
Application of Electroencephalography (EEG) in Combat Sports—Review of Findings, Perspectives, and Limitations


Abstract
1. Introduction
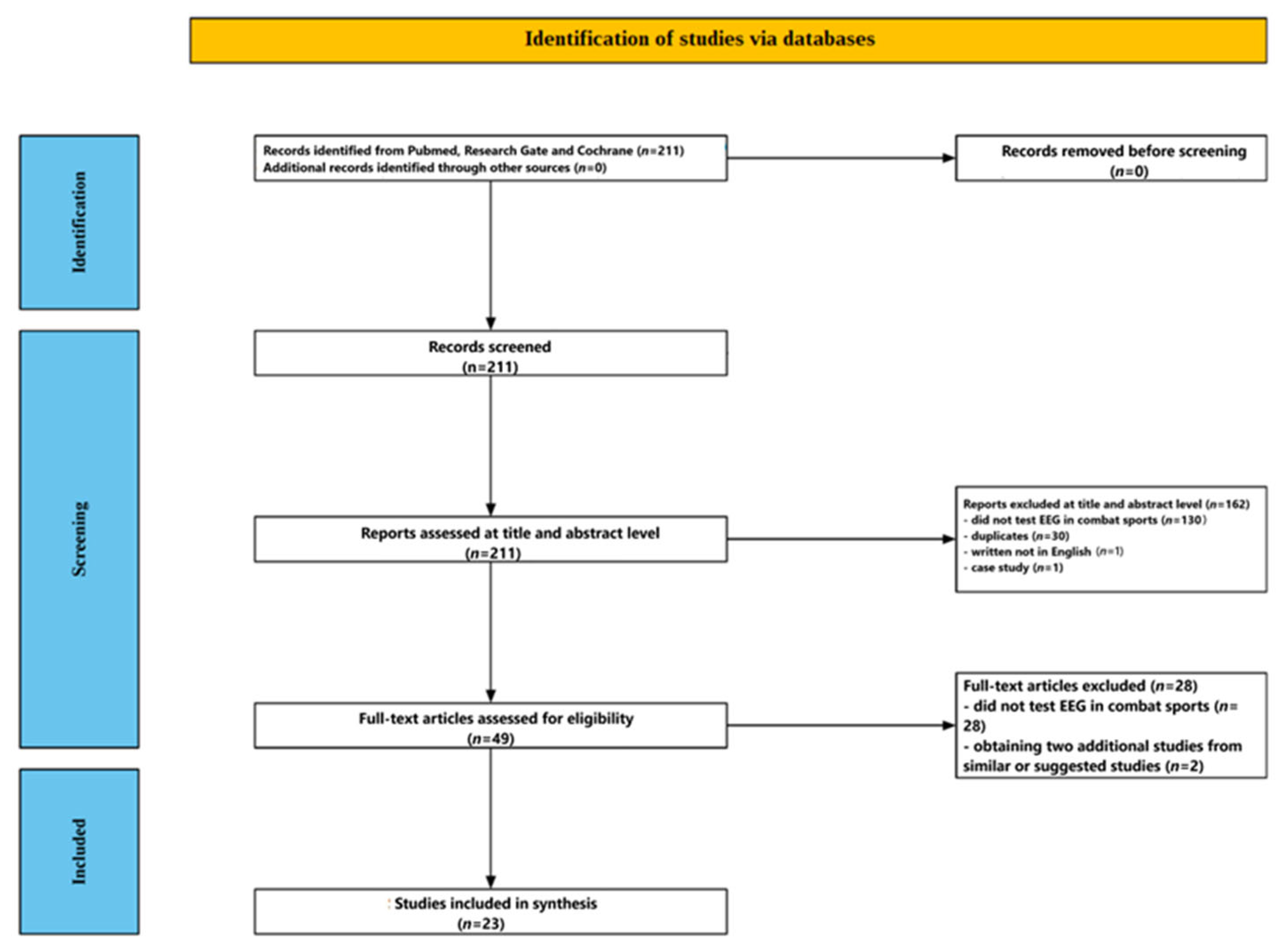
2. Methods
2.1. Data Sources and Search Strategy
2.2. Study Selection Criteria
2.3. Screening Process
2.3.1. Title and Abstract Screening
2.3.2. Full-Text Assessment
3. Results
3.1. Participant Characteristics
3.2. EEG Recording Parameters
3.2.1. Montage and Channel Count
3.2.2. Acquisition Conditions
3.2.3. Sampling Rate, Filtering, and Artifact Control
3.2.4. Spectral and Connectivity Analysis
3.2.5. Reference Schemes and Spatial Transforms
3.3. Time of EEG Recording in the Included Studies
3.3.1. Chronic “Off-Ring” Baselines (≥24 h After the Last Bout)
3.3.2. Laboratory Tasks and Short Controlled Maneuvers
3.3.3. Immediate Pre-Contest Readiness
3.3.4. Acute Post-Combat Recovery
3.4. Electrophysiological Outcomes
3.4.1. Multimodal Imaging Cohorts (Boxing vs. Judo)
3.4.2. Spectral and Network Changes in Boxing
3.4.3. Sport-Specific Resting Profiles
3.4.4. Task-Evoked and Perturbation Studies
Neural Efficiency Tests in Karate
3.4.5. Transient Grappling Maneuvers
3.4.6. Acute Competition Responses

{kind=link}
| Key EEG Findings | Analysis | EEG Set-Up | Control/Comparison | N | Sport/Population | Study |
|---|---|---|---|---|---|---|
| Slowing in 2 pros and 1 amateur; none in judoka; matched regional hypoperfusion | Visual scoring | 14 ch, eyes-closed rest | Age- and sex-matched healthy | 44 boxers + 10 judoka | Boxers (pro and am) + judoka | [39] |
| Abnormalities ↑ with number of bouts; no link to symptoms | Visual (+CT) | Clinical EEG (details NR) | None | 40 (24 EEG) | Former boxers | [40] |
| Abnormal EEG in 8 (40%); associated with younger age; 4 overlap with neuro signs | Visual | Clinical EEG | None | 20 | Active amateur boxers | [41] |
| Slight/moderate deviations in 32–36% of boxers; no severe or exposure correlation | Visual + BEAM | 21 ch, rest + photic | Soccer and track–field | 47 boxers + 25 soccer + 25 track | Former amateur boxers (HM/LM) | [42] |
| Advanced: ↓theta/alpha power, ↓ PLE and LRTC; AD-like entropy shift | FFT + PLE, DFA, MSE | 19 ch, 7 min EO/EC | Within-sport groups | 21 | Amateur boxers (beginner vs. advanced) | [43] |
| Boxers: hyperconnectivity but ↓ global/local efficiency and small-worldness | PLV and graph theory | 64 ch, 5 min EC | Non-athletes | 24 boxers/25 ctr | Active boxers vs. controls | [44] |
| Boxers show lower ratios→better attentional indices | Theta/β and Theta/SMR ratios | Single Cz, 1 min each cond. | Phys-ed students | 36/52 | Amateur boxers vs. students | [45] |
| Non-sig trend to lower alpha power in boxers | FFT (alpha) | 13 ch, 3 min EO/EC | Non-athletes | 7/9 | Amateur boxers vs. sedentary | [46] |
| ↓ Alpha power and ↑ reactivity coefficient in wrestlers | Band power, reactivity | C3 and C4, EC/EO/EC | Non-athletes | 30/30 | Elite wrestlers vs. controls | [47] |
| Elevated alpha, SMR, β; frontal asymmetry and hyper-arousal profile | QEEG (FFT) | 9 ch, EC | None | 18 | Elite K-1 kickboxers | [48] |
| Kickboxers: ↑ δ, θ, α, β across scalp; SMR ↑ Cz/P3 | QEEG | 9 ch, EO | Non-athletes | 18/18 | Elite K-1 vs. controls | [49] |
| Post-choke: ↑ δ/θ, ↓ α (occipital); resolves < 70 s | Spectral | 19 ch, EC | Pre- vs. post | 6 | Judoka (juji-jime choke) | [50] |
| Elite: lower ERD in dorsal and MNS→greater neural efficiency; aligns with rating accuracy | Alpha ERD, sLORETA | 56 ch | 3-group | 16/15/17 | Karate (elite/amateur/NA) judgment task | [51] |
| Elite: ↑ parietal/occipital α1 and δ/θ; trait marker of expertise | LORETA sources | 56 ch, 3 min EC | Multiple | 21/23/30 | Karate (elite/amateur/NA) rest (+ gymnast ctrl) | [52] |
| Athletes keep higher α/β and lower ERD under load | α/β ERD/ERS | 16 ch, EC rest→math | Matched controls | 10/10 | Elite karate vs. NA (mental arithmetic) | [53] |
| Smaller TRPD (less α desync) in athletes→reduced cortical reactivity | Alpha TRPD | 56 ch | Non-athletes | 18/28 | Elite karate vs. NA (EO vs. EC) | [54] |
| Athletes show lower desync during demanding stance | Alpha TRPD | 56 ch, bipodalic vs. monopodalic | Multi-group | 10 KAR/10 FEN/12 NA | Karate and fencing elites vs. NA (posture) | [55] |
| Only elites: strong parietal α ERD correlates with visual balance gain | Alpha ERD + sway | 56 ch | Multi-group | 19 KAR/18 FEN/10 NA | Karate and fencing elites vs. NA (stance EO/EC) | [56] |
| Elites: lower α cortex-muscle coherence; stable cortex→muscle drive | EEG-EMG coherence and DTF | 56 ch + EMG | Multi-group | 19 EL/14 AM/18 FEN/9 NA | Karate (elite/am) + fencing vs. NA | [57] |
| Reduced α ERD in motor areas during movement | Alpha ERD, sLORETA | 56 ch | Non-athletes | 10/12 | Elite karate vs. NA (wrist movement) | [58] |
| High β2 and θ; amplitudes higher with eyes closed→internal rehearsal/arousal | Theta and β2 amplitude | 9 ch | EO vs. EC within-subj | 15 | K-1 kickboxers pre-fight | [59] |
| Post-fight: ↑ δ and β2; magnitude correlates with head blows | QEEG bands | 9 ch, pre and ≤3 min post | Punchbag simulation | 50 fight/50 bag | K-1: fight vs. bag | [60] |
| Fight: ↑ SMR and β1 (F3/P3) with cortisol surge; T/C ratio drops | QEEG + T/C ratio | 9 ch, pre and post | Punchbag simulation | 50 fight/50 bag | K-1: fight vs. bag (+hormones) | [61] |
4. Discussion
4.1. Neural Efficiency: A Protective or Confounding Factor?
4.2. Evidence for Trauma-Related Dysfunction
4.3. Pathophysiological Integration
4.4. Discipline-Specific Phenotypes
4.5. State-Dependent Confounders
4.6. Relationship Between EEG Abnormalities and Clinical or Functional Indices
4.6.1. Exposure Load, Imaging, and EEG
4.6.2. Clinical Examination, Neuropsychology, and EEG
4.6.3. Performance-Specific or Stress-Related Couplings
5. Limitations and Future Directions
5.1. Methodological Heterogeneity
5.2. Sample Composition and Exposure Bias
5.3. Lack of Longitudinal, Multimodal Evidence
5.4. Relating Other Functional Outcomes of Athletes to EEG Outcomes
5.5. Translational and Regulatory Gaps
5.6. Taking State Confounders into Account
5.7. Under-Explored Research Questions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Franchini, E. Energy System Contributions during Olympic Combat Sports: A Narrative Review. Metabolites 2023, 13, 297. [Google Scholar] [CrossRef] [PubMed]
- Brechney, G.C.; Cannon, J.; Goodman, S.P. Effects of Weight Cutting on Exercise Performance in Combat Athletes: A Meta-Analysis. Int. J. Sports Physiol. Perform. 2022, 17, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Choma, C.W.; Sforzo, G.A.; Keller, B.A. Impact of rapid weight loss on cognitive function in collegiate wrestlers. Med. Sci. Sports Exerc. 1998, 30, 746–749. [Google Scholar] [CrossRef]
- Ceylan, B.; Aydos, L.; Šimenko, J. Effect of Rapid Weight Loss on Hydration Status and Performance in Elite Judo Athletes. Biology 2022, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Ruddock, A.; James, L.; French, D.; Rogerson, D.; Driller, M.; Hembrough, D. High-Intensity Conditioning for Combat Athletes: Practical Recommendations. Appl. Sci. 2021, 11, 10658. [Google Scholar] [CrossRef]
- Gonçalves, A.F.; Miarka, B.; Maurício, C.A.; Teixeira, R.P.A.; Brito, C.J.; Ignácio Valenzuela Pérez, D.; Slimani, M.; Znazen, H.; Bragazzi, N.L.; Reis, V.M. Enhancing performance: Unveiling the physiological impact of submaximal and supramaximal tests on mixed martial arts athletes in the −61 kg and −66 kg weight divisions. Front. Physiol. 2024, 14, 1257639, Erratum in Front. Physiol. 2024, 15, 1447631. [Google Scholar] [CrossRef]
- James, L.P.; Haff, G.G.; Kelly, V.G.; Beckman, E.M. Physiological determinants of mixed martial arts performance and method of competition outcome. Int. J. Sports Sci. Coach. 2018, 13, 978–984. [Google Scholar] [CrossRef]
- James, L.P.; Haff, G.G.; Kelly, V.G.; Beckman, E.M. Sports-specific conditioning: Physiological characteristics of mixed martial arts athletes. Sports Med. 2016, 46, 1319–1335. [Google Scholar] [CrossRef]
- Fernandes, J.R.; de Brito, M.A.; Brito, C.J.; Aedo-Munoz, E.; Miarka, B. Technical-tactical actions of fighters specialized in striking, grappling, and mixed combat in the Ultimate Fighting Championship. Ido Movement for Culture. J. Martial Arts Anthropol. 2022, 22, 23–31. [Google Scholar]
- Marinho, B.F.; Follmer, B.; Del Conti Esteves, J.V.; Andreato, L.V. Body composition, somatotype, and physical fitness of mixed martial arts athletes. Sport Sci. Health 2016, 12, 157–165. [Google Scholar] [CrossRef]
- Mojtahedi, D.; Dagnall, N.; Denovan, A.; Clough, P.; Dewhurst, S.; Hillier, M.; Papageorgiou, K.; Perry, J. Competition Anxiety in Combat Sports and the Importance of Mental Toughness. Behav. Sci. 2023, 13, 713. [Google Scholar] [CrossRef] [PubMed]
- Slimani, M.; Davis, P.; Franchini, E.; Moalla, W. Rating of Perceived Exertion for Quantification of Training and Combat Loads During Combat Sport-Specific Activities: A Short Review. J. Strength Cond. Res. 2017, 31, 2889–2902. [Google Scholar] [CrossRef] [PubMed]
- Mańka-Malara, K.; Mierzwińska-Nastalska, E. Head Trauma Exposure in Mixed Martial Arts. Int. J. Environ. Res. Public Health 2022, 19, 13050. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, R.R.; Ugbolue, U.C.; Gao, Y.; Gu, Y.; Dutheil, F.; Baker, J.S. A Systematic Review and Meta-Analysis Investigating Head Trauma in Boxing. Clin. J. Sport Med. 2023, 33, 658–674. [Google Scholar] [CrossRef]
- Lemme, N.J.; Johnston, B.; DeFroda, S.F.; Owens, B.D.; Kriz, P.K. Incidence of Combat Sport-Related Mild Traumatic Brain Injuries Presenting to the Emergency Department From 2012 to 2016. Clin. J. Sport Med. 2020, 30, 585–590. [Google Scholar] [CrossRef]
- Munakomi, S.; Puckett, Y. Chronic Traumatic Encephalopathy. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2025. Available online: https://www.ncbi.nlm.nih.gov/books/NBK541013/ (accessed on 12 May 2025).
- Montenigro, P.H.; Baugh, C.M.; Daneshvar, D.H.; Mez, J.; Budson, A.E.; Au, R.; Katz, D.I.; Cantu, R.C.; Stern, R.A. Clinical subtypes of chronic traumatic encephalopathy: Literature review and proposed research diagnostic criteria for traumatic encephalopathy syndrome. Alzheimers Res. Ther. 2014, 6, 68. [Google Scholar] [CrossRef]
- Maroon, J.C.; Winkelman, R.; Bost, J.; Amos, A.; Mathyssek, C.; Miele, V. Chronic traumatic encephalopathy in contact sports: A systematic review of all reported pathological cases. PLoS ONE 2015, 10, e0117338, Erratum in PLoS ONE 2015, 10, e0130507. [Google Scholar] [CrossRef]
- Monllor, P.; Cervera-Ferri, A.; Lloret, M.A.; Esteve, D.; Lopez, B.; Leon, J.L.; Lloret, A. Electroencephalography as a non-invasive biomarker of Alzheimer’s disease: A forgotten candidate to substitute CSF molecules? Int. J. Mol. Sci. 2021, 22, 10889. [Google Scholar] [CrossRef]
- Glomb, K.; Cabral, J.; Cattani, A.; Mazzoni, A.; Raj, A.; Franceschiello, B. Computational Models in Electroencephalography. Brain Topogr. 2022, 35, 142–161. [Google Scholar] [CrossRef]
- Light, G.A.; Williams, L.E.; Minow, F.; Sprock, J.; Rissling, A.; Sharp, R.; Swerdlow, N.R.; Braff, D.L. Electroencephalography (EEG) and event-related potentials (ERPs) with human participants. Curr. Protoc. Neurosci. 2010, 25, 1–24. [Google Scholar] [CrossRef]
- Bernardi, G.; Betta, M.; Ricciardi, E.; Pietrini, P.; Tononi, G.; Siclari, F. Regional Delta Waves In Human Rapid Eye Movement Sleep. J. Neurosci. 2019, 39, 2686–2697. [Google Scholar] [CrossRef] [PubMed]
- Cowdin, N.; Kobayashi, I.; Mellman, T.A. Theta frequency activity during rapid eye movement (REM) sleep is greater in people with resilience versus PTSD. Exp. Brain Res. 2014, 232, 1479–1485. [Google Scholar] [CrossRef] [PubMed]
- Schwabedal, J.T.; Riedl, M.; Penzel, T.; Wessel, N. Alpha-wave frequency characteristics in health and insomnia during sleep. J. Sleep Res. 2016, 25, 278–286. [Google Scholar] [CrossRef]
- Posada-Quintero, H.F.; Reljin, N.; Bolkhovsky, J.B.; Orjuela-Cañón, A.D.; Chon, K.H. Brain Activity Correlates With Cognitive Performance Deterioration During Sleep Deprivation. Front. Neurosci. 2019, 13, 1001. [Google Scholar] [CrossRef]
- Başar, E. A review of gamma oscillations in healthy subjects and in cognitive impairment. Int. J. Psychophysiol. 2013, 90, 99–117. [Google Scholar] [CrossRef]
- Gevins, A.; Smith, M.E.; McEvoy, L.K.; Leong, H.; Le, J. Electroencephalographic imaging of higher brain function. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 1125–1133. [Google Scholar] [CrossRef]
- Burle, B.; Spieser, L.; Roger, C.; Casini, L.; Hasbroucq, T.; Vidal, F. Spatial and temporal resolutions of EEG: Is it really black and white? A scalp current density view. Int. J. Psychophysiol. 2015, 97, 210–220. [Google Scholar] [CrossRef]
- Noachtar, S.; Rémi, J. The role of EEG in epilepsy: A critical review. Epilepsy Behav. 2009, 15, 22–33. [Google Scholar] [CrossRef]
- Zhao, W.; Van Someren, E.J.W.; Li, C.; Chen, X.; Gui, W.; Tian, Y.; Liu, Y.; Lei, X. EEG spectral analysis in insomnia disorder: A systematic review and meta-analysis. Sleep Med. Rev. 2021, 59, 101457. [Google Scholar] [CrossRef]
- Rubinos, C.; Bruzzone, M.J.; Viswanathan, V.; Figueredo, L.; Maciel, C.B.; LaRoche, S. Electroencephalography as a Biomarker of Prognosis in Acute Brain Injury. Semin. Neurol. 2023, 43, 675–688. [Google Scholar] [CrossRef]
- Morris, H.; Kaplan, P.W.; Kane, N. Electroencephalography in encephalopathy and encephalitis. Pract. Neurol. 2024, 24, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Thompson, T.; Steffert, T.; Ros, T.; Leach, J.; Gruzelier, J. EEG applications for sport and performance. Methods 2008, 45, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Fang, C.; Li, L.; Song, Y. Impact of sport training on adaptations in neural functioning and behavioral performance: A scoping review with meta-analysis on EEG research. J. Exerc. Sci. Fit. 2022, 20, 206–215. [Google Scholar] [CrossRef]
- Morrone, J.M.; Pedlar, C.R. EEG-based neurophysiological indices for expert psychomotor performance—A review. Brain Cogn. 2024, 175, 106132. [Google Scholar] [CrossRef]
- Cheron, G.; Petit, G.; Cheron, J.; Leroy, A.; Cebolla, A.; Cevallos, C.; Petieau, M.; Hoellinger, T.; Zarka, D.; Clarinval, A.M.; et al. Brain Oscillations in Sport: Toward EEG Biomarkers of Performance. Front. Psychol. 2016, 7, 246. [Google Scholar] [CrossRef]
- Ismail, L.; Karwowski, W.; Hancock, P.A.; Taiar, R.; Fernandez-Sumano, R. Electroencephalography (EEG) Physiological Indices Reflecting Human Physical Performance: A Systematic Review Using Updated PRISMA. J. Integr. Neurosci. 2023, 22, 62. [Google Scholar] [CrossRef]
- Conley, A.C.; Cooper, P.S.; Karayanidis, F.; Gardner, A.J.; Levi, C.R.; Stanwell, P.; Gaetz, M.B.; Iverson, G.L. Resting State Electroencephalography and Sports-Related Concussion: A Systematic Review. J. Neurotrauma, 2018; online ahead of print. 2018. [Google Scholar] [CrossRef]
- Rodriguez, G.; Vitali, P.; Nobili, F. Long-term effects of boxing and judo-choking techniques on brain function. Ital. J. Neurol. Sci. 1998, 19, 367–372. [Google Scholar] [CrossRef]
- Ross, R.J.; Cole, M.; Thompson, J.S.; Kim, K.H. Boxers—Computed tomography, EEG, and neurological evaluation. JAMA 1983, 249, 211–213. [Google Scholar] [CrossRef]
- McLatchie, G.; Brooks, N.; Galbraith, S.; Hutchison, J.S.; Wilson, L.; Melville, I.; Teasdale, E. Clinical neurological examination, neuropsychology, electroencephalography and computed tomographic head scanning in active amateur boxers. J. Neurol. Neurosurg. Psychiatry 1987, 50, 96–99. [Google Scholar] [CrossRef]
- Haglund, Y.; Persson, H.E. Does Swedish amateur boxing lead to chronic brain damage? 3. A retrospective clinical neurophysiological study. Acta Neurol. Scand. 1990, 82, 353–360. [Google Scholar] [CrossRef] [PubMed]
- De Donato, R.; Maiorana, N.V.; Vergari, M.; De Sandi, A.; Naci, A.; Aglieco, G.; Albizzati, T.; Guidetti, M.; Ferrara, R.; Bocci, T.; et al. ‘Knock down the brain’: A nonlinear analysis of electroencephalography to study the effects of sub-concussion in boxers. Eur. J. Neurol. 2025, 32, e16411. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Fu, Z.; Wang, S.; Zheng, F.; Qiu, W.; Xu, G.; Zhang, D.; Song, J. Disrupted Functional Brain Network Architecture in Sufferers with Boxing-Related Repeated Mild Traumatic Brain Injury: A Resting-State EEG Study. J. Integr. Neurosci. 2024, 23, 102. [Google Scholar] [CrossRef] [PubMed]
- Ziółkowski, A.; Gorkovenko, A.; Pasek, M.; Włodarczyk, P.; Zarańska, B.; Dornowski, M.; Graczyk, M. EEG Correlates of Attention Concentration in Successful Amateur Boxers. Neurophysiology 2014, 46, 422–427. [Google Scholar] [CrossRef]
- Çemç, M.S.; Korkmaz, O.E. Comparision of alpha frequency band power in active amateur boxers and sedentary individuals using EEG technology. Spormetre Beden Eğitimi Ve Spor Bilim. Derg. 2025, 23, 18–29. [Google Scholar]
- Yu Tarabrina, N. Features of the Brain Electrical Activity of Athletes-Fighters in Comparison with Non-athletes. Int. J. Hum. Mov. Sports Sci. 2023, 11, 902–907. [Google Scholar] [CrossRef]
- Rydzik, Ł.; Pałka, T.; Sobiło-Rydzik, E.; Tota, Ł.; Ambroży, D.; Ambroży, T.; Ruzbarsky, P.; Czarny, W.; Kopańska, M. An Attempt to Develop a Model of Brain Waves Using Quantitative Electroencephalography with Closed Eyes in K1 Kickboxing Athletes-Initial Concept. Sensors 2023, 23, 4136. [Google Scholar] [CrossRef]
- Rydzik, Ł.; Ambroży, T.; Pałka, T.; Wąsacz, W.; Spieszny, M.; Perliński, J.; Król, P.; Kopańska, M. Preliminary Development of a Brainwave Model for K1 Kickboxers Using Quantitative Electroencephalography (QEEG) with Open Eyes. Int. J. Mol. Sci. 2023, 24, 8882. [Google Scholar] [CrossRef]
- Rau, R.; Raschka, C.; Brunner, K.; Banzer, W. Spectral analysis of electroencephalography changes after choking in judo (juji-jime). Med. Sci. Sports Exerc. 1998, 30, 1356–1362. [Google Scholar]
- Babiloni, C.; Marzano, N.; Infarinato, F.; Iacoboni, M.; Rizza, G.; Aschieri, P.; Cibelli, G.; Soricelli, A.; Eusebi, F.; Del Percio, C. “Neural efficiency” of experts’ brain during judgment of actions: A high-resolution EEG study in elite and amateur karate athletes. Behav. Brain Res. 2010, 207, 466–475. [Google Scholar] [CrossRef]
- Babiloni, C.; Marzano, N.; Iacoboni, M.; Infarinato, F.; Aschieri, P.; Buffo, P.; Cibelli, G.; Soricelli, A.; Eusebi, F.; Del Percio, C. Resting state cortical rhythms in athletes: A high-resolution EEG study. Brain Res. Bull. 2010, 81, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Duru, A.D.; Assem, M. Investigating neural efficiency of elite karate athletes during a mental arithmetic task using EEG. Cogn. Neurodyn. 2018, 12, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Del Percio, C.; Infarinato, F.; Marzano, N.; Iacoboni, M.; Aschieri, P.; Lizio, R.; Soricelli, A.; Limatola, C.; Rossini, P.M.; Babiloni, C. Reactivity of alpha rhythms to eyes opening is lower in athletes than non-athletes: A high-resolution EEG study. Int. J. Psychophysiol. 2011, 82, 240–247. [Google Scholar] [CrossRef]
- Del Percio, C.; Babiloni, C.; Marzano, N.; Iacoboni, M.; Infarinato, F.; Vecchio, F.; Lizio, R.; Aschieri, P.; Fiore, A.; Toràn, G.; et al. “Neural efficiency” of athletes’ brain for upright standing: A high-resolution EEG study. Brain Res. Bull. 2009, 79, 193–200. [Google Scholar] [CrossRef]
- Del Percio, C.; Brancucci, A.; Bergami, F.; Marzano, N.; Fiore, A.; Di Ciolo, E.; Aschieri, P.; Lino, A.; Vecchio, F.; Iacoboni, M.; et al. Cortical alpha rhythms are correlated with body sway during quiet open-eyes standing in athletes: A high-resolution EEG study. Neuroimage 2007, 36, 822–829. [Google Scholar] [CrossRef]
- Vecchio, F.; Del Percio, C.; Marzano, N.; Fiore, A.; Toran, G.; Aschieri, P.; Gallamini, M.; Cabras, J.; Rossini, P.M.; Babiloni, C.; et al. Functional cortico-muscular coupling during upright standing in athletes and nonathletes: A coherence electroencephalographic-electromyographic study. Behav. Neurosci. 2008, 122, 917–927. [Google Scholar] [CrossRef]
- Del Percio, C.; Infarinato, F.; Iacoboni, M.; Marzano, N.; Soricelli, A.; Aschieri, P.; Eusebi, F.; Babiloni, C. Movement-related desynchronization of alpha rhythms is lower in athletes than non-athletes: A high-resolution EEG study. Clin. Neurophysiol. 2010, 121, 482–491. [Google Scholar] [CrossRef]
- Rydzik, L.; Wąsacz, W.; Ambroży, T.; Kasicki, K.; Ratkowski, W.; Kopańska, M. Kickboxing on Theta and Beta2 waves: Unravelling the mind’s secrets through QEEG analysis. Arch. Budo Sci. Martial Arts Extrem. Sports 2023, 19, 211–218. [Google Scholar]
- Rydzik, Ł.; Kopańska, M.; Wąsacz, W.; Ouergui, I.; Obmiński, Z.; Pałka, T.; Ambroży, T.; Malliaropoulos, N.; Maffulli, N.; Lota, K.S.; et al. Brain Punch: K-1 Fights Affect Brain Wave Activity in Professional Kickboxers. Sports Med. 2024, 54, 3169–3179. [Google Scholar] [CrossRef]
- Rydzik, Ł.; Obmiński, Z.; Wąsacz, W.; Kopańska, M.; Kubacki, R.; Bagińska, M.; Tota, Ł.; Ambroży, T.; Witkowski, K.; Pałka, T. The effect of physical exercise during competitions and in simulated conditions on hormonal-neurophysiological relationships in kickboxers. Biol. Sport 2024, 41, 61–68. [Google Scholar] [CrossRef]
- Haier, R.J.; Siegel, B.V.; Nuechterlein, K.H.; Hazlett, E.; Wu, J.C.; Paek, J.; Browning, H.L.; Buchsbaum, M.S. Cortical glucose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. Intelligence 1988, 12, 199–217. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef] [PubMed]
- Filho, E.; Dobersek, U.; Husselman, T.A. The role of neural efficiency, transient hypofrontality and neural proficiency in optimal performance in self-paced sports: A meta-analytic review. Exp. Brain Res. 2021, 239, 1381–1393. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Smith, D.M. Neural Efficiency in Athletes: A Systematic Review. Front. Behav. Neurosci. 2021, 15, 698555, Erratum in Front. Behav. Neurosci. 2022, 16, 841772. [Google Scholar] [CrossRef]
- Martins, L.C.X.; Russo, M.T.; Ribeiro, P. Neural Correlates of Shooting Sports Performance: A Systematic Review on Neural Efficiency Hypothesis. Revista De Educação Física/J. Phys. Educ. 2023, 91, 350–374. [Google Scholar] [CrossRef]
- Jensen, O.; Mazaheri, A. Shaping functional architecture by oscillatory alpha activity: Gating by inhibition. Front. Hum. Neurosci. 2010, 4, 186. [Google Scholar] [CrossRef]
- Ianof, J.N.; Anghinah, R. Traumatic brain injury: An EEG point of view. Dement. Neuropsychol. 2017, 11, 3–5. [Google Scholar] [CrossRef]
- Kadri, A.; Apriani, N. Electroencephalography Findings in Traumatic Brain Injury. Open Neurol. J. 2022, 16, e1874205X2206100. [Google Scholar] [CrossRef]
- Haneef, Z.; Levin, H.S.; Frost, J.D., Jr.; Mizrahi, E.M. Electroencephalography and quantitative electroencephalography in mild traumatic brain injury. J. Neurotrauma 2013, 30, 653–656. [Google Scholar] [CrossRef]
- Mathewson, K.E.; Lleras, A.; Beck, D.M.; Fabiani, M.; Ro, T.; Gratton, G. Pulsed out of awareness: EEG alpha oscillations represent a pulsed-inhibition of ongoing cortical processing. Front. Psychol. 2011, 2, 99. [Google Scholar] [CrossRef]
- Jensen, O. Distractor inhibition by alpha oscillations is controlled by an indirect mechanism governed by goal-relevant information. Commun. Psychol. 2024, 2, 36. [Google Scholar] [CrossRef] [PubMed]
- Oeltzschner, G.; Zöllner, H.J.; Hui, S.C.N.; Tapper, S.; Mikkelsen, M.; Raisanen, J.M.; Edden, R.A.E. Connecting occipital alpha-band peak frequency, visual temporal resolution and occipital GABA levels in healthy participants and hepatic-encephalopathy patients. NeuroImage Clin. 2018, 19, 102–111. [Google Scholar]
- Brown, J.C.; Goldszer, I.M.; Brooks, M.C.; Milano, N.J. An Evaluation of the Emerging Techniques in Sports-Related Concussion. J. Clin. Neurophysiol. 2023, 40, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Alosco, M.L.; Armañanzas, R.; Martin, B.M.; Tripodis, Y.; Stern, R.A.; Prichep, L.S. Long-Term Changes in Brain Connectivity Reflected in Quantitative Electrophysiology of Symptomatic Former National Football League Players. J. Neurotrauma 2023, 40, 309–317. [Google Scholar] [CrossRef]
- Sponheim, S.R.; McGuire, K.A.; Kang, S.S.; Davenport, N.D.; Aviyente, S.; Bernat, E.M.; Lim, K.O. Evidence of disrupted functional connectivity in the brain after combat-related blast injury. Neuroimage 2011, 54 (Suppl. S1), S21–S29. [Google Scholar] [CrossRef]
- Wang, C.; Costanzo, M.E.; Rapp, P.E.; Darmon, D.; Nathan, D.E.; Bashirelahi, K.; Pham, D.L.; Roy, M.J.; Keyser, D.O. Disrupted Gamma Synchrony after Mild Traumatic Brain Injury and Its Correlation with White Matter Abnormality. Front. Neurol. 2017, 8, 571. [Google Scholar] [CrossRef]
- Shenoy Handiru, V.; Alivar, A.; Hoxha, A.; Saleh, S.; Suviseshamuthu, E.S.; Yue, G.H.; Allexandre, D. Graph-theoretical analysis of EEG functional connectivity during balance perturbation in traumatic brain injury: A pilot study. Hum. Brain Mapp. 2021, 42, 4427–4447. [Google Scholar] [CrossRef]
- Shahim, P.; Politis, A.; van der Merwe, A.; Moore, B.; Chou, Y.Y.; Pham, D.L.; Butman, J.A.; Diaz-Arrastia, R.; Gill, J.M.; Brody, D.L.; et al. Neurofilament light as a biomarker in traumatic brain injury. Neurology 2020, 95, e610–e622. [Google Scholar] [CrossRef]
- May, H.G.; Tsikonofilos, K.; Donat, C.K.; Sastre, M.; Kozlov, A.S.; Sharp, D.J.; Bruyns-Haylett, M. EEG hyperexcitability and hyperconnectivity linked to GABAergic inhibitory interneuron loss following traumatic brain injury. Brain Commun. 2024, 6, fcae385. [Google Scholar] [CrossRef]
- Wang, Y.; Bartels, H.M.; Nelson, L.D. A Systematic Review of ASL Perfusion MRI in Mild TBI. Neuropsychol. Rev. 2023, 33, 160–191. [Google Scholar] [CrossRef]
- Foreman, B.; Claassen, J. Quantitative EEG for the detection of brain ischemia. Crit. Care 2012, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Konduru, S.S.; Wallace, E.P.; Pfammatter, J.A.; Rodrigues, P.V.; Jones, M.V.; Maganti, R.K. Sleep-wake characteristics in a mouse model of severe traumatic brain injury: Relation to posttraumatic epilepsy. Epilepsia Open 2021, 6, 181–194. [Google Scholar] [CrossRef]
- Vasilevskaya, A.; Anastassiadis, C.; Thapa, S.; Taghdiri, F.; Khodadadi, M.; Multani, N.; Rusjan, P.; Ozzoude, M.; Tarazi, A.; Mushtaque, A.; et al. 18F-Flortaucipir (AV1451) imaging identifies grey matter atrophy in retired athletes. J. Neurol. 2024, 271, 6068–6079. [Google Scholar] [CrossRef] [PubMed]
- Seeck, M.; Koessler, L.; Bast, T.; Leijten, F.; Michel, C.; Baumgartner, C.; He, B.; Beniczky, S. The standardized EEG electrode array of the IFCN. Clin. Neurophysiol. 2017, 128, 2070–2077. [Google Scholar] [CrossRef]
- Qin, Y.; Xu, P.; Yao, D. A comparative study of different references for EEG default mode network: The use of the infinity reference. Clin. Neurophysiol. 2010, 121, 1981–1991. [Google Scholar] [CrossRef]
- Ono, K.E.; Burns, T.G.; Bearden, D.J.; McManus, S.M.; King, H.; Reisner, A. Sex-Based Differences as a Predictor of Recovery Trajectories in Young Athletes After a Sports-Related Concussion. Am. J. Sports Med. 2016, 44, 748–752. [Google Scholar] [CrossRef]
- Merritt, V.C.; Padgett, C.R.; Jak, A.J. A systematic review of sex differences in concussion outcome: What do we know? Clin. Neuropsychol. 2019, 33, 1016–1043. [Google Scholar] [CrossRef]
- King, N.S. A systematic review of age and gender factors in prolonged post-concussion symptoms after mild head injury. Brain Inj. 2014, 28, 1639–1645. [Google Scholar] [CrossRef]
- McGroarty, N.K.; Brown, S.M.; Mulcahey, M.K. Sport-Related Concussion in Female Athletes: A Systematic Review. Orthop. J. Sports Med. 2020, 8, 2325967120932306. [Google Scholar] [CrossRef]
- Iverson, G.L.; Gardner, A.J.; Terry, D.P.; Ponsford, J.L.; Sills, A.K.; Broshek, D.K.; Solomon, G.S. Predictors of clinical recovery from concussion: A systematic review. Br. J. Sports Med. 2017, 51, 941–948. [Google Scholar] [CrossRef]
- Davenport, E.M.; Urban, J.E.; Vaughan, C.; DeSimone, J.C.; Wagner, B.; Espeland, M.A.; Powers, A.K.; Whitlow, C.T.; Stitzel, J.D.; Maldjian, J.A. MEG measured delta waves increase in adolescents after concussion. Brain Behav. 2022, 12, e2720. [Google Scholar] [CrossRef] [PubMed]
- Wayment, H.A.; Huffman, A.H.; Lane, T.S.; Lininger, M.R. Relationship of athletic and academic identity to concussion reporting intentions. Musculoskelet. Sci. Pract. 2019, 42, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Goh, S.Y.; Irimia, A.; Torgerson, C.M.; Horn, J.D. Neuroinformatics challenges to the structural, connectomic, functional and electrophysiological multimodal imaging of human traumatic brain injury. Front. Neuroinform. 2014, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, J.D.; Bhattrai, A.; Irimia, A. Multimodal Imaging of Neurometabolic Pathology due to Traumatic Brain Injury. Trends Neurosci. 2017, 40, 39–59. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Shah, S.A.; Lowder, R.J.; Kuceyeski, A. Quantitative multimodal imaging in traumatic brain injuries producing impaired cognition. Curr. Opin. Neurol. 2020, 33, 691–698. [Google Scholar] [CrossRef]
- Alosco, M.L.; Tripodis, Y.; Baucom, Z.H.; Mez, J.; Stein, T.D.; Martin, B.; Haller, O.; Conneely, S.; McClean, M.; Nosheny, R.; et al. Late contributions of repetitive head impacts and TBI to depression symptoms and cognition. Neurology 2020, 95, e793–e804. [Google Scholar] [CrossRef]
- Mishra, V.R.; Zhuang, X.; Sreenivasan, K.R.; Banks, S.J.; Yang, Z.; Bernick, C.; Cordes, D. Multimodal MR imaging signatures of cognitive impairment in active professional fighters. Radiology 2017, 285, 555–567. [Google Scholar] [CrossRef]
- Meghdadi, A.H.; Stevanović Karić, M.; McConnell, M.; Rupp, G.; Richard, C.; Hamilton, J.; Salat, D.; Berka, C. Resting state EEG biomarkers of cognitive decline associated with Alzheimer’s disease and mild cognitive impairment. PLoS ONE 2021, 16, e0244180. [Google Scholar] [CrossRef]
- Tanaka, M.; Yamada, E.; Mori, F. Neurophysiological markers of early cognitive decline in older adults: A mini-review of electroencephalography studies for precursors of dementia. Front. Aging Neurosci. 2024, 16, 1486481. [Google Scholar] [CrossRef]
- Cave, A.E.; De Blasio, F.M.; Chang, D.H.; Münch, G.W.; Steiner-Lim, G.Z. Eyes-open and eyes-closed EEG of older adults with subjective cognitive impairment versus healthy controls: A frequency principal components analysis study. Brain Res. 2025, 1850, 149399. [Google Scholar] [CrossRef]
- Katayama, O.; Stern, Y.; Habeck, C.; Coors, A.; Lee, S.; Harada, K.; Makino, K.; Tomida, K.; Morikawa, M.; Yamaguchi, R.; et al. Detection of neurophysiological markers of cognitive reserve: An EEG study. Front. Aging Neurosci. 2024, 16, 1401818. [Google Scholar] [CrossRef] [PubMed]
- Banks, S.J.; Obuchowski, N.; Shin, W.; Lowe, M.; Phillips, M.; Modic, M.; Bernick, C. The protective effect of education on cognition in professional fighters. Arch. Clin. Neuropsychol. 2014, 29, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Tenney, J.R.; Gloss, D.; Arya, R.; Kaplan, P.W.; Lesser, R.; Sexton, V.; Nuwer, M. Practice Guideline: Use of Quantitative EEG for the Diagnosis of Mild Traumatic Brain Injury: Report of the Guideline Committee of the American Clinical Neurophysiology Society. J. Clin. Neurophysiol. 2021, 38, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Sethi, N.K.; Neidecker, J. Neuroimaging in professional combat sports: Consensus statement from the association of ringside physicians. Phys. Sportsmed. 2023, 51, 343–350. [Google Scholar] [CrossRef]
- Burma, J.S.; Lapointe, A.P.; Wilson, M.; Penner, L.C.; Kennedy, C.M.; Newel, K.T.; Galea, O.A.; Miutz, L.N.; Dunn, J.F.; Smirl, J.D. Adolescent Sport-Related Concussion and the Associated Neurophysiological Changes: A Systematic Review. Pediatr. Neurol. 2024, 150, 97–106. [Google Scholar] [CrossRef]
- Kamali, A.M.; Ijadi, M.; Keshtkarhesamabadi, B.; Kazemiha, M.; Mahmoudi, R.; Roozbehi, A.; Nami, M. A dual-mode neurostimulation approach to enhance athletic performance outcome in experienced taekwondo practitioners. Sci. Rep. 2023, 13, 1748. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chmiel, J.; Nadobnik, J. Application of Electroencephalography (EEG) in Combat Sports—Review of Findings, Perspectives, and Limitations. J. Clin. Med. 2025, 14, 4113. https://doi.org/10.3390/jcm14124113
Chmiel J, Nadobnik J. Application of Electroencephalography (EEG) in Combat Sports—Review of Findings, Perspectives, and Limitations. Journal of Clinical Medicine. 2025; 14(12):4113. https://doi.org/10.3390/jcm14124113
Chicago/Turabian StyleChmiel, James, and Jarosław Nadobnik. 2025. "Application of Electroencephalography (EEG) in Combat Sports—Review of Findings, Perspectives, and Limitations" Journal of Clinical Medicine 14, no. 12: 4113. https://doi.org/10.3390/jcm14124113
APA StyleChmiel, J., & Nadobnik, J. (2025). Application of Electroencephalography (EEG) in Combat Sports—Review of Findings, Perspectives, and Limitations. Journal of Clinical Medicine, 14(12), 4113. https://doi.org/10.3390/jcm14124113