

Further Association of Germline CHEK2 Loss-of-Function Variants with Testicular Germ Cell Tumors

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Genetic Analyses
2.3. Statistical Analyses
3. Results
3.1. Clinical Characteristics of the Studied Cases
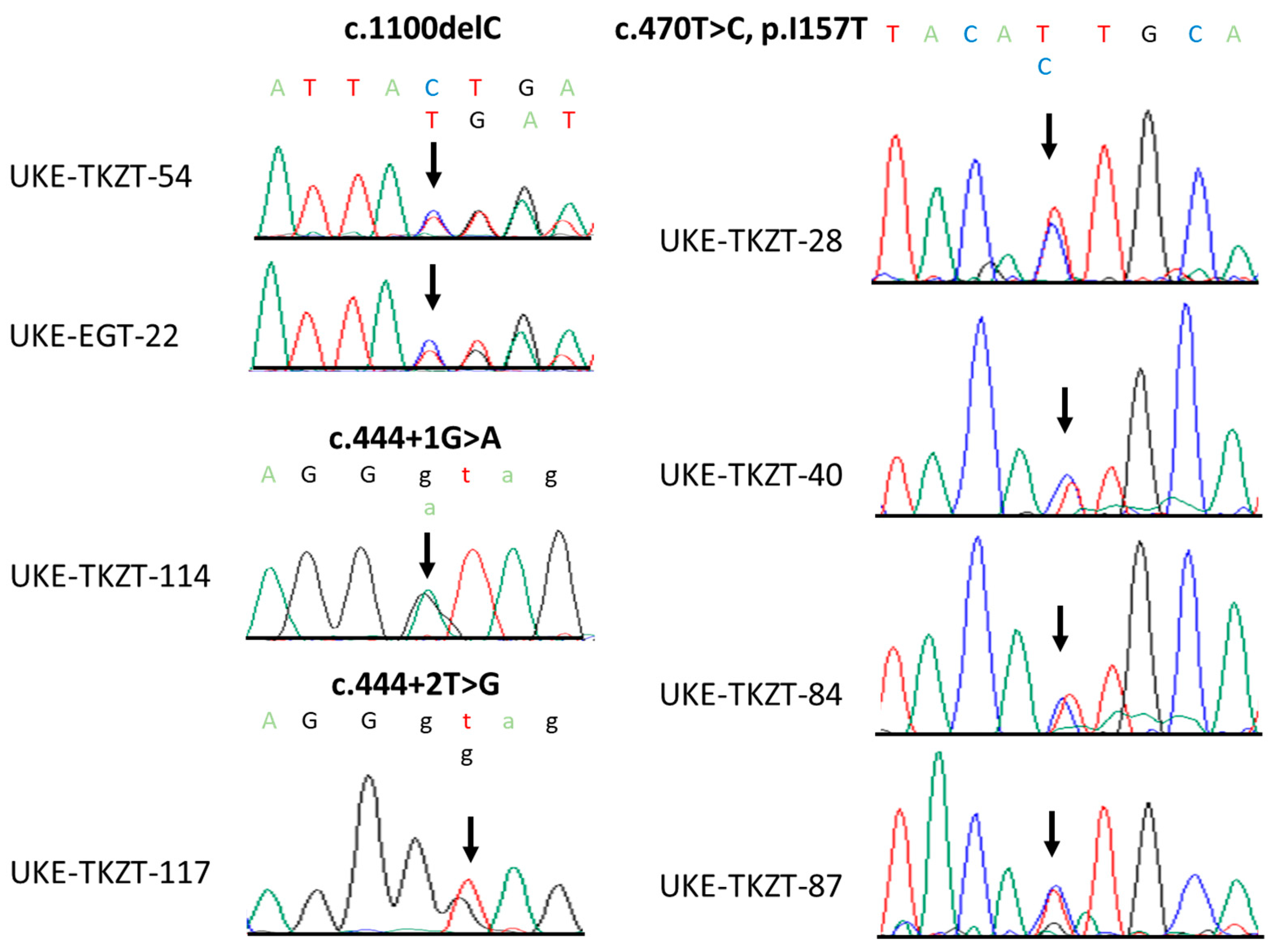
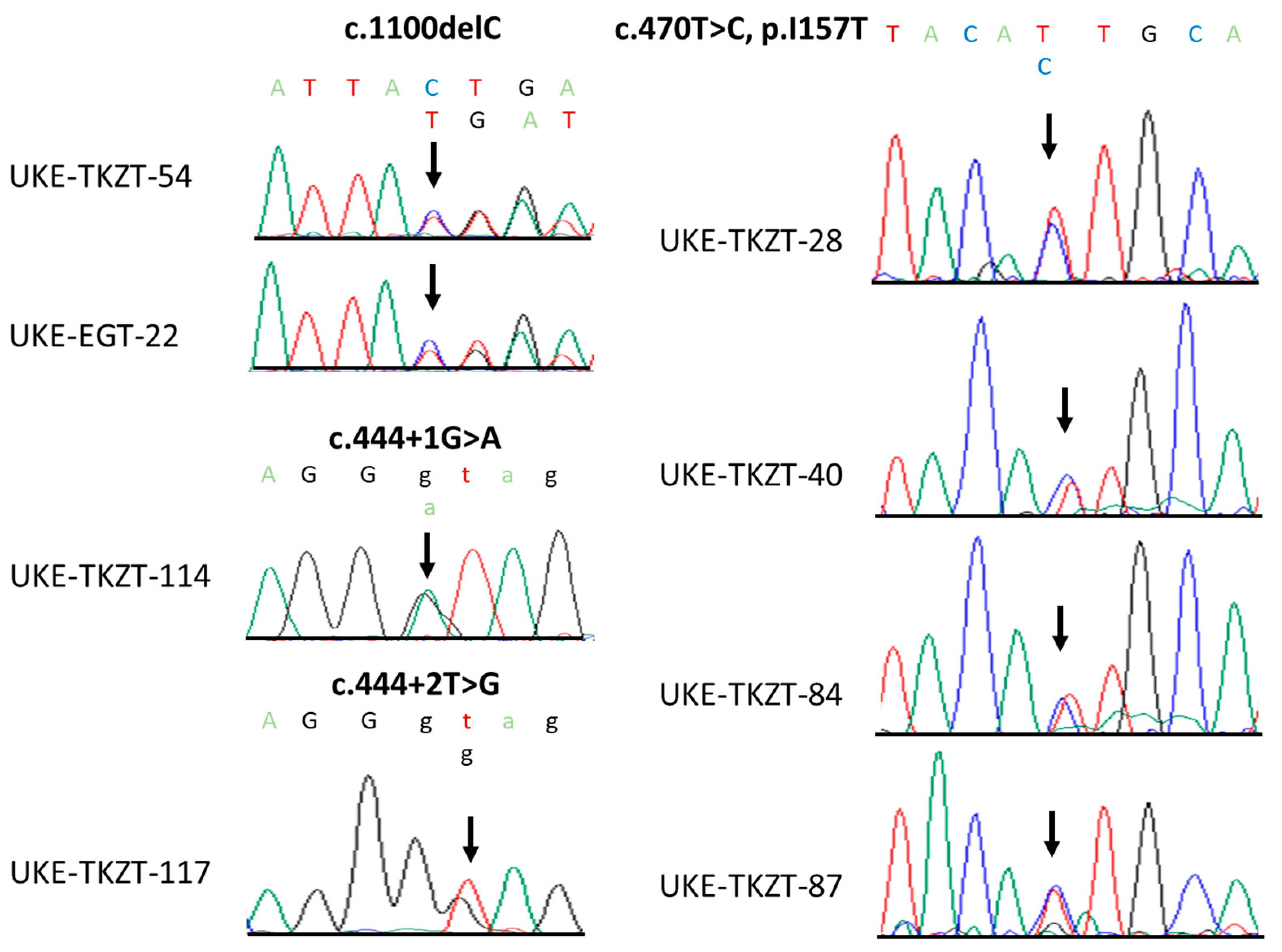
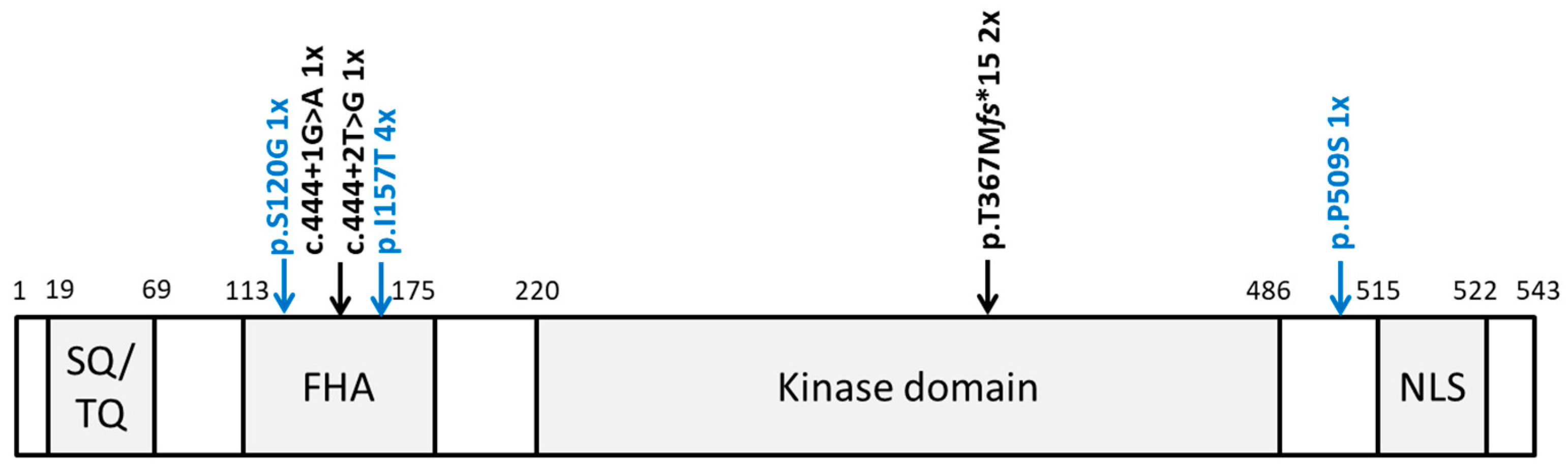
3.2. Identification of Rare CHEK2 Variants
3.3. Clinical and Histopathological Characteristics of CHEK2 Germline Variant Carriers Identified in This Study
3.4. Meta-Analysis of Histological Subtypes and Age of Diagnosis of CHEK2 Germline Variant Carriers Identified So Far
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oosterhuis, J.W.; Looijenga, L.H. Testicular germ-cell tumours in a broader perspective. Nat. Rev. Cancer 2005, 5, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Wylie, C. Germ cells. Cell 1999, 96, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Mai, P.L.; Friedlander, M.; Tucker, K.; Phillips, K.A.; Hogg, D.; Jewett, M.A.; Lohynska, R.; Daugaard, G.; Richard, S.; Bonaiti-Pellie, C.; et al. The International Testicular Cancer Linkage Consortium: A clinicopathologic descriptive analysis of 461 familial malignant testicular germ cell tumor kindred. Urol. Oncol. 2010, 28, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Litchfield, K.; Levy, M.; Huddart, R.A.; Shipley, J.; Turnbull, C. The genomic landscape of testicular germ cell tumours: From susceptibility to treatment. Nat. Rev. Urol. 2016, 13, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Shih, J.; Hollern, D.P.; Wang, L.; Bowlby, R.; Tickoo, S.K.; Thorsson, V.; Mungall, A.J.; Newton, Y.; Hegde, A.M.; et al. Integrated Molecular Characterization of Testicular Germ Cell Tumors. Cell Rep. 2018, 23, 3392–3406. [Google Scholar] [CrossRef] [PubMed]
- Czene, K.; Lichtenstein, P.; Hemminki, K. Environmental and heritable causes of cancer among 9.6 million individuals in the Swedish Family-Cancer Database. Int. J. Cancer 2002, 99, 260–266. [Google Scholar] [CrossRef]
- Litchfield, K.; Thomsen, H.; Mitchell, J.S.; Sundquist, J.; Houlston, R.S.; Hemminki, K.; Turnbull, C. Quantifying the heritability of testicular germ cell tumour using both population-based and genomic approaches. Sci. Rep. 2015, 5, 13889. [Google Scholar] [CrossRef]
- Litchfield, K.; Levy, M.; Orlando, G.; Loveday, C.; Law, P.J.; Migliorini, G.; Holroyd, A.; Broderick, P.; Karlsson, R.; Haugen, T.B.; et al. Identification of 19 new risk loci and potential regulatory mechanisms influencing susceptibility to testicular germ cell tumor. Nat. Genet. 2017, 49, 1133–1140. [Google Scholar] [CrossRef]
- Wang, Z.; McGlynn, K.A.; Rajpert-De Meyts, E.; Bishop, D.T.; Chung, C.C.; Dalgaard, M.D.; Greene, M.H.; Gupta, R.; Grotmol, T.; Haugen, T.B.; et al. Meta-analysis of five genome-wide association studies identifies multiple new loci associated with testicular germ cell tumor. Nat. Genet. 2017, 49, 1141–1147. [Google Scholar] [CrossRef]
- Pluta, J.; Pyle, L.C.; Nead, K.T.; Wilf, R.; Li, M.; Mitra, N.; Weathers, B.; D’Andrea, K.; Almstrup, K.; Anson-Cartwright, L.; et al. Identification of 22 susceptibility loci associated with testicular germ cell tumors. Nat. Commun. 2021, 12, 4487. [Google Scholar] [CrossRef]
- AlDubayan, S.H.; Pyle, L.C.; Gamulin, M.; Kulis, T.; Moore, N.D.; Taylor-Weiner, A.; Hamid, A.A.; Reardon, B.; Wubbenhorst, B.; Godse, R.; et al. Association of Inherited Pathogenic Variants in Checkpoint Kinase 2 (CHEK2) With Susceptibility to Testicular Germ Cell Tumors. JAMA Oncol. 2019, 5, 514–522. [Google Scholar] [CrossRef]
- Lessel, D.; Gamulin, M.; Kulis, T.; Toliat, M.R.; Grgic, M.; Friedrich, K.; Zunec, R.; Balija, M.; Nurnberg, P.; Kastelan, Z.; et al. Replication of genetic susceptibility loci for testicular germ cell cancer in the Croatian population. Carcinogenesis 2012, 33, 1548–1552. [Google Scholar] [CrossRef]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Kirchner, K.; Gamulin, M.; Kulis, T.; Sievers, B.; Kastelan, Z.; Lessel, D. Comprehensive Clinical and Genetic Analysis of CHEK2 in Croatian Men with Prostate Cancer. Genes 2022, 13, 1955. [Google Scholar] [CrossRef]
- Cybulski, C.; Wokolorczyk, D.; Huzarski, T.; Byrski, T.; Gronwald, J.; Gorski, B.; Debniak, T.; Masojc, B.; Jakubowska, A.; Gliniewicz, B.; et al. A large germline deletion in the Chek2 kinase gene is associated with an increased risk of prostate cancer. J. Med. Genet. 2006, 43, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Reese, M.G.; Eeckman, F.H.; Kulp, D.; Haussler, D. Improved splice site detection in Genie. J. Comput. Biol. 1997, 4, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, L.; Taniguchi, K.; Wang, X.; Cunningham, J.M.; McDonnell, S.K.; Qian, C.; Marks, A.F.; Slager, S.L.; Peterson, B.J.; et al. Mutations in CHEK2 associated with prostate cancer risk. Am. J. Hum. Genet. 2003, 72, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Havranek, O.; Kleiblova, P.; Hojny, J.; Lhota, F.; Soucek, P.; Trneny, M.; Kleibl, Z. Association of Germline CHEK2 Gene Variants with Risk and Prognosis of Non-Hodgkin Lymphoma. PLoS ONE 2015, 10, e0140819. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alfoldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Kleiblova, P.; Stolarova, L.; Krizova, K.; Lhota, F.; Hojny, J.; Zemankova, P.; Havranek, O.; Vocka, M.; Cerna, M.; Lhotova, K.; et al. Identification of deleterious germline CHEK2 mutations and their association with breast and ovarian cancer. Int. J. Cancer 2019, 145, 1782–1797. [Google Scholar] [CrossRef]
- Ni, V.I.; Ivantsov, A.O.; Kotkova, M.A.; Baskina, S.V.; Ponomareva, E.V.; Orlova, R.V.; Topuzov, E.E.; Kryukov, K.K.; Shelekhova, K.V.; Aleksakhina, S.N.; et al. Small fraction of testicular cancer cases may be causatively related to CHEK2 inactivating germ-line mutations: Evidence for somatic loss of the remaining CHEK2 allele in the tumor tissue. Fam. Cancer 2021, 20, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Bartek, J.; Lukas, J. Chk1 and Chk2 kinases in checkpoint control and cancer. Cancer Cell 2003, 3, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Cybulski, C.; Wokolorczyk, D.; Jakubowska, A.; Huzarski, T.; Byrski, T.; Gronwald, J.; Masojc, B.; Deebniak, T.; Gorski, B.; Blecharz, P.; et al. Risk of breast cancer in women with a CHEK2 mutation with and without a family history of breast cancer. J. Clin. Oncol. 2011, 29, 3747–3752. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.P.; Geng, X.P.; Ge, W.W.; Li, H. Meta-analysis of CHEK2 1100delC variant and colorectal cancer susceptibility. Eur. J. Cancer 2011, 47, 2546–2551. [Google Scholar] [CrossRef]
- Zannini, L.; Delia, D.; Buscemi, G. CHK2 kinase in the DNA damage response and beyond. J. Mol. Cell Biol. 2014, 6, 442–457. [Google Scholar] [CrossRef]
- Oishi, I.; Iwai, K.; Kagohashi, Y.; Fujimoto, H.; Kariya, K.; Kataoka, T.; Sawa, H.; Okano, H.; Otani, H.; Yamamura, H.; et al. Critical role of Caenorhabditis elegans homologs of Cds1 (Chk2)-related kinases in meiotic recombination. Mol. Cell. Biol. 2001, 21, 1329–1335. [Google Scholar] [CrossRef]
- Ertych, N.; Stolz, A.; Valerius, O.; Braus, G.H.; Bastians, H. CHK2-BRCA1 tumor-suppressor axis restrains oncogenic Aurora-A kinase to ensure proper mitotic microtubule assembly. Proc. Natl. Acad. Sci. USA 2016, 113, 1817–1822. [Google Scholar] [CrossRef]
- Potapova, T.; Gorbsky, G.J. The Consequences of Chromosome Segregation Errors in Mitosis and Meiosis. Biology 2017, 6, 12. [Google Scholar] [CrossRef]
- Oosterhuis, J.W.; Castedo, S.M.; de Jong, B.; Cornelisse, C.J.; Dam, A.; Sleijfer, D.T.; Schraffordt Koops, H. Ploidy of primary germ cell tumors of the testis. Pathogenetic and clinical relevance. Lab. Investig. 1989, 60, 14–21. [Google Scholar]
- Mayer, F.; Stoop, H.; Sen, S.; Bokemeyer, C.; Oosterhuis, J.W.; Looijenga, L.H. Aneuploidy of human testicular germ cell tumors is associated with amplification of centrosomes. Oncogene 2003, 22, 3859–3866. [Google Scholar] [CrossRef]
- Han, F.F.; Guo, C.L.; Liu, L.H. The effect of CHEK2 variant I157T on cancer susceptibility: Evidence from a meta-analysis. DNA Cell Biol. 2013, 32, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Falck, J.; Mailand, N.; Syljuasen, R.G.; Bartek, J.; Lukas, J. The ATM-Chk2-Cdc25A checkpoint pathway guards against radioresistant DNA synthesis. Nature 2001, 410, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, J.K.; Lovly, C.M.; Piwnica-Worms, H. Regulation of the Chk2 protein kinase by oligomerization-mediated cis- and trans-phosphorylation. Mol. Cancer Res. 2003, 1, 598–609. [Google Scholar]
- Crockford, G.P.; Linger, R.; Hockley, S.; Dudakia, D.; Johnson, L.; Huddart, R.; Tucker, K.; Friedlander, M.; Phillips, K.A.; Hogg, D.; et al. Genome-wide linkage screen for testicular germ cell tumour susceptibility loci. Hum. Mol. Genet. 2006, 15, 443–451. [Google Scholar] [CrossRef]
- Litchfield, K.; Levy, M.; Dudakia, D.; Proszek, P.; Shipley, C.; Basten, S.; Rapley, E.; Bishop, D.T.; Reid, A.; Huddart, R.; et al. Rare disruptive mutations in ciliary function genes contribute to testicular cancer susceptibility. Nat. Commun. 2016, 7, 13840. [Google Scholar] [CrossRef] [PubMed]
- Loveday, C.; Sud, A.; Litchfield, K.; Levy, M.; Holroyd, A.; Broderick, P.; Kote-Jarai, Z.; Dunning, A.M.; Muir, K.; Peto, J.; et al. Runs of homozygosity and testicular cancer risk. Andrology 2019, 7, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.C.; Conduit, C.; Loveland, K.L.; Thomas, B.; Lewin, J.; Tran, B. Genetics of testicular cancer: A review. Curr. Opin. Urol. 2022, 32, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Pyle, L.C.; Kim, J.; Bradfield, J.; Damrauer, S.M.; D’Andrea, K.; Einhorn, L.H.; Godse, R.; Hakonarson, H.; Kanetsky, P.A.; Kember, R.L.; et al. Germline Exome Sequencing for Men with Testicular Germ Cell Tumor Reveals Coding Defects in Chromosomal Segregation and Protein-targeting Genes. Eur. Urol. 2023. [Google Scholar] [CrossRef]
- Loveday, C.; Law, P.; Litchfield, K.; Levy, M.; Holroyd, A.; Broderick, P.; Kote-Jarai, Z.; Dunning, A.M.; Muir, K.; Peto, J.; et al. Large-scale Analysis Demonstrates Familial Testicular Cancer to have Polygenic Aetiology. Eur. Urol. 2018, 74, 248–252. [Google Scholar] [CrossRef]
- Lessel, D.; Vaz, B.; Halder, S.; Lockhart, P.J.; Marinovic-Terzic, I.; Lopez-Mosqueda, J.; Philipp, M.; Sim, J.C.; Smith, K.R.; Oehler, J.; et al. Mutations in SPRTN cause early onset hepatocellular carcinoma, genomic instability and progeroid features. Nat. Genet. 2014, 46, 1239–1244. [Google Scholar] [CrossRef]
- Oing, C.; Seidel, C.; Bokemeyer, C. Therapeutic approaches for refractory germ cell cancer. Expert Rev. Anticancer. Ther. 2018, 18, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Alsdorf, W.; Seidel, C.; Bokemeyer, C.; Oing, C. Current pharmacotherapy for testicular germ cell cancer. Expert Opin. Pharmacother. 2019, 20, 837–850. [Google Scholar] [CrossRef] [PubMed]
- de Bono, J.; Mateo, J.; Fizazi, K.; Saad, F.; Shore, N.; Sandhu, S.; Chi, K.N.; Sartor, O.; Agarwal, N.; Olmos, D.; et al. Olaparib for Metastatic Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 382, 2091–2102. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Status Total | Cases n = 129 |
|---|---|
| Mean age at diagnosis (range) | 34.99 (17–69) |
| Anatomical site | |
| Testicular (%) | 107 (83) |
| Extragonadal (%) | 22 (17) |
| Histology | |
| Seminoma (%) | 47 (36.4) |
| Non-seminoma (%) | 82 (63.6) |
| Stage grouping (AJCC) | |
| Stage I (%) | 44 (34) |
| Stage II (%) | 33 (25.7) |
| Stage III (%) | 52 (40.3) |
| Family history of GCT | |
| (%; available for cases) | 4 (3.48; 115) |
| Family history of any tumor | |
| (%; available for cases) | 58 (63; 92) |
| CHEK2 Variants | Individuals, No. (%) | OR (95% CI) Per Allele | p-Value | |
|---|---|---|---|---|
| Controls | Cases | |||
| (n = 27,173) | (n = 129) | |||
| LOFs | 210 (0.8) | 4 (3.1) | 4.06 (1.59–10.54) | 0.0191 |
| c.470T > C, p.I157T | 231 (0.9) | 4 (3.1) | 3.69 (1.45–9.55) | 0.0259 |
| CHEK2 Non LOF Carriers | CHEK2 LOF Carriers | p-Value | CHEK2 WT | CHEK2 p.I157T Carriers | p-Value | |
|---|---|---|---|---|---|---|
| Age of diagnosis (mean) a | 35.17 | 29.5 | 0.2315 | 35.14 | 36 | 0.9022 |
| Family history of GCT b | 4/111 (111/125 patients) | 0/4 | >0.9999 | 4/109 (109/121 patients) | 0/4 | >0.9999 |
| Family history of any cancer b | 55/88 (88/125 patients) | 3/4 | >0.9999 | 53/86 (86/121 patients) | 2/2 | 0.5259 |
| Histology (seminoma vs. non-seminoma) b | 45/80 | 2/2 | 0.6219 | 44/77 | 3/1 | 0.1489 |
| Aggressive GCT (>stage I) b | 83/125 | 2/4 | 0.6053 | 79/121 | 4/4 | 0.2995 |
| CHEK2 Non-LOF Carriers | CHEK2 p.I157T Carriers | |
|---|---|---|
| Age of diagnosis (mean) | 18–41 (28.19) | 14–51 (32.52) |
| Histology (seminoma vs. non-seminoma) | 14/13 | 26/26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchner, K.; Seidel, C.; Paulsen, F.-O.; Sievers, B.; Bokemeyer, C.; Lessel, D. Further Association of Germline CHEK2 Loss-of-Function Variants with Testicular Germ Cell Tumors. J. Clin. Med. 2023, 12, 7065. https://doi.org/10.3390/jcm12227065
Kirchner K, Seidel C, Paulsen F-O, Sievers B, Bokemeyer C, Lessel D. Further Association of Germline CHEK2 Loss-of-Function Variants with Testicular Germ Cell Tumors. Journal of Clinical Medicine. 2023; 12(22):7065. https://doi.org/10.3390/jcm12227065
Chicago/Turabian StyleKirchner, Kira, Christoph Seidel, Finn-Ole Paulsen, Bianca Sievers, Carsten Bokemeyer, and Davor Lessel. 2023. "Further Association of Germline CHEK2 Loss-of-Function Variants with Testicular Germ Cell Tumors" Journal of Clinical Medicine 12, no. 22: 7065. https://doi.org/10.3390/jcm12227065
APA StyleKirchner, K., Seidel, C., Paulsen, F.-O., Sievers, B., Bokemeyer, C., & Lessel, D. (2023). Further Association of Germline CHEK2 Loss-of-Function Variants with Testicular Germ Cell Tumors. Journal of Clinical Medicine, 12(22), 7065. https://doi.org/10.3390/jcm12227065