
Impact of Protein Citrullination by Periodontal Pathobionts on Oral and Systemic Health: A Systematic Review of Preclinical and Clinical Studies

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Protocol and Registration
2.2. Eligibility Criteria
2.3. Information and Search Sources
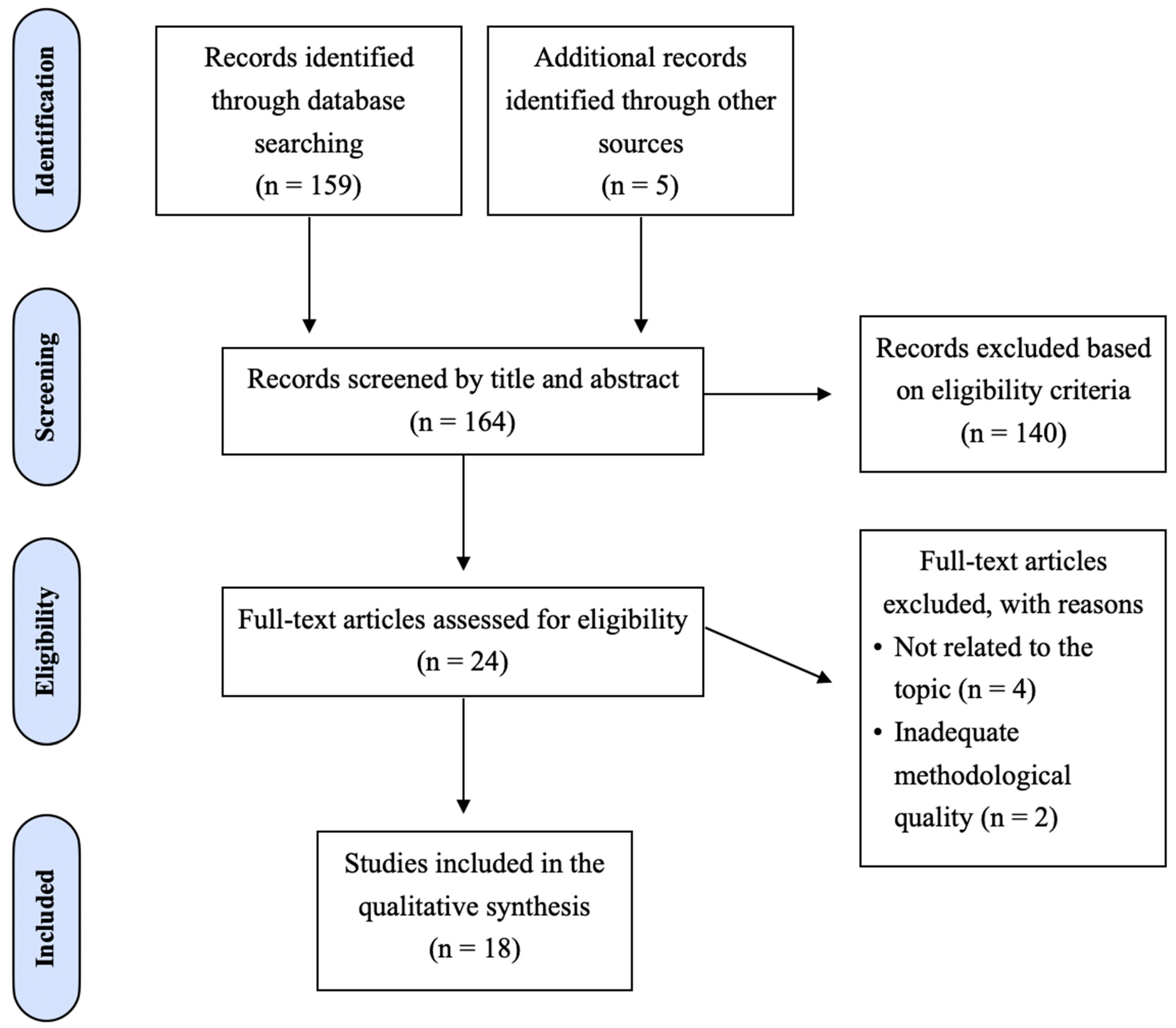
2.4. Selection of Studies
2.5. Data Extraction
2.6. Risk of Bias and Quality Assessment
3. Results
3.1. Risk of Bias and Quality Assessment
3.2. Study Selection
3.3. Study Characteristics
4. Discussion
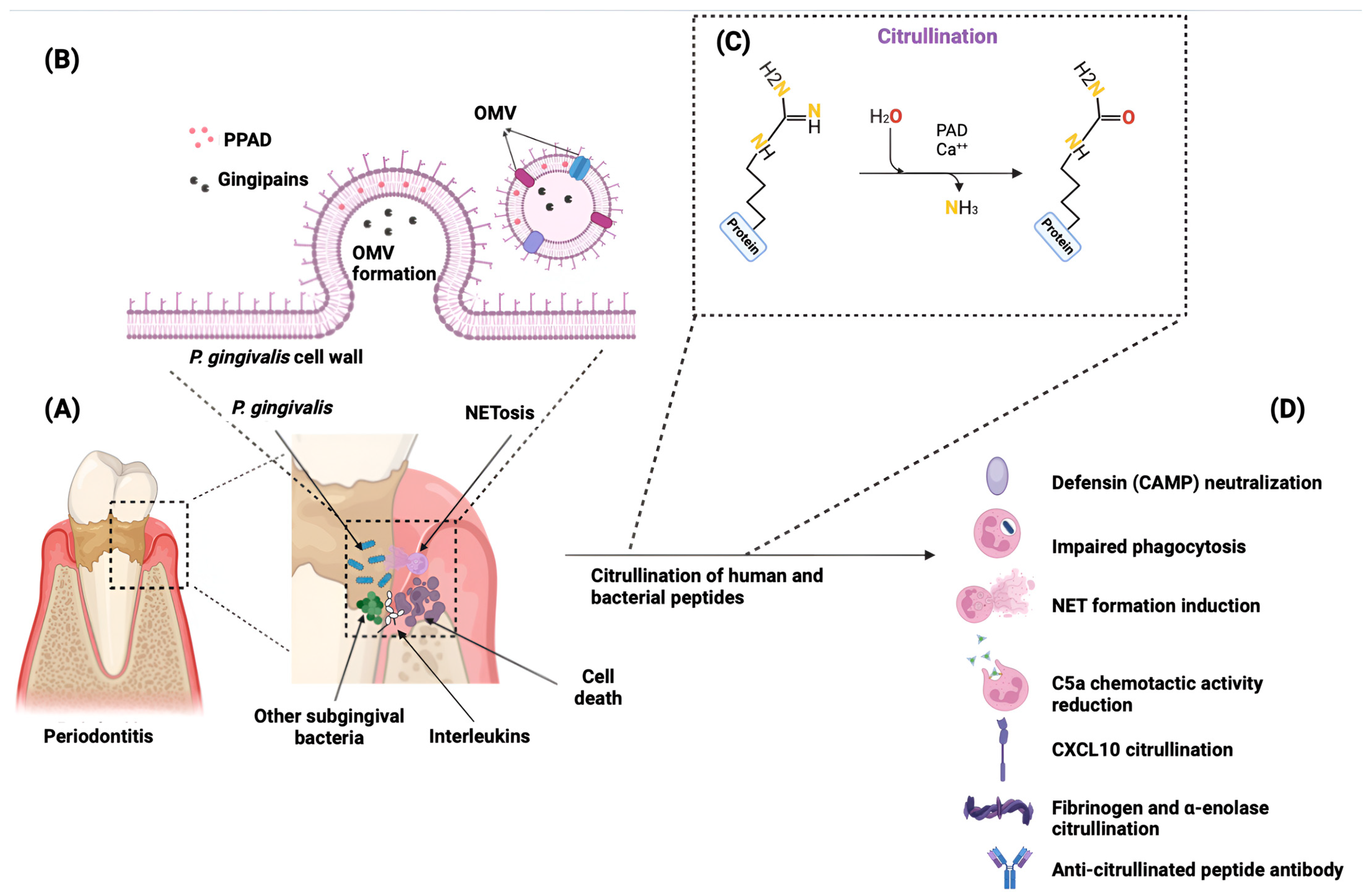
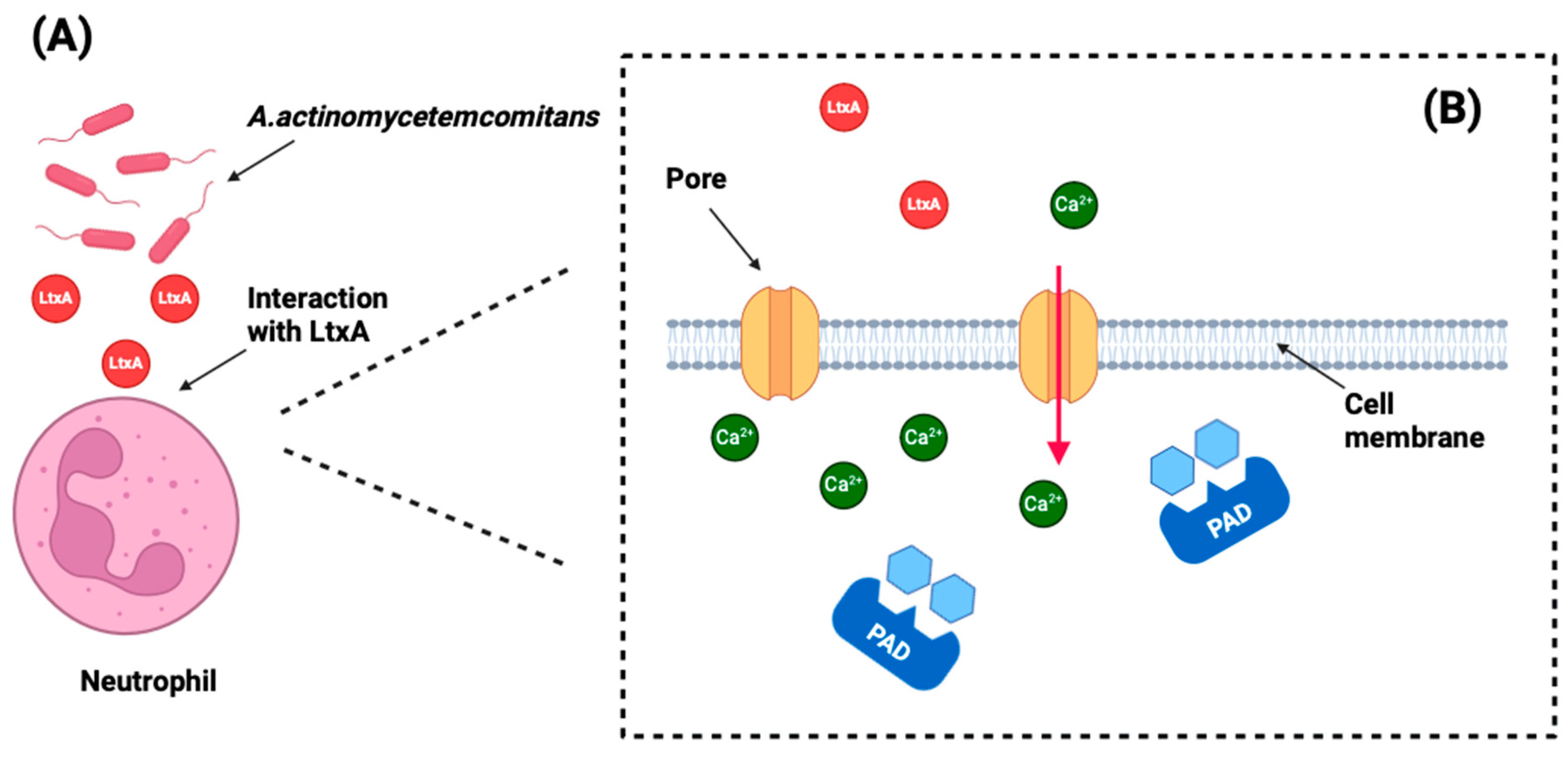
4.1. Impact on Systemic Health
4.2. Impact on Oral Health
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hajishengallis, G.; Chavakis, T. Local and Systemic Mechanisms Linking Periodontal Disease and Inflammatory Comorbidities. Nat. Rev. Immunol. 2021, 21, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Mesa, F.; Liebana, J.; Galindo-Moreno, P.; O’Valle, F. Oral Pathogens, Immunity, and Periodontal Diseases. Curr. Immunol. Rev. 2011, 7, 83–91. [Google Scholar] [CrossRef]
- Ray, R.R. Periodontitis: An Oral Disease with Severe Consequences. Appl. Biochem. Biotechnol. 2022, 195, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Trindade, D.; Carvalho, R.A.; Mendes, J.J.; Chambrone, L. Prevalence of Periodontitis in Dentate People Between 2011 and 2020: A Systematic Review and Meta-Analysis of Epidemiological Studies. J. Clin. Periodontol. 2023, 50, 604–626. [Google Scholar] [CrossRef] [PubMed]
- Curtis, M.A.; Diaz, P.I.; Van Dyke, T.E. The Role of the Microbiota in Periodontal Disease. Periodontology 2000 2020, 83, 14–25. [Google Scholar] [CrossRef]
- Horvat Aleksijević, L.; Aleksijević, M.; Škrlec, I.; Šram, M.; Talapko, J. Porphyromonas gingivalis Virulence Factors and Clinical Significance in Periodontal Disease and Coronary Artery Diseases. Pathogens 2022, 11, 1173. [Google Scholar] [CrossRef]
- Xu, W.; Zhou, W.; Wang, H.; Liang, S. Roles of Porphyromonas gingivalis and Its Virulence Factors in Periodontitis. Adv. Protein Chem. Struct. Biol. 2020, 120, 45–84. [Google Scholar]
- Di Stefano, M.; Polizzi, A.; Santonocito, S.; Romano, A.; Lombardi, T.; Isola, G. Impact of Oral Microbiome in Periodontal Health and Periodontitis: A Critical Review on Prevention and Treatment. Int. J. Mol. Sci. 2022, 23, 5142. [Google Scholar] [CrossRef]
- Hajishengallis, G. Interconnection of Periodontal Disease and Comorbidities: Evidence, Mechanisms, and Implications. Periodontology 2000 2022, 9, 9–18. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Chavakis, T.; Lambris, J.D. Current Understanding of Periodontal Disease Pathogenesis and Targets for Host-Modulation Therapy. Periodontology 2000 2020, 84, 14–34. [Google Scholar] [CrossRef]
- Kim, T.S.; Moutsopoulos, N.M. Neutrophils and Neutrophil Extracellular Traps in Oral Health and Disease. Exp. Mol. Med. 2024, 56, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Magán-Fernández, A.; Al-Bakri, S.M.R.; O’Valle, F.; Benavides-Reyes, C.; Abadía-Molina, F.; Mesa, F. Neutrophil Extracellular Traps in Periodontitis. Cells 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]
- Bryzek, D.; Ciaston, I.; Dobosz, E.; Gasiorek, A.; Makarska, A.; Sarna, M.; Eick, S.; Puklo, M.; Lech, M.; Potempa, B.; et al. Triggering NETosis via Protease-Activated Receptor (PAR)-2 Signaling as a Mechanism of Hijacking Neutrophil Function for Pathogen Benefits. PLoS Pathog. 2019, 15, e1007773. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tong, Y.; Zhu, Q.; Gao, L.; Sun, Y. Neutrophil Extracellular Traps Induced by Porphyromonas gingivalis Lipopolysaccharide Modulate Inflammatory Responses via a Ca2+-Dependent Pathway. Arch. Oral Biol. 2022, 141, 105467. [Google Scholar] [CrossRef]
- Rogers, G.E.; Simmonds, D.H. Content of Citrulline and Other Amino Acids in a Protein of Hair Follicles. Nature 1958, 182, 186–187. [Google Scholar] [CrossRef]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; Aljaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders. J. Immunol. Res. 2019, 2019, 7592851. [Google Scholar] [CrossRef]
- Gabarrini, G.; Heida, R.; van Ieperen, N.; Curtis, M.A.; van Winkelhoff, A.J.; van Dijl, J.M. Dropping Anchor: Attachment of Peptidylarginine Deiminase via A-LPS to Secreted Outer Membrane Vesicles of Porphyromonas gingivalis. Sci. Rep. 2018, 8, 8949. [Google Scholar] [CrossRef]
- Li, C.; Yu, R.; Ding, Y. Association between Porphyromonas gingivalis and Systemic Diseases: Focus on T Cells-Mediated Adaptive Immunity. Front. Cell. Infect. Microbiol. 2022, 12, 1026457. [Google Scholar] [CrossRef]
- Kurowska, W.; Kuca-Warnawin, E.H.; Radzikowska, A.; Maśliński, W. The Role of Anti-Citrullinated Protein Antibodies (ACPA) in the Pathogenesis of Rheumatoid Arthritis. Cent. Eur. J. Immunol. 2017, 42, 390–398. [Google Scholar] [CrossRef]
- Åberg, C.H.; Kelk, P.; Johansson, A. Aggregatibacter actinomycetemcomitans: Virulence of Its Leukotoxin and Association with Aggressive Periodontitis. Virulence 2014, 6, 188–195. [Google Scholar] [CrossRef]
- Svärd, A.; LoMartire, R.; Martinsson, K.; Öhman, C.; Kastbom, A.; Johansson, A. Presence and Immunoreactivity of Aggregatibacter actinomycetemcomitans in Rheumatoid Arthritis. Pathogens 2024, 13, 368. [Google Scholar] [CrossRef] [PubMed]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.A.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA Statement for Reporting Systematic Reviews and Meta-Analyses of Studies That Evaluate Health Care Interventions: Explanation and Elaboration. PLoS Med. 2009, 6, e1000100. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Prisma-P Group. Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols (PRISMA-P) 2015 Statement. Syst. Rev. 2015, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Handbook for Conducting a Literature-Based Health Assessment Using OHAT Approach for Systematic Review and Evidence Integration. 4 March 2019. Available online: https://ntp.niehs.nih.gov/ntp/ohat/pubs/handbookmarch2019_508.pdf (accessed on 5 August 2024).
- Sheth, V.H.; Shah, N.P.; Jain, R.; Bhanushali, N.; Bhatnagar, V. Development and Validation of a Risk-of-Bias Tool for Assessing In Vitro Studies Conducted in Dentistry: The QUIN. J. Prosthet. Dent. 2022, 131, 1038–1042. [Google Scholar] [CrossRef] [PubMed]
- Bereta, G.P.; Strzelec, K.; Łazarz-Bartyzel, K.; Dziedzic-Kowalska, A.; Nowakowska, Z.; Krutyhołowa, A.; Bielecka, E.; Kantyka, T.; Grabiec, A.M.; Kaczmarzyk, T.; et al. Identification of a New Genetic Variant (G231N, E232T, N235D) of Peptidylarginine Deiminase from P. gingivalis in Advanced Periodontitis. Front. Immunol. 2024, 15, 1355357. [Google Scholar] [CrossRef]
- Vermilyea, D.M.; Moradali, M.F.; Kim, H.M.; Davey, M.E. PPAD Activity Promotes Outer Membrane Vesicle Biogenesis and Surface Translocation by Porphyromonas gingivalis. J. Bacteriol. 2021, 203, e00343-20. [Google Scholar] [CrossRef]
- Larsen, D.N.; Mikkelsen, C.E.; Kierkegaard, M.; Bereta, G.; Nowakowska, Z.; Kaczmarek, J.Z.; Potempa, J.; Højrup, P. Citrullinome of Porphyromonas gingivalis Outer Membrane Vesicles: Confident Identification of Citrullinated Peptides. Mol. Cell. Proteomics 2020, 19, 167–180. [Google Scholar] [CrossRef]
- Aliko, A.; Kamińska, M.; Bergum, B.; Gawron, K.; Benedyk, M.; Lamont, R.J.; Malicki, S.; Delaleu, N.; Potempa, J.; Mydel, P. Impact of Porphyromonas gingivalis Peptidylarginine Deiminase on Bacterial Biofilm Formation, Epithelial Cell Invasion, and Epithelial Cell Transcriptional Landscape. Sci. Rep. 2018, 8, 14144. [Google Scholar] [CrossRef]
- Hamamoto, Y.; Ouhara, K.; Munenaga, S.; Shoji, M.; Ozawa, T.; Hisatsune, J.; Kado, I.; Kajiya, M.; Matsuda, S.; Kawai, T.; et al. Effect of Porphyromonas gingivalis Infection on Gut Dysbiosis and Resultant Arthritis Exacerbation in Mouse Model. Arthritis Res. Ther. 2020, 22, 249. [Google Scholar] [CrossRef]
- Vermilyea, D.M.; Ottenberg, G.K.; Davey, M.E. Citrullination Mediated by PPAD Constrains Biofilm Formation in P. gingivalis Strain 381. Npj Biofilms Microbiomes 2019, 5, 7. [Google Scholar] [CrossRef]
- Stobernack, T.; du Teil Espina, M.; Mulder, L.M.; Palma Medina, L.M.; Piebenga, D.R.; Gabarrini, G.; Zhao, X.; Janssen, K.M.; Hulzebos, J.; Brouwer, E.; et al. A Secreted Bacterial Peptidylarginine Deiminase Can Neutralize Human Innate Immune Defenses. mBio 2018, 9, e01704-18. [Google Scholar] [CrossRef] [PubMed]
- Engström, M.; Eriksson, K.; Lee, L.; Hermansson, M.; Johansson, A.; Nicholas, A.P.; Gerasimcik, N.; Lundberg, K.; Klareskog, L.; Catrina, A.I.; et al. Increased Citrullination and Expression of Peptidylarginine Deiminases Independently of P. gingivalis and A. actinomycetemcomitans in Gingival Tissue of Patients with Periodontitis. J. Transl. Med. 2018, 16, 214. [Google Scholar] [CrossRef] [PubMed]
- Elkaim, R.; Bugueno-Valdebenito, I.M.; Benkirane-Jessel, N.; Tenenbaum, H. Porphyromonas gingivalis and Its LPS Differentially Regulate the Expression of Peptidyl Arginine Deiminases in Human Chondrocytes. Innate Immun. 2017, 23, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Konig, M.F.; Abusleme, L.; Reinholdt, J.; Palmer, R.J.; Teles, R.P.; Sampson, K.; Rosen, A.; Nigrovic, P.A.; Sokolove, J.; Giles, J.T.; et al. Aggregatibacter actinomycetemcomitans-Induced Hypercitrullination Links Periodontal Infection to Autoimmunity in Rheumatoid Arthritis. Sci. Transl. Med. 2016, 8, 369ra176. [Google Scholar] [CrossRef] [PubMed]
- Stobernack, T.; Glasner, C.; Junker, S.; Gabarrini, G.; de Smit, M.; de Jong, A.; Otto, A.; Becher, D.; van Winkelhoff, A.J.; van Dijl, J.M. Extracellular Proteome and Citrullinome of the Oral Pathogen Porphyromonas gingivalis. J. Proteome Res. 2016, 15, 4532–4543. [Google Scholar] [CrossRef]
- Montgomery, A.B.; Kopec, J.; Shrestha, L.; Thezenas, M.L.; Burgess-Brown, N.A.; Fischer, R.; Yue, W.W.; Venables, P.J. Crystal Structure of Porphyromonas gingivalis Peptidylarginine Deiminase: Implications for Autoimmunity in Rheumatoid Arthritis. Ann. Rheum. Dis. 2015, 75, 1255–1261. [Google Scholar] [CrossRef]
- Reichert, S.; Schlumberger, W.; Dähnrich, C.; Hornig, N.; Altermann, W.; Schaller, H.G.; Schulz, S. Association of Levels of Antibodies Against Citrullinated Cyclic Peptides and Citrullinated α-Enolase in Chronic and Aggressive Periodontitis as a Risk Factor of Rheumatoid Arthritis: A Case-Control Study. J. Transl. Med. 2015, 13, 283. [Google Scholar] [CrossRef]
- Bielecka, E.; Scavenius, C.; Kantyka, T.; Jusko, M.; Mizgalska, D.; Szmigielski, B.; Potempa, B.; Enghild, J.J.; Prossnitz, E.R.; Blom, A.M.; et al. Peptidyl Arginine Deiminase from Porphyromonas gingivalis Abolishes Anaphylatoxin C5a Activity. J. Biol. Chem. 2014, 289, 32481–32487. [Google Scholar] [CrossRef]
- Moelants, E.A.V.; Loozen, G.; Mortier, A.; Martens, E.; Opdenakker, G.; Mizgalska, D.; Szmigielski, B.; Potempa, J.; Van Damme, J.; Teughels, W.; et al. Citrullination and Proteolytic Processing of Chemokines by Porphyromonas gingivalis. Infect. Immun. 2014, 82, 2511–2519. [Google Scholar] [CrossRef]
- Abdullah, S.N.; Farmer, E.A.; Spargo, L.; Logan, R.; Gully, N. Porphyromonas gingivalis Peptidylarginine Deiminase Substrate Specificity. Anaerobe 2013, 23, 102–108. [Google Scholar] [CrossRef]
- Maresz, K.J.; Hellvard, A.; Sroka, A.; Adamowicz, K.; Bielecka, E.; Koziel, J.; Gawron, K.; Mizgalska, D.; Marcinska, K.A.; Benedyk, M.; et al. Porphyromonas gingivalis Facilitates the Development and Progression of Destructive Arthritis through Its Unique Bacterial Peptidylarginine Deiminase (PAD). PLoS Pathog. 2013, 9, e1003627. [Google Scholar] [CrossRef] [PubMed]
- Wegner, N.; Wait, R.; Sroka, A.; Eick, S.; Nguyen, K.A.; Lundberg, K.; Kinloch, A.; Culshaw, S.; Potempa, J.; Venables, P.J. Peptidylarginine Deiminase from Porphyromonas gingivalis Citrullinates Human Fibrinogen and α-Enolase: Implications for Autoimmunity in Rheumatoid Arthritis. Arthritis Rheum. 2010, 62, 2662–2672. [Google Scholar] [CrossRef] [PubMed]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial Complexes in Subgingival Plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Darveau, R.P.; Curtis, M.A. The Keystone-Pathogen Hypothesis. Nat. Rev. Microbiol. 2012, 10, 717–725. [Google Scholar] [CrossRef]
- Kaplan, A.H.; Weber, D.J.; Oddone, E.Z.; Perfect, J.R. Infection Due to Actinobacillus actinomycetemcomitans: 15 Cases and Review. Rev. Infect. Dis. 1989, 11, 46–63. [Google Scholar] [CrossRef]
- Fine, D.H.; Markowitz, K.; Fairlie, K.; Tischio-Bereski, D.; Ferrendiz, J.; Furgang, D.; Paster, B.J.; Dewhirst, F.E. A Consortium of Aggregatibacter actinomycetemcomitans, Streptococcus parasanguinis, and Filifactor alocis Is Present in Sites Prior to Bone Loss in a Longitudinal Study of Localized Aggressive Periodontitis. J. Clin. Microbiol. 2013, 51, 2850–2861. [Google Scholar] [CrossRef]
- de Jong, C.; Bikker, F.J.; Gibbs, S.; Krom, B.P. Mechanisms of Porphyromonas gingivalis to Translocate over the Oral Mucosa and Other Tissue Barriers. J. Oral Microbiol. 2023, 15, 2205291. [Google Scholar] [CrossRef]
- Ellis, T.N.; Kuehn, M.J. Virulence and Immunomodulatory Roles of Bacterial Outer Membrane Vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef]
- Zhou, L.; Srisatjaluk, R.; Justus, D.E.; Doyle, R.J. On the Origin of Membrane Vesicles in Gram-Negative Bacteria. FEMS Microbiol. Lett. 1998, 163, 223–228. [Google Scholar] [CrossRef]
- Mantri, C.K.; Chen, C.H.; Dong, X.; Goodwin, J.S.; Pratap, S.; Paromov, V.; Xie, H. Fimbriae-Mediated Outer Membrane Vesicle Production and Invasion of Porphyromonas gingivalis. Microbiol. Open 2014, 4, 53–65. [Google Scholar] [CrossRef]
- Leliefeld, P.H.C.; Wessels, C.M.; Leenen, L.P.H.; Koenderman, L.; Pillay, J. The Role of Neutrophils in Immune Dysfunction during Severe Inflammation. Crit. Care 2016, 20, 73. [Google Scholar] [CrossRef] [PubMed]
- Li, W. Phagocyte Dysfunction, Tissue Aging and Degeneration. Ageing Res. Rev. 2013, 12, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Pan, B.; Alam, H.B.; Liang, Y.; Wu, Z.; Liu, B.; Mor-Vaknin, N.; Duan, X.; Williams, A.M.; Tian, Y.; et al. Citrullinated Histone H3 as a Therapeutic Target for Endotoxic Shock in Mice. Front. Immunol. 2020, 10, 2957. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Alam, H.B.; Chong, W.; Mobley, J.; Liu, B.; Deng, Q.; Liang, Y.; Wang, Y.; Chen, E.; Wang, T.; et al. CitH3: A Reliable Blood Biomarker for Diagnosis and Treatment of Endotoxic Shock. Sci. Rep. 2017, 7, 8972. [Google Scholar] [CrossRef] [PubMed]
- Sorvillo, N.; Cherpokova, D.; Martinod, K.; Wagner, D.D. Extracellular DNA NET-Works with Dire Consequences for Health. Circ. Res. 2019, 125, 470–488. [Google Scholar] [CrossRef]
- O’Donoghue, E.J.; Krachler, A.M. Mechanisms of Outer Membrane Vesicle Entry into Host Cells. Cell Microbiol. 2016, 18, 1508–1517. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The Roles of Antimicrobial Peptides in Innate Host Defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef]
- Duarte-Mata, D.I.; Salinas-Carmona, M.C. Antimicrobial Peptides’ Immune Modulation Role in Intracellular Bacterial Infection. Front. Immunol. 2023, 14, 1119574. [Google Scholar] [CrossRef]
- Al Adwani, S.; Padhi, A.; Karadottir, H.; Mörman, C.; Gräslund, A.; Végvári, Á.; Johansson, J.; Rising, A.; Agerberth, B.; Bergman, P. Citrullination Alters the Antibacterial and Anti-Inflammatory Functions of the Host Defense Peptide Canine Cathelicidin K9CATH In Vitro. J. Immunol. 2021, 207, 974–984. [Google Scholar] [CrossRef]
- Vazirinejad, R.; Ahmadi, Z.; Kazemi Arababadi, M.; Hassanshahi, G.; Kennedy, D. The Biological Functions, Structure, and Sources of CXCL10 and Its Outstanding Role in the Pathophysiology of Multiple Sclerosis. Neuroimmunomodulation 2014, 21, 322–330. [Google Scholar] [CrossRef]
- Loos, T.; Mortier, A.; Gouwy, M.; Ronsse, I.; Put, W.; Lenaerts, J.P.; Van Damme, J.; Proost, P. Citrullination of CXCL10 and CXCL11 by Peptidylarginine Deiminase: A Naturally Occurring Posttranslational Modification of Chemokines and New Dimension of Immunoregulation. Blood 2008, 112, 2648–2656. [Google Scholar] [CrossRef] [PubMed]
- Bickel, M. The Role of Interleukin-8 in Inflammation and Mechanisms of Regulation. J. Periodontol. 1993, 64 (Suppl. 5), 456–460. [Google Scholar] [PubMed]
- Vermeersch, G.; Proost, P.; Struyf, S.; Gouwy, M.; Devos, T. CXCL8 and Its Cognate Receptors CXCR1/CXCR2 in Primary Myelofibrosis. Haematologica 2024, 109, 2060. [Google Scholar] [CrossRef] [PubMed]
- Proost, P.; Loos, T.; Mortier, A.; Schutyser, E.; Gouwy, M.; Noppen, S.; Dillen, C.; Ronsse, I.; Conings, R.; Struyf, S.; et al. Citrullination of CXCL8 by Peptidylarginine Deiminase Alters Receptor Usage, Prevents Proteolysis, and Dampens Tissue Inflammation. J. Exp. Med. 2008, 205, 2085–2097. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Gao, R.; Chen, H.; Hu, J.; Zhang, P.; Wei, X.; Shi, J.; Chen, Y.; Zhang, L.; Chen, J.; et al. Myocardial Reperfusion Injury Exacerbation due to ALDH2 Deficiency Is Mediated by Neutrophil Extracellular Traps and Prevented by Leukotriene C4 Inhibition. Eur. Heart J. 2024, 45, 1662–1680. [Google Scholar] [CrossRef]
- Savchenko, A.S.; Borissoff, J.I.; Martinod, K.; De Meyer, S.F.; Gallant, M.; Erpenbeck, L.; Brill, A.; Wang, Y.; Wagner, D.D. VWF-Mediated Leukocyte Recruitment with Chromatin Decondensation by PAD4 Increases Myocardial Ischemia/Reperfusion Injury in Mice. Blood 2014, 123, 141–148. [Google Scholar] [CrossRef]
- Meegan, J.E.; Yang, X.; Beard, R.S., Jr.; Jannaway, M.; Chatterjee, V.; Taylor-Clark, T.E.; Yuan, S.Y. Citrullinated Histone 3 Causes Endothelial Barrier Dysfunction. Biochem. Biophys. Res. Commun. 2018, 503, 1498–1502. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Question Author | Was Administered Dose or Exposure Level Adequately Randomized? | Was Allocation to Study Groups Adequately Concealed? | Were Experimental Conditions Identical Across Study Groups? | Were Research Personnel Blinded to the Study Group During the Study? | Were Outcome Data Complete Without Attrition or Exclusion from the Analysis? | Can We Be Confident in the Exposure Characterization? | Can We Be Confident in the Outcome Assessment (Including Blinding of Assessors)? | Were There No Other Potential Threats to Internal Validity? |
|---|---|---|---|---|---|---|---|---|
| Bereta GP et al. (2024) [26] | − | ++ | ++ | − | + | + | − | + |
| Vermilyea DM et al. (2021) [27] | − | ++ | ++ | − | ++ | + | − | + |
| Larsen DN et al. (2020) [28] | − | ++ | ++ | − | + | + | − | + |
| Aliko A et al. (2020) [29] | − | ++ | ++ | − | ++ | + | − | + |
| Hamamoto Y et al. (2020) [30] | − | ++ | + | − | ++ | + | − | + |
| Vermilyea DM et al. (2019) [31] | − | ++ | ++ | − | ++ | + | − | + |
| Stobernack T et al. (2018) [32] | − | ++ | ++ | − | ++ | + | − | + |
| Engström M et al. (2018) [33] | − | ++ | ++ | − | ++ | + | − | + |
| Elkaim R et al. (2017) [34] | − | ++ | ++ | − | ++ | + | − | + |
| Konig MF et al. (2016) [35] | − | ++ | ++ | − | + | + | − | + |
| Stobernack et al. (2016) [36] | − | ++ | ++ | − | + | + | − | + |
| Montgomery AB et al. (2016) [37] | − | ++ | ++ | − | + | + | − | + |
| Reichert S et al. (2015) [38] | − | ++ | ++ | − | ++ | + | − | + |
| Bielecka E et al. (2014) [39] | − | ++ | ++ | − | ++ | + | − | + |
| Moelants EAV et al. (2014) [40] | − | ++ | + | − | + | + | − | + |
| Abdullah SN et al. (2013) [41] | − | ++ | ++ | − | ++ | + | − | + |
| Maresz K et al. (2013) [42] | − | ++ | ++ | ++ | + | + | − | − |
| Wegner N et al. (2011) [43] | − | ++ | + | − | ++ | + | − | + |
| Quality Assessment Tool for In Vitro Studies | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clearly Stated Aims | Detailed Explanation of Sample Size Calculation | Detailed Explanation of Sampling Technique | Details of Comparison Group | Detailed Explanation of Methodology | Operator Details | Randomization | Method of Measurement Outcome | Outcome Assessor Details | Blinding | Statistical Analysis | Presentation of Results | Score (%) | |
| Bereta GP et al. (2024) [26] | ++ | − | + | ++ | ++ | − | − | ++ | + | − | + | + | 50 |
| Vermilyea DM et al. (2021) [27] | + | NA | ++ | ++ | ++ | − | − | ++ | + | NA | + | ++ | 65 |
| Larsen DN et al. (2020) [28] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | NA | ++ | 83 |
| Aliko A et al. (2020) [29] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | ++ | ++ | 75 |
| Hamamoto Y et al. (2020) [30] | + | ++ | ++ | ++ | ++ | − | − | ++ | + | − | ++ | ++ | 75 |
| Vermilyea DM et al. (2019) [31] | − | NA | ++ | ++ | ++ | − | − | ++ | + | NA | NA | ++ | 61 |
| Stobernack T et al. (2018) [32] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | ++ | ++ | 75 |
| Engström M et al. (2018) [33] | ++ | − | + | ++ | ++ | − | − | ++ | + | ++ | ++ | ++ | 67 |
| Elkaim R et al. (2017) [34] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | + | ++ | 70 |
| Konig MF et al. (2016) [35] | ++ | − | − | ++ | − | − | − | ++ | + | − | + | ++ | 42 |
| Stobernack et al. (2016) [36] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | ++ | ++ | 75 |
| Montgomery AB et al. (2016) [37] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | NA | ++ | 72 |
| Reichert S et al. (2015) [38] | ++ | − | + | ++ | ++ | − | − | ++ | + | − | ++ | ++ | 58 |
| Bielecka E et al. (2014) [39] | + | NA | ++ | ++ | ++ | − | − | ++ | + | NA | ++ | ++ | 70 |
| Moelants EAV et al. (2014) [40] | − | − | + | NA | ++ | − | − | ++ | + | − | + | + | 40 |
| Abdullah SN et al. (2013) [41] | ++ | NA | ++ | ++ | ++ | − | − | ++ | + | NA | NA | + | 67 |
| Maresz K et al. (2013) [42] | − | − | + | ++ | ++ | − | − | + | + | + | ++ | ++ | 50 |
| Wegner N et al. (2011) [43] | ++ | − | ++ | NA | ++ | − | − | ++ | + | − | NA | ++ | 55 |
| Author/Year | Strain | Bacteria Provenance | Control Group (Non Citrullinating) | Citrullination Ability | Main Techniques Employed | Periodontal Effects | Systemic Effects |
|---|---|---|---|---|---|---|---|
| Clinical Studies on P. gingivalis | |||||||
| Bereta GP et al. (2024) [26] | Not specified | Clinical isolates | P. gingivalis W83 and ATCC 22377 | Yes | PCR, Western Blot, HPLC, RNA-seq | Advanced chronic periodontitis showed more citrullinating activity. CAL and PPD values correlate with the presence of P. gingivalis strain harboring a triple polymorphic variant (G231N, E232T, N235D) P. gingivalis strains with higher PPAD activity were found more frequently in subjects with moderate/advanced and advanced CP | Not applicable |
| Stobernack et al. (2016) [36] | P. gingivalis ATCC 33277 and W83 and three isolates: 20658, MDS16, MDS45 | American Type Culture Collection and clinical isolates (CP-RA) | P. Gingivalis W83 and TCC 33277Δppad | Yes | Mass spectrometry, exoproteome analysis | The RA-associated isolates MDS16 and MDS45 showed unique citrullinated proteins, such as the Mfa1 fimbrilin. Gingipains and other key virulence factors like RgpA were citrullinated in specific isolates like W83 and MDS45, but not in PPAD-deficient mutants or other strains | Not applicable |
| Reichert S et al. (2015) [38] | Not specified | Clinical isolates | Non-RA non-Periodontitis patients | Yes | ELISA, HLA typing, PCR | Anti- α-enolase antibodies were slightly elevated in the periodontitis group. | Not applicable |
| Moelants EAV et al. (2014) [40] | P. gingivalis ATCC 33277, 5 clinical isolates (P. gingivalis to P. gingivalis5) | Research laboratory and clinical isolates | Protease inhibitors, Five capsule-typed strains (K1 to K5) | Yes | Enzymatic assay, ELISA | Not applicable | No citrullination was detected in CCL3, which lacks NH2-terminal Arg and hence could not be sequenced for citrullination. CXCL8 incubated with different strains of P. gingivalis showed minimal citrullination (up to 5%). CXCL10 levels rapidly decreased, indicating rapid degradation by P. gingivalis |
| Wegner N et al. (2011) [43] | P. gingivalis W83, 4 clinical isolates (MaRL, D243, JH16, J430) | Research laboratory and clinical isolates (CP) | P. gingivalis mutants (Δppad, ppad+, Δrgp, Δkgp, Δrgp+kgp), Prevotella intermedia H13 (clinical isolate), Prevotella oralis ATCC 33269, Capnocytophaga gingivalis ATCC 33624, Capnocytophaga ochracea ATCC 27872 | Yes | Immunoblot, HPLC, mass spectrometry | P. gingivalis expresses endogenous citrullinated proteins, a feature absent in other oral bacteria tested. The presence of citrullinated proteins in P. gingivalis is entirely dependent on the PPAD. | Arginine-gingipains play a crucial role in generating substrates for PPAD by cleaving proteins at arginine residues. P. gingivalis can citrullinate human proteins such as fibrinogen and α-enolase after cleavage by arginine-gingipains |
| Clinical Studies on A. actinomycetemcomitans | |||||||
| Engström M et al. (2018) [33] | Not specified | Clinical isolates from periodontal patients | Healthy controls | Yes | Immunohistochemistry, qPCR | Citrullinated proteins were found predominantly in gingival tissues of periodontitis patients (80%) versus healthy controls (27%). No significant differences in citrullination were noted between the epithelial compartments of periodontitis and healthy tissues. PAD2 and PAD4 expressed higher levels in periodontitis. | Not applicable |
| Konig MF et al. (2016) [35] | Not specified | Clinical isolates from RA patients | Healthy controls | Yes | PCR, ELISA, mass spectrometry | Not applicable | LtxA induces hypercitrullination in neutrohpils by creating pores in the cell membrane, allowing uncontrolled calcium influx into the cell. |
| In Vivo Studies on Laboratory Animals on P. gingivalis | |||||||
| Hamamoto Y et al. (2020) [30] | P. gingivalis W83 and 33277 | American Type Culture Collection | PPAD knockout | Yes | ELISA, pirosequencing | FMT from P. gingivalis-inoculated mice exacerbated joint destruction and increased IL-6 and CP levels in recipient mice. P. gingivalis oral infection led to severe joint destruction, elevated arthritis scores (AS), and increased production of IL-6 and citrullinated peptides (CP) in serum, joint, and intestinal tissues. | |
| Maresz K et al. (2013) [42] | P. gingivalis W83 | Not specified | PPAD knockout, Prevotella Intermedia and control mice not inoculated with P. gingivalis | Yes | PCR, immunoscan, MPO determination | Not applicable | Mice infected with live P. gingivalis showed earlier onset and more severe arthritis compared to control mice. Higher MPO activity, indicating more neutrophil infiltration, was observed in joints of P. gingivalis-infected mice. Higher levels of antibodies against citrullinated α-enolase were found in the serum of mice infected with live P. gingivalis. |
| In Vitro Studies on P. gingivalis | |||||||
| Vermilyea DM et al. (2021) [27] | P. gingivalis 381 | H. Kuramitsu, State University of Buffalo, Buffalo, NY and American Type Culture Collection | P. gingivalis 381 Δppad | Yes | Enzymatic and biofilm assay, quantification of OMVs, RNA-seq, PCR | PPAD plays a crucial role in regulating biofilm dynamics, specifically by facilitating surface translocation and reducing biofilm formation. PPAD plays a key role in regulating sessile lifestyle towards surface translocation. | P. gingivalis 381 Δppad accumulates more intracellular arginine (lacks PPAD activity) and creates fewer and smaller OMVs. |
| Larsen DN et al. (2020) [28] | P. gingivalis W83 | Not specified | P. gingivalis W83 Δppad and C351A (cysteine mutated to alanine) | Yes | HPLC separation, mass spectrometry, aminoacid assay | P. gingivalis W83 exhibited more citrullination compared to the mutant strain. | Citrullination in OMVs is dependent on functional PPAD, no citrullinated proteins were identified in the ΔPPAD strain and only one in the PPADC351A mutant. |
| Aliko A et al. (2020) [29] | P. gingivalis WT 33277 | American Type Culture Collection | P. gingivalis 33277 Δppad and PPADC351A (inactive) | Yes | PCR, flow cytometry, SEM, RNA-Seq | PPAD activity does not affect the ability of P. gingivalis to adhere to or be internalized by human oral keratinocytes. | The gene expression analysis revealed that PPAD activity in P. gingivalis specifically affects immune response pathways in oral keratynocites, notably impacting IL-1 signaling and immune cell chemotaxis. Key genes such as CXCL8, IL36G, CCL20, and IL1B showed significant PPAD-dependent expression changes, underscoring PPAD’s role in modulating immune responses. |
| Vermilyea DM et al. (2019) [31] | P. gingivalis 381 | State University of Buffalo, Buffalo, NY | P. gingivalis Δ8820 | Yes | TEM, cryo-SEM, enzymatic assay, immunoblot | Δppad in P. gingivalis enhances biofilm formation. PPAD inhibits biofilm formation by regulating matrix production. This enhanced biofilm formation is not due to increased fimbriae expression but is linked to the absence of citrullination in key proteins like RgpA and Kgp. Δ8820 biofilms contained more gingipain-derived adhesin proteins and more matrix. | PPAD impacts growth, colonization, attachment, and invasion of host cells and tissues. |
| Stobernack T et al. (2018) [32] | P. gingivalis W83 | American Type Culture Collection and clinical isolates (CP-RA) | P. gingivalis W83 Δppad | Yes | Immunohistochemistry, phagocytosis assay, flow cytometry, mass spectrometry | Not applicable | The presence of PPAD significantly impairs the binding and internalization of P. gingivalis by neutrophils. PPAD interferes with the phagocytosis process by citrullinating gingipains, which are involved in modulating actin polymerization and phagocytosis. PPAD citrulliantes histone H3, impairing NETosis, it also citrullinates LP9 and therefore neutralizes CAMPs. |
| Elkaim R et al. (2017) [34] | P, gingivalis 33277 | American Type Culture Collection | Uninfected human chondrocytes | Yes | qPCR, immunoblot, enzymatic assay | Not applicable | P. gingivalis can suppress the expression and activity of human PADs while enhancing the activity of its own PAD enzyme. Infection with live P. gingivalis resulted in new citrullinated peptide bands (15 kDa to 45 kDa) in cellular extracts. |
| Montgomery AB et al. (2016) [37] | P. gingivalis W83 | Not specified | P. gingivalis tPPADC351A, tPPADR152A, tPPADR154A, | Yes | Enzymatic assay, mass spectormetry | Not applicable | The Cys351Ala mutation rendered the enzyme catalytically inactive, Cys351 plays a key role in PPAD’s enzymatic function. PPAD specifically citrullinates C-terminal arginine residues, distinguishing its substrate specificity from human PAD2 and PAD4, which prefer internal and N-terminal arginine residues. tPPADWT citrullinated peptides derived from RA autoantigens, such as fibrinogen and α-enolase. |
| Bielecka E et al. (2014) [39] | P. gingivalis W83 | Not specified | P. gingivalis W83 Δppad | Yes | Isolation of OMV, HPLC, mass spectrometry, calcium mobilization assay | Not applicable | PPAD citrullinates C5a. OMVs from P. gingivalis W83 efficiently citrullinate C5a, thus indicating that OMVs carry both Arg-specific gingipains and PPAD. C5a-Cit showed significantly reduced chemotactic activity for neutrophils. At high concentrations and/or prolonged incubation with PPAD, C5a capacity to activate neutrophils is abrogated. |
| Abdullah SN et al. (2013) [41] | P. gingivalis W50 | Research laboratory | Protease inhibitors | Yes | Gingipains inhibition, azocasein assay, colorimetric assay, protein assay | Optimal activity was observed between pH 7.5 and 8, with significant activity retained at pH 9. PPAD exhibited 87% of its activity in heat-treated cells, indicating heat stability. Vimentin showed the highest rate of citrullination. | Gingipain inhibition did not affect PPAD activity. PPAD and gingipains function independently in terms of their enzymatic activities. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonilla, M.; Martín-Morales, N.; Gálvez-Rueda, R.; Raya-Álvarez, E.; Mesa, F. Impact of Protein Citrullination by Periodontal Pathobionts on Oral and Systemic Health: A Systematic Review of Preclinical and Clinical Studies. J. Clin. Med. 2024, 13, 6831. https://doi.org/10.3390/jcm13226831
Bonilla M, Martín-Morales N, Gálvez-Rueda R, Raya-Álvarez E, Mesa F. Impact of Protein Citrullination by Periodontal Pathobionts on Oral and Systemic Health: A Systematic Review of Preclinical and Clinical Studies. Journal of Clinical Medicine. 2024; 13(22):6831. https://doi.org/10.3390/jcm13226831
Chicago/Turabian StyleBonilla, Marco, Natividad Martín-Morales, Rocío Gálvez-Rueda, Enrique Raya-Álvarez, and Francisco Mesa. 2024. "Impact of Protein Citrullination by Periodontal Pathobionts on Oral and Systemic Health: A Systematic Review of Preclinical and Clinical Studies" Journal of Clinical Medicine 13, no. 22: 6831. https://doi.org/10.3390/jcm13226831
APA StyleBonilla, M., Martín-Morales, N., Gálvez-Rueda, R., Raya-Álvarez, E., & Mesa, F. (2024). Impact of Protein Citrullination by Periodontal Pathobionts on Oral and Systemic Health: A Systematic Review of Preclinical and Clinical Studies. Journal of Clinical Medicine, 13(22), 6831. https://doi.org/10.3390/jcm13226831