.; et al. Increased Neutrophil Count and Decreased Neutrophil CD15 Expression Correlate With TB Disease Severity and Treatment Response Irrespective of HIV Co-infection. Front. Immunol. 2020, 11, 1872. [Google Scholar] [CrossRef]
- Li, S.; Cong, X.; Gao, H.; Lan, X.; Li, Z.; Wang, W.; Song, S.; Wang, Y.; Li, C.; Zhang, H.; et al. Tumor-associated neutrophils induce EMT by IL-17a to promote migration and invasion in gastric cancer cells. J. Exp. Clin. Cancer Res. CR 2019, 38, 6. [Google Scholar] [CrossRef]
- Rehring, J.F.; Bui, T.M.; Galán-Enríquez, C.S.; Urbanczyk, J.M.; Ren, X.; Wiesolek, H.L.; Sullivan, D.P.; Sumagin, R. Released Myeloperoxidase Attenuates Neutrophil Migration and Accumulation in Inflamed Tissue. Front. Immunol. 2021, 12, 654259. [Google Scholar] [CrossRef]
- Moretta, A.; Marcenaro, E.; Parolini, S.; Ferlazzo, G.; Moretta, L. NK cells at the interface between innate and adaptive immunity. Cell Death Differ. 2008, 15, 226–233. [Google Scholar] [CrossRef]
- Kitashima, D.Y.; Kobayashi, T.; Woodring, T.; Idouchi, K.; Doebel, T.; Voisin, B.; Adachi, T.; Ouchi, T.; Takahashi, H.; Nishifuji, K.; et al. Langerhans Cells Prevent Autoimmunity via Expansion of Keratinocyte Antigen-Specific Regulatory T Cells. EBioMedicine 2018, 27, 293–303. [Google Scholar] [CrossRef]
- Hardman, C.S.; Chen, Y.L.; Wegrecki, M.; Ng, S.W.; Murren, R.; Mangat, D.; Silva, J.P.; Munro, R.; Chan, W.Y.; O’Dowd, V.; et al. CD1a promotes systemic manifestations of skin inflammation. Nat. Commun. 2022, 13, 7535. [Google Scholar] [CrossRef]
- Pan, X.; Hobbs, R.P.; Coulombe, P.A. The expanding significance of keratin intermediate filaments in normal and diseased epithelia. Curr. Opin. Cell Biol. 2013, 25, 47–56. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Stojanovska, L.; Gargosky, S.E. MUC1 (CD227): A multi-tasked molecule. Cell. Mol. Life Sci. CMLS 2015, 72, 4475–4500. [Google Scholar] [CrossRef]
- Li, Y.; Giovannini, S.; Wang, T.; Fang, J.; Li, P.; Shao, C.; Wang, Y.; Shi, Y.; Candi, E.; Melino, G.; et al. p63: A crucial player in epithelial stemness regulation. Oncogene 2023, 42, 3371–3384. [Google Scholar] [CrossRef] [PubMed]
- King, K.E.; Weinberg, W.C. p63: Defining roles in morphogenesis, homeostasis, and neoplasia of the epidermis. Mol. Carcinog. 2007, 46, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.; Zhou, H. Master regulatory role of p63 in epidermal development and disease. Cell. Mol. Life Sci. CMLS 2018, 75, 1179–1190. [Google Scholar] [CrossRef] [PubMed]
- Remnant, L.; Kochanova, N.Y.; Reid, C.; Cisneros-Soberanis, F.; Earnshaw, W.C. The intrinsically disorderly story of Ki-67. Open Biol. 2021, 11, 210120. [Google Scholar] [CrossRef]
- Menon, S.S.; Guruvayoorappan, C.; Sakthivel, K.M.; Rasmi, R.R. Ki-67 protein as a tumour proliferation marker. Clin. Chim. Acta Int. J. Clin. Chem. 2019, 491, 39–45. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S. The cell biology of antigen presentation in dendritic cells. Curr. Opin. Immunol. 2001, 13, 45–51. [Google Scholar] [CrossRef]
- Kashem, S.W.; Haniffa, M.; Kaplan, D.H. Antigen-Presenting Cells in the Skin. Annu. Rev. Immunol. 2017, 35, 469–499. [Google Scholar] [CrossRef]
- Merad, M.; Ginhoux, F.; Collin, M. Origin, homeostasis and function of Langerhans cells and other langerin-expressing dendritic cells. Nat. Rev. Immunol. 2008, 8, 935–947. [Google Scholar] [CrossRef]
- Mathers, A.R.; Larregina, A.T. Professional antigen-presenting cells of the skin. Immunol. Res. 2006, 36, 127–136. [Google Scholar] [CrossRef]
- van der Vlist, M.; Geijtenbeek, T.B. Langerin functions as an antiviral receptor on Langerhans cells. Immunol. Cell Biol. 2010, 88, 410–415. [Google Scholar] [CrossRef] [PubMed]
- de Witte, L.; Nabatov, A.; Pion, M.; Fluitsma, D.; de Jong, M.A.; de Gruijl, T.; Piguet, V.; van Kooyk, Y.; Geijtenbeek, T.B. Langerin is a natural barrier to HIV-1 transmission by Langerhans cells. Nat. Med. 2007, 13, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Föhlinger, M.; Palamides, P.; Mansmann, U.; Beigel, F.; Siebeck, M.; Gropp, R. Immunological profiling of patients with ulcerative colitis leads to identification of two inflammatory conditions and CD1a as a disease marker. J. Transl. Med. 2016, 14, 310. [Google Scholar] [CrossRef]
- Hunger, R.E.; Sieling, P.A.; Ochoa, M.T.; Sugaya, M.; Burdick, A.E.; Rea, T.H.; Brennan, P.J.; Belisle, J.T.; Blauvelt, A.; Porcelli, S.A.; et al. Langerhans cells utilize CD1a and langerin to efficiently present nonpeptide antigens to T cells. J. Clin. Investig. 2004, 113, 701–708. [Google Scholar] [CrossRef]
- Verdijk, P.; Dijkman, R.; Plasmeijer, E.I.; Mulder, A.A.; Zoutman, W.H.; Mieke Mommaas, A.; Tensen, C.P. A lack of Birbeck granules in Langerhans cells is associated with a naturally occurring point mutation in the human Langerin gene. J. Investig. Dermatol. 2005, 124, 714–717. [Google Scholar] [CrossRef]
- Tang, A.; Amagai, M.; Granger, L.G.; Stanley, J.R.; Udey, M.C. Adhesion of epidermal Langerhans cells to keratinocytes mediated by E-cadherin. Nature 1993, 361, 82–85. [Google Scholar] [CrossRef]
- Borkowski, T.A.; Letterio, J.J.; Farr, A.G.; Udey, M.C. A role for endogenous transforming growth factor beta 1 in Langerhans cell biology: The skin of transforming growth factor beta 1 null mice is devoid of epidermal Langerhans cells. J. Exp. Med. 1996, 184, 2417–2422. [Google Scholar] [CrossRef]
- Jakob, T.; Ring, J.; Udey, M.C. Multistep navigation of Langerhans/dendritic cells in and out of the skin. J. Allergy Clin. Immunol. 2001, 108, 688–696. [Google Scholar] [CrossRef]
- Fleming, M.D.; Pinkus, J.L.; Fournier, M.V.; Alexander, S.W.; Tam, C.; Loda, M.; Sallan, S.E.; Nichols, K.E.; Carpentieri, D.F.; Pinkus, G.S.; et al. Coincident expression of the chemokine receptors CCR6 and CCR7 by pathologic Langerhans cells in Langerhans cell histiocytosis. Blood 2003, 101, 2473–2475. [Google Scholar] [CrossRef]
- Ohl, L.; Mohaupt, M.; Czeloth, N.; Hintzen, G.; Kiafard, Z.; Zwirner, J.; Blankenstein, T.; Henning, G.; Förster, R. CCR7 governs skin dendritic cell migration under inflammatory and steady-state conditions. Immunity 2004, 21, 279–288. [Google Scholar] [CrossRef]
- Ronger-Savle, S.; Valladeau, J.; Claudy, A.; Schmitt, D.; Peguet-Navarro, J.; Dezutter-Dambuyant, C.; Thomas, L.; Jullien, D. TGFbeta inhibits CD1d expression on dendritic cells. J. Investig. Dermatol. 2005, 124, 116–118. [Google Scholar] [CrossRef] [PubMed]
- van Kooyk, Y.; Geijtenbeek, T.B. DC-SIGN: Escape mechanism for pathogens. Nat. Rev. Immunol. 2003, 3, 697–709. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Chen, Y.; Hao, L.; Zhang, Y. DC-SIGN and immunoregulation. Cell. Mol. Immunol. 2006, 3, 279–283. [Google Scholar] [PubMed]
- Zhang, Q.; Cao, X. Epigenetic Remodeling in Innate Immunity and Inflammation. Annu. Rev. Immunol. 2021, 39, 279–311. [Google Scholar] [CrossRef]
- Garris, C.S.; Luke, J.J. Dendritic Cells, the T-cell-inflamed Tumor Microenvironment, and Immunotherapy Treatment Response. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 3901–3907. [Google Scholar] [CrossRef]
- Gustafson, J.; Eklund, C.; Wallström, M.; Zellin, G.; Magnusson, B.; Hasséus, B. Langerin-expressing and CD83-expressing cells in oral lichen planus lesions. Acta Odontol. Scand. 2007, 65, 156–161. [Google Scholar] [CrossRef]
- Solhaug, M.B.; Schreurs, O.; Schenck, K.; Blix, I.J.; Baekkevold, E.S. Origin of langerin (CD207)-expressing antigen presenting cells in the normal oral mucosa and in oral lichen planus lesions. Eur. J. Oral Sci. 2022, 130, e12835. [Google Scholar] [CrossRef]
- Xiao, C.; Zhu, Z.; Zhang, C.; Gao, J.; Luo, Y.; Fang, H.; Qiao, H.; Li, W.; Wang, G.; Fu, M. A population of dermal Langerin(+) dendritic cells promote the inflammation in mouse model of atopic dermatitis. Front. Immunol. 2022, 13, 981819. [Google Scholar] [CrossRef]
- Uzunçakmak, T.K.; Akdeniz, N.; Karadağ, A.S.; Çobanoğlu, B. Pyodermatitis vegetans after total colectomy. Indian Dermatol. Online J. 2015, 6, S9–S11. [Google Scholar] [CrossRef]
- Kissenpfennig, A.; Henri, S.; Dubois, B.; Laplace-Builhé, C.; Perrin, P.; Romani, N.; Tripp, C.H.; Douillard, P.; Leserman, L.; Kaiserlian, D.; et al. Dynamics and function of Langerhans cells in vivo: Dermal dendritic cells colonize lymph node areas distinct from slower migrating Langerhans cells. Immunity 2005, 22, 643–654. [Google Scholar] [CrossRef]
- Kervevan, J.; Bouteau, A.; Lanza, J.S.; Hammoudi, A.; Zurawski, S.; Surenaud, M.; Dieudonné, L.; Bonnet, M.; Lefebvre, C.; Hocini, H.; et al. Targeting human langerin promotes HIV-1 specific humoral immune responses. PLoS Pathog. 2021, 17, e1009749. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LC | Langerin+ DC | Langerin− DC | Macrophage | Location | |
|---|---|---|---|---|---|
| CD1a | + | + | + | − | Epithelial cell, B cell, dendritic cells, and thymocyte |
| Langerin | + | + | − | − | Skin, oral, nasal, eyelids, intestine, liver, lungs, lymph nodes, thymus, and spleen |
| Birbeck granules | + | + | − | − | Skin, oral, nasal, eyelids, and intestine |
| E-cadherin | + | + | − | − | Epithelial cell |
| EpCAM | + | − | − | − | Epithelial cell |
| CCR6 | + | − | − | − | All lymphatic and non-lymphoid tissues |
| CCR7 | − | + | + | − | Blood, bone marrow, lymph nodes, and intestines |
| CD1d | − | − | + | − | All antigen-presenting cells and thymic cortex cells |
| DC-SIGN/CD209 | + | + | + | − | Surface of immature dendritic cells |
| Location Site-specificity of PPV | Skin, oral, nasal, eyelids, and intestine | Skin, oral, nasal, eyelids, and intestine | Lymph nodes, thymus, spleen, blood, skin, nasal, and intestinal | Blood and tissues throughout the body |
| PPV | Current Status | New Insights |
|---|---|---|
| Clinic manifestations |
| |
| Mechanism | Unknown etiology, hypothesis shown as below:
| Langerin deficiency mediated disease
|
| Laboratory findings |
| |
| Histopathological manifestations | Infiltration of neutrophils and eosinophils | Infiltration of neutrophils and eosinophils. Immunohistochemical staining: negative langerin |
| Treatment | Hormone therapy Prednisolone sulfasalazine The combination of azathioprine and infliximab | Langerin target therapy to avoid the side effects of previous treatment |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, D.; Wang, J.; Cai, L.; Huang, M.; Han, Q.; Zheng, Z.; Zeng, X.; Chen, Q.; Ai, R.; Zhou, Y. Pyodermatitis–Pyostomatitis Vegetans: The Role of Langerin Deficiency in Disease Pathogenesis. J. Clin. Med. 2025, 14, 4198. https://doi.org/10.3390/jcm14124198
Pan D, Wang J, Cai L, Huang M, Han Q, Zheng Z, Zeng X, Chen Q, Ai R, Zhou Y. Pyodermatitis–Pyostomatitis Vegetans: The Role of Langerin Deficiency in Disease Pathogenesis. Journal of Clinical Medicine. 2025; 14(12):4198. https://doi.org/10.3390/jcm14124198
Chicago/Turabian StylePan, Dan, Jiongke Wang, Luyao Cai, Mei Huang, Qi Han, Zhijian Zheng, Xin Zeng, Qianming Chen, Ruixue Ai, and Yu Zhou. 2025. "Pyodermatitis–Pyostomatitis Vegetans: The Role of Langerin Deficiency in Disease Pathogenesis" Journal of Clinical Medicine 14, no. 12: 4198. https://doi.org/10.3390/jcm14124198
APA StylePan, D., Wang, J., Cai, L., Huang, M., Han, Q., Zheng, Z., Zeng, X., Chen, Q., Ai, R., & Zhou, Y. (2025). Pyodermatitis–Pyostomatitis Vegetans: The Role of Langerin Deficiency in Disease Pathogenesis. Journal of Clinical Medicine, 14(12), 4198. https://doi.org/10.3390/jcm14124198