
Noninvasive Neuromodulation in Parkinson’s Disease: Insights from Animal Models
 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Transcranial Magnetic Stimulation (TMS)
3.1.1. Background
3.1.2. Animal Studies

{kind=link}
{kind=link}
| Study | PD Model | Sham Control | TMS Parameters | Outcome Measures and Findings |
|---|---|---|---|---|
| Ba, et al. (2017) [33] | Lactacystin-induced (rat) |
|
|
|
| Yang, et al. (2010) [24] | 6-OHDA (rat) |
|
|
|
| Ghiglieri, et al. (2012) [28] | 6-OHDA (rat) |
|
|
|
| Lee, et al. (2013) [25] | 6-OHDA (rat) |
|
|
|
| Dong, et al. (2015) [34] | MPTP (mouse) |
|
|
|
| Hsieh, et al., 2015 [26] | 6-OHDA (rat) |
|
|
|
| Cacace, et al. (2017) [30] | 6-OHDA (rat) |
|
|
|
| Ba, et al. (2019) [36] | 6-OHDA (mouse) |
|
|
|
| Hsieh, et al. (2021) [27] | 6-OHDA (rat) |
|
|
|
| Ba, et al. (2016) [32] | 6-OHDA + levodopa (rat) |
|
|
|
| Sun, et al. (2023) [35] | MPTP (mouse) |
|
|
|
| Kang, et al. (2022) [31] | 6-OHDA or LPS (rat) |
|
|
|
| Jovanovic, et al. (2023) [29] | 6-OHDA (rat) |
|
|
|
3.1.3. Limitations
3.2. Transcranial Direct Current Stimulation (tDCS) and Transcranial Alternating Current Stimulation (tACS)
3.2.1. Background
3.2.2. Animal Models
| Study | PD Model | Sham Control | tDCS Parameters | Outcome Measures and Findings |
|---|---|---|---|---|
| Li, et al. (2011) [79] | 6-OHDA (rat) |
|
|
|
| Li, et al. (2015) [77] | MPTP (monkey) |
|
|
|
| Lu, et al. (2015) [69] | MPTP (mouse) |
|
|
|
| Winkler, et al. (2017) [76] * | 6-OHDA (rat) |
|
|
|
| Lee, et al. (2018) [70] ** | MPTP (mouse) |
|
|
|
| Kade, et al. (2019) [73] | Rotenone (rat) |
|
|
|
| Feng, et al. (2020) [71] | 6-OHDA (rat) |
|
|
|
| Lee, et al. (2022) [72] | MPTP (mouse) |
|
|
|
3.2.3. Limitations
3.3. Electroconvulsive Therapy (ECT)
3.3.1. Background
3.3.2. Animal Models
| Study | PD Model | Sham Control | ECT Parameters | Outcome Measures |
|---|---|---|---|---|
| Garcia & Sotelo (1993) [82] | MPTP (mouse) |
|
|
|
| Anastasia, et al. (2007) [83] | 6-OHDA (rat) |
|
|
|
| Strome, et al. (2007) [84] | 6-OHDA (rat) |
|
|
|
| Anastasia, et al. (2011) [85] | 6-OHDA (rat) |
|
|
|
| Landau, et al. (2012) [86] | MPTP (monkey) |
|
|
|
| Yang, et al. (2020) [87] * | MPTP (mouse) |
|
|
|
| Huh, et al. (2023) [88] | MPTP (mouse) |
|
|
|
3.3.3. Limitations
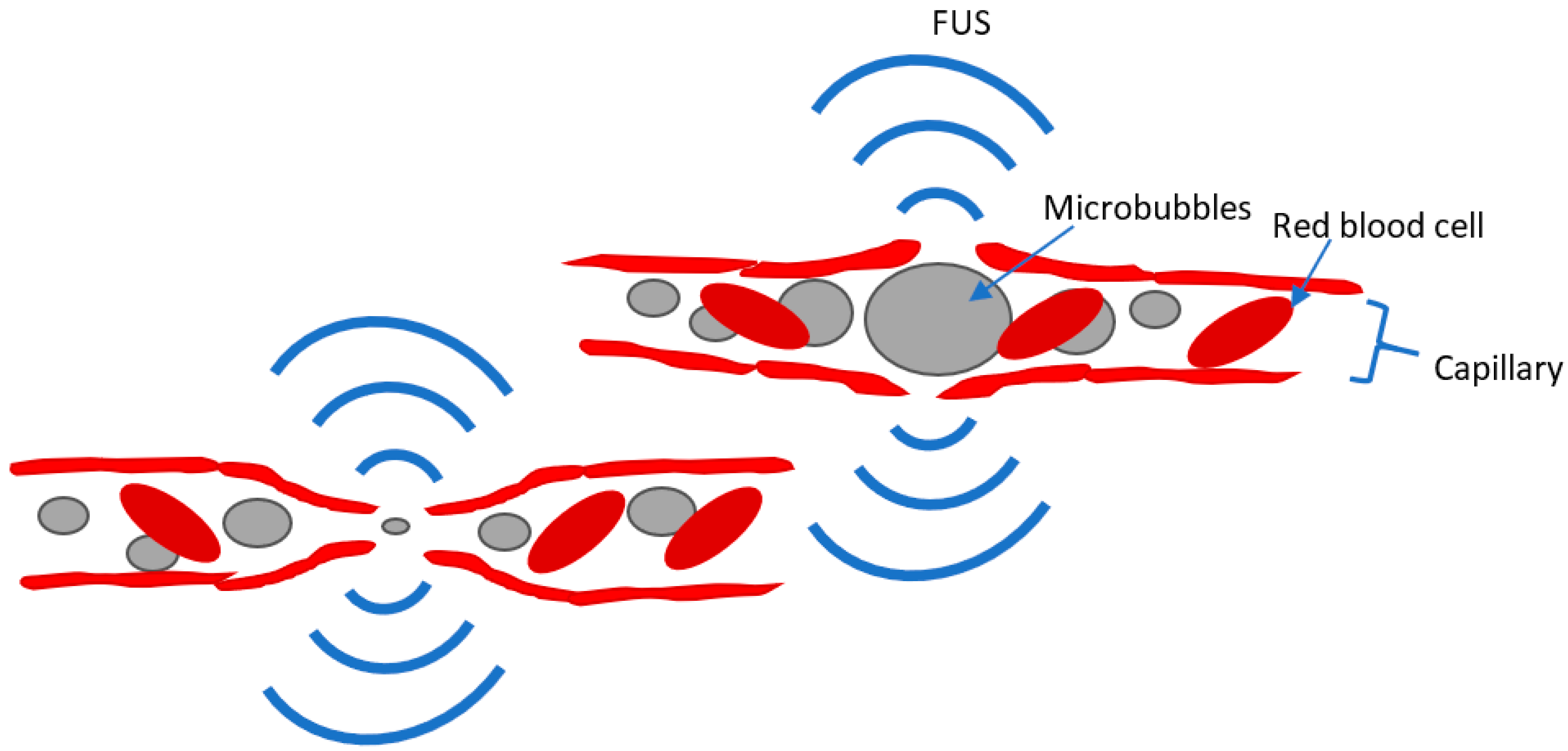
3.4. Focused Ultrasound (FUS)
3.4.1. Background
3.4.2. LIFU Animal Models
| Study | PD Model | Sham Control | Ultrasound Parameters | Outcome Measures |
|---|---|---|---|---|
| Wang, et al. (2019) [104] | MPTP (mouse) |
|
|
|
| Zhou, et al. (2019) (2) [106] | MPTP (mouse) |
|
|
|
| Zhou, et al. (2019) (1) [105] | MPTP (mouse) |
|
|
|
| Xu, et al. (2020) [107] | MPTP (mouse) and cell line (not discussed) |
|
|
|
| Dong, et al. (2021) [110] | 6-OHDA (rat) |
|
|
|
| Zhou, et al. (2021) [108] | MPTP (mouse) |
|
|
|
| Song, et al. (2022) [109] | 6-OHDA (rat) |
|
|
|
3.4.3. FUS-MB Animal Models
| Study | PD Model | Sham Control | FUS-MB Parameters | Outcome Measures |
|---|---|---|---|---|
| Fan, et al. (2016) [115] | 6-OHDA (rat) |
|
|
|
| Lin, et al. (2016) [116] | MPTP (mouse) |
|
|
|
| Long, et al. (2017) [121] | 6-OHDA (rat) |
|
|
|
| Mead, et al. (2017) [117] | 6-OHDA (rat) |
|
|
|
| Xhima, et al. (2018) [137] | Transgenic humanized alpha-synuclein (mouse) |
|
|
|
| Yue, et al. (2018) [118] | 6-OHDA (rat) |
|
|
|
| Zhang, et al. (2018) [129] ** | MPTP (mouse) |
|
|
|
| Niu, et al. (2018) [125] ** | 6-OHDA (rat) |
|
|
|
| Ji, et al. (2019) [123] | MPTP (mouse) |
|
|
|
| Karakatsani, et al. (2019) [119] | MPTP (mouse, early PD model) |
|
|
|
| Wu, et al. (2020) [122] ** | MitoPark (mouse, genetic) |
|
|
|
| Yan, et al. (2021) [128] | MPTP (mouse) |
|
|
|
| Feng, et al. (2022) [132] ** | rAAV2/5-wild type α-syn or A53T α-syn (mouse) |
|
|
|
| Wang, et al. (2022) [135] | MPTP (mouse) |
|
|
|
| Kim, et al. (2023) [139] | MPTP (mouse) |
|
|
|
3.4.4. Limitations
4. Discussion and Future Directions
4.1. Applications to Clinical Practice
4.2. Limitations
4.3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Goldman, J.G.; Postuma, R. Premotor and Nonmotor Features of Parkinson’s Disease. Curr. Opin. Neurol. 2014, 27, 434. [Google Scholar] [CrossRef]
- Roheger, M.; Kalbe, E.; Liepelt-Scarfone, I. Progression of Cognitive Decline in Parkinson’s Disease. J. Park. Dis. 2018, 8, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Van Den Eeden, S.K.; Tanner, C.M.; Bernstein, A.L.; Fross, R.D.; Leimpeter, A.; Bloch, D.A.; Nelson, L.M. Incidence of Parkinson’s Disease: Variation by Age, Gender, and Race/Ethnicity. Am. J. Epidemiol. 2003, 157, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Moosa, S.; Martínez-Fernández, R.; Elias, W.J.; del Alamo, M.; Eisenberg, H.M.; Fishman, P.S. The Role of High-Intensity Focused Ultrasound as a Symptomatic Treatment for Parkinson’s Disease. Mov. Disord. 2019, 34, 1243–1251. [Google Scholar] [CrossRef] [PubMed]
- Begemann, M.J.; Brand, B.A.; Ćurčić-Blake, B.; Aleman, A.; Sommer, I.E. Efficacy of Non-Invasive Brain Stimulation on Cognitive Functioning in Brain Disorders: A Meta-Analysis. Psychol. Med. 2020, 50, 2465–2486. [Google Scholar] [CrossRef] [PubMed]
- Brak, I.V.; Filimonova, E.; Zakhariya, O.; Khasanov, R.; Stepanyan, I. Transcranial Current Stimulation as a Tool of Neuromodulation of Cognitive Functions in Parkinson’s Disease. Front. Neurosci. 2022, 16, 781488. [Google Scholar] [CrossRef]
- Ridding, M.C.; Rothwell, J.C. Is There a Future for Therapeutic Use of Transcranial Magnetic Stimulation? Nat. Rev. Neurosci. 2007, 8, 559–567. [Google Scholar] [CrossRef]
- Rizvi, S.; Khan, A.M. Use of Transcranial Magnetic Stimulation for Depression. Cureus 2019, 11, e4736. [Google Scholar] [CrossRef]
- Cheng, B.; Zhu, T.; Zhao, W.; Sun, L.; Shen, Y.; Xiao, W.; Zhang, S. Effect of Theta Burst Stimulation-Patterned RTMS on Motor and Nonmotor Dysfunction of Parkinson’s Disease: A Systematic Review and Metaanalysis. Front. Neurol. 2022, 12, 762100. [Google Scholar] [CrossRef]
- Somaa, F.A.; de Graaf, T.A.; Sack, A.T. Transcranial Magnetic Stimulation in the Treatment of Neurological Diseases. Front. Neurol. 2022, 13, 793253. [Google Scholar] [CrossRef]
- Benninger, D.H.; Hallett, M. Non-Invasive Brain Stimulation for Parkinson’s Disease: Current Concepts and Outlook 2015. NeuroRehabilitation 2015, 37, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; Simon, D.K.; Wu, A.; Pascual-Leone, A. Non-Invasive Brain Stimulation for Parkinson’s Disease: A Systematic Review and Meta-Analysis of the Literature. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1614–1623. [Google Scholar] [CrossRef] [PubMed]
- Elahi, B.; Elahi, B.; Chen, R. Effect of Transcranial Magnetic Stimulation on Parkinson Motor Function–Systematic Review of Controlled Clinical Trials. Mov. Disord. 2009, 24, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Dong, Z.; Pan, L.; Liu, Y.; Ye, Z.; Qin, L.; Liu, Q.; Qin, C. Effects of Repetitive Transcranial Magnetic Stimulation on Gait Disorders and Cognitive Dysfunction in Parkinson’s Disease: A Systematic Review with Meta-analysis. Brain Behav. 2022, 12, e2697. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.; Hickey, P.T.; Sundman, M.; Song, A.W.; Chen, N. Effects of Repetitive Transcranial Magnetic Stimulation on Motor Symptoms in Parkinson Disease: A Systematic Review and Meta-Analysis. JAMA Neurol. 2015, 72, 432–440. [Google Scholar] [CrossRef]
- Zanjani, A.; Zakzanis, K.K.; Daskalakis, Z.J.; Chen, R. Repetitive Transcranial Magnetic Stimulation of the Primary Motor Cortex in the Treatment of Motor Signs in Parkinson’s Disease: A Quantitative Review of the Literature. Mov. Disord. 2015, 30, 750–758. [Google Scholar] [CrossRef]
- Li, S.; Jiao, R.; Zhou, X.; Chen, S. Motor Recovery and Antidepressant Effects of Repetitive Transcranial Magnetic Stimulation on Parkinson Disease: A PRISMA-Compliant Meta-Analysis. Medicine 2020, 99, e19642. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, Z.; McClure, M.A.; He, L.; Mu, Q. Effect of RTMS on Parkinson’s Cognitive Function: A Systematic Review and Meta-Analysis. BMC Neurol. 2020, 20, 377. [Google Scholar] [CrossRef]
- Zhu, H.; Lu, Z.; Jin, Y.; Duan, X.; Teng, J.; Duan, D. Low-Frequency Repetitive Transcranial Magnetic Stimulation on Parkinson Motor Function: A Meta-Analysis of Randomised Controlled Trials. Acta Neuropsychiatr. 2015, 27, 82–89. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, X.; Zeng, W.; Zhai, H.; Zhang, X.; Yang, X.; Cheng, C.; Wang, J.; Yang, X.; Xu, Y. Transcranial Magnetic Stimulation Alleviates Levodopa-Induced Dyskinesia in Parkinson’s Disease and the Related Mechanisms: A Mini-Review. Front. Neurol. 2021, 12, 758345. [Google Scholar] [CrossRef]
- Keck, M.E.; Welt, T.; Müller, M.B.; Erhardt, A.; Ohl, F.; Toschi, N.; Holsboer, F.; Sillaber, I. Repetitive Transcranial Magnetic Stimulation Increases the Release of Dopamine in the Mesolimbic and Mesostriatal System. Neuropharmacology 2002, 43, 101–109. [Google Scholar] [CrossRef]
- Kanno, M.; Matsumoto, M.; Togashi, H.; Yoshioka, M.; Mano, Y. Effects of Acute Repetitive Transcranial Magnetic Stimulation on Dopamine Release in the Rat Dorsolateral Striatum. J. Neurol. Sci. 2004, 217, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Strafella, A.P.; Paus, T.; Barrett, J.; Dagher, A. Repetitive Transcranial Magnetic Stimulation of the Human Prefrontal Cortex Induces Dopamine Release in the Caudate Nucleus. J. Neurosci. 2001, 21, RC157. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Song, L.; Liu, Z. The Effect of Repetitive Transcranial Magnetic Stimulation on a Model Rat of Parkinson’s Disease. Neuroreport 2010, 21, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, S.H.; Ko, A.-R.; Lee, J.S.; Yu, J.H.; Seo, J.H.; Cho, B.P.; Cho, S.-R. Therapeutic Effects of Repetitive Transcranial Magnetic Stimulation in an Animal Model of Parkinson’s Disease. Brain Res. 2013, 1537, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.-H.; Huang, Y.-Z.; Rotenberg, A.; Pascual-Leone, A.; Chiang, Y.-H.; Wang, J.-Y.; Chen, J.-J.J. Functional Dopaminergic Neurons in Substantia Nigra Are Required for Transcranial Magnetic Stimulation-Induced Motor Plasticity. Cereb. Cortex 2015, 25, 1806–1814. [Google Scholar] [CrossRef]
- Hsieh, T.-H.; He, X.-K.; Liu, H.-H.; Chen, J.-J.J.; Peng, C.-W.; Liu, H.-L.; Rotenberg, A.; Chen, K.-T.; Chang, M.-Y.; Chiang, Y.-H.; et al. Early Repetitive Transcranial Magnetic Stimulation Exerts Neuroprotective Effects and Improves Motor Functions in Hemiparkinsonian Rats. Neural Plast. 2021, 2021, 1763533. [Google Scholar] [CrossRef]
- Ghiglieri, V.; Pendolino, V.; Sgobio, C.; Bagetta, V.; Picconi, B.; Calabresi, P. Θ-Burst Stimulation and Striatal Plasticity in Experimental Parkinsonism. Exp. Neurol. 2012, 236, 395–398. [Google Scholar] [CrossRef]
- Zeljkovic Jovanovic, M.; Stanojevic, J.; Stevanovic, I.; Stekic, A.; Bolland, S.J.; Jasnic, N.; Ninkovic, M.; Zaric Kontic, M.; Ilic, T.V.; Rodger, J.; et al. Intermittent Theta Burst Stimulation Improves Motor and Behavioral Dysfunction through Modulation of NMDA Receptor Subunit Composition in Experimental Model of Parkinson’s Disease. Cells 2023, 12, 1525. [Google Scholar] [CrossRef]
- Cacace, F.; Mineo, D.; Viscomi, M.T.; Latagliata, E.C.; Mancini, M.; Sasso, V.; Vannelli, A.; Pascucci, T.; Pendolino, V.; Marcello, E.; et al. Intermittent Theta-Burst Stimulation Rescues Dopamine-Dependent Corticostriatal Synaptic Plasticity and Motor Behavior in Experimental Parkinsonism: Possible Role of Glial Activity. Mov. Disord. 2017, 32, 1035–1046. [Google Scholar] [CrossRef]
- Kang, X.; Zhang, B.; Du, W.; Zhao, R.; Liu, X.; Bai, Y.; Jiang, X.; Pang, J.; Zhao, C.; Mou, X.; et al. High-Frequency Repetitive Transcranial Magnetic Stimulation Regulates Astrocyte Activation by Modulating the Endocannabinoid System in Parkinson’s Disease. Mol. Neurobiol. 2022, 59, 5121–5134. [Google Scholar] [CrossRef]
- Ba, M.; Kong, M.; Guan, L.; Yi, M.; Zhang, H. Repetitive Transcranial Magnetic Stimulation (RTMS) Improves Behavioral and Biochemical Deficits in Levodopa-Induced Dyskinetic Rats Model. Oncotarget 2016, 7, 58802–58812. [Google Scholar] [CrossRef] [PubMed]
- Ba, M.; Ma, G.; Ren, C.; Sun, X.; Kong, M. Repetitive Transcranial Magnetic Stimulation for Treatment of Lactacystin-Induced Parkinsonian Rat Model. Oncotarget 2017, 8, 50921–50929. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Wang, Y.; Gu, P.; Shao, R.; Zhao, L.; Liu, X.; Wang, Z.; Wang, M. The Neuroprotective Mechanism of Low-Frequency RTMS on Nigral Dopaminergic Neurons of Parkinson’s Disease Model Mice. Park. Dis. 2015, 2015, 564095. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, F.; Han, J.; Bai, L.; Du, J. Repetitive Transcranial Magnetic Stimulation Reduces Neuronal Loss and Neuroinflammation in Parkinson?S Disease Through the MiR-195a-5p/CREB Axis. Turk. Neurosurg. 2023, 33, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Ba, F.; Zhou, Y.; Zhou, J.; Chen, X. Repetitive Transcranial Magnetic Stimulation Protects Mice against 6-OHDA-Induced Parkinson’s Disease Symptoms by Regulating Brain Amyloid Β1–42 Level. Mol. Cell. Biochem. 2019, 458, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Björklund, A.; Dunnett, S.B. The Amphetamine Induced Rotation Test: A Re-Assessment of Its Use as a Tool to Monitor Motor Impairment and Functional Recovery in Rodent Models of Parkinson’s Disease. J. Park. Dis. 2019, 9, 17–29. [Google Scholar] [CrossRef]
- van Dijk, K.D.; Teunissen, C.E.; Drukarch, B.; Jimenez, C.R.; Groenewegen, H.J.; Berendse, H.W.; van de Berg, W.D.J. Diagnostic Cerebrospinal Fluid Biomarkers for Parkinson’s Disease: A Pathogenetically Based Approach. Neurobiol. Dis. 2010, 39, 229–241. [Google Scholar] [CrossRef]
- More, S.V.; Choi, D.-K. Promising Cannabinoid-Based Therapies for Parkinson’s Disease: Motor Symptoms to Neuroprotection. Mol. Neurodegener. 2015, 10, 17. [Google Scholar] [CrossRef]
- Ba, M.; Kong, M.; Ma, G. Postsynaptic Density Protein 95-Regulated NR2B Tyrosine Phosphorylation and Interactions of Fyn with NR2B in Levodopa-Induced Dyskinesia Rat Models. Drug Des. Dev. Ther. 2015, 9, 199–206. [Google Scholar] [CrossRef]
- Oh, J.D.; Russell, D.S.; Vaughan, C.L.; Chase, T.N. Enhanced Tyrosine Phosphorylation of Striatal NMDA Receptor Subunits: Effect of Dopaminergic Denervation and L-DOPA Administration. Brain Res. 1998, 813, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Suppa, A.; Marsili, L.; Belvisi, D.; Conte, A.; Iezzi, E.; Modugno, N.; Fabbrini, G.; Berardelli, A. Lack of LTP-like Plasticity in Primary Motor Cortex in Parkinson’s Disease. Exp. Neurol. 2011, 227, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, Y.; Zhou, J. Neuroinflammation in Parkinson’s Disease and Its Potential as Therapeutic Target. Transl. Neurodegener. 2015, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Troncoso-Escudero, P.; Parra, A.; Nassif, M.; Vidal, R.L. Outside in: Unraveling the Role of Neuroinflammation in the Progression of Parkinson’s Disease. Front. Neurol. 2018, 9, 860. [Google Scholar] [CrossRef] [PubMed]
- Tansey, M.G.; Wallings, R.L.; Houser, M.C.; Herrick, M.K.; Keating, C.E.; Joers, V. Inflammation and Immune Dysfunction in Parkinson Disease. Nat. Rev. Immunol. 2022, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]
- Aftanas, L.I.; Gevorgyan, M.M.; Zhanaeva, S.Y.; Dzemidovich, S.S.; Kulikova, K.I.; Al’perina, E.L.; Danilenko, K.V.; Idova, G.V. Therapeutic Effects of Repetitive Transcranial Magnetic Stimulation (RTMS) on Neuroinflammation and Neuroplasticity in Patients with Parkinson’s Disease: A Placebo-Controlled Study. Bull. Exp. Biol. Med. 2018, 165, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Lee, J.; Lee, S.-Y. Recent Progress in the Development of TSPO PET Ligands for Neuroinflammation Imaging in Neurological Diseases. Nucl. Med. Mol. Imaging 2017, 51, 283–296. [Google Scholar] [CrossRef]
- Zhang, P.-F.; Gao, F. Neuroinflammation in Parkinson’s Disease: A Meta-Analysis of PET Imaging Studies. J. Neurol. 2022, 269, 2304–2314. [Google Scholar] [CrossRef]
- Doot, R.K.; Young, A.J.; Nasrallah, I.M.; Wetherill, R.R.; Siderowf, A.; Mach, R.H.; Dubroff, J.G. [18F]NOS PET Brain Imaging Suggests Elevated Neuroinflammation in Idiopathic Parkinson’s Disease. Cells 2022, 11, 3081. [Google Scholar] [CrossRef]
- Palasz, E.; Wysocka, A.; Gasiorowska, A.; Chalimoniuk, M.; Niewiadomski, W.; Niewiadomska, G. BDNF as a Promising Therapeutic Agent in Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 1170. [Google Scholar] [CrossRef]
- Abercrombie, E.D.; Keefe, K.A.; DiFrischia, D.S.; Zigmond, M.J. Differential Effect of Stress on In Vivo Dopamine Release in Striatum, Nucleus Accumbens, and Medial Frontal Cortex. J. Neurochem. 1989, 52, 1655–1658. [Google Scholar] [CrossRef]
- Wexler, A. The Practices of Do-It-Yourself Brain Stimulation: Implications for Ethical Considerations and Regulatory Proposals. J. Med. Ethics 2016, 42, 211–215. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Boggio, P.S.; Fregni, F.; Pascual-Leone, A. Treatment of Depression with Transcranial Direct Current Stimulation (TDCS): A Review. Exp. Neurol. 2009, 219, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Marangolo, P.; Fiori, V.; Calpagnano, M.A.; Campana, S.; Razzano, C.; Caltagirone, C.; Marini, A. TDCS over the Left Inferior Frontal Cortex Improves Speech Production in Aphasia. Front. Hum. Neurosci. 2013, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Holland, R.; Crinion, J. Can TDCS Enhance Treatment of Aphasia after Stroke? Aphasiology 2012, 26, 1169–1191. [Google Scholar] [CrossRef] [PubMed]
- Boggio, P.S.; Ferrucci, R.; Rigonatti, S.P.; Covre, P.; Nitsche, M.; Pascual-Leone, A.; Fregni, F. Effects of Transcranial Direct Current Stimulation on Working Memory in Patients with Parkinson’s Disease. J. Neurol. Sci. 2006, 249, 31–38. [Google Scholar] [CrossRef]
- Ferrucci, R.; Mameli, F.; Ruggiero, F.; Priori, A. Transcranial Direct Current Stimulation as Treatment for Parkinson’s Disease and Other Movement Disorders. Basal Ganglia 2016, 6, 53–61. [Google Scholar] [CrossRef]
- Lefaucheur, J.-P.; Antal, A.; Ayache, S.S.; Benninger, D.H.; Brunelin, J.; Cogiamanian, F.; Cotelli, M.; De Ridder, D.; Ferrucci, R.; Langguth, B.; et al. Evidence-Based Guidelines on the Therapeutic Use of Transcranial Direct Current Stimulation (TDCS). Clin. Neurophysiol. 2017, 128, 56–92. [Google Scholar] [CrossRef]
- Nitsche, M.A.; Paulus, W. Excitability Changes Induced in the Human Motor Cortex by Weak Transcranial Direct Current Stimulation. J. Physiol. 2000, 527, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Kronberg, G.; Bridi, M.; Abel, T.; Bikson, M.; Parra, L.C. Direct Current Stimulation Modulates LTP and LTD: Activity Dependence and Dendritic Effects. Brain Stimul. 2017, 10, 51–58. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Liu, Z.; Rao, J.; Wang, J.; Wang, P.; Gong, X.; Wen, Y. Transcranial Direct Current Stimulation for Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2021, 13, 746797. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; Boggio, P.S.; Santos, M.C.; Lima, M.; Vieira, A.L.; Rigonatti, S.P.; Silva, M.T.A.; Barbosa, E.R.; Nitsche, M.A.; Pascual-Leone, A. Noninvasive Cortical Stimulation with Transcranial Direct Current Stimulation in Parkinson’s Disease. Mov. Disord. 2006, 21, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Guerra, A.; Colella, D.; Giangrosso, M.; Cannavacciuolo, A.; Paparella, G.; Fabbrini, G.; Suppa, A.; Berardelli, A.; Bologna, M. Driving Motor Cortex Oscillations Modulates Bradykinesia in Parkinson’s Disease. Brain J. Neurol. 2022, 145, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Del Felice, A.; Castiglia, L.; Formaggio, E.; Cattelan, M.; Scarpa, B.; Manganotti, P.; Tenconi, E.; Masiero, S. Personalized Transcranial Alternating Current Stimulation (TACS) and Physical Therapy to Treat Motor and Cognitive Symptoms in Parkinson’s Disease: A Randomized Cross-over Trial. NeuroImage Clin. 2019, 22, 101768. [Google Scholar] [CrossRef]
- Brittain, J.-S.; Probert-Smith, P.; Aziz, T.Z.; Brown, P. Tremor Suppression by Rhythmic Transcranial Current Stimulation. Curr. Biol. 2013, 23, 436–440. [Google Scholar] [CrossRef]
- Tanaka, T.; Takano, Y.; Tanaka, S.; Hironaka, N.; Kobayashi, K.; Hanakawa, T.; Watanabe, K.; Honda, M. Transcranial Direct-Current Stimulation Increases Extracellular Dopamine Levels in the Rat Striatum. Front. Syst. Neurosci. 2013, 7, 6. [Google Scholar] [CrossRef]
- Takano, Y.; Yokawa, T.; Masuda, A.; Niimi, J.; Tanaka, S.; Hironaka, N. A Rat Model for Measuring the Effectiveness of Transcranial Direct Current Stimulation Using FMRI. Neurosci. Lett. 2011, 491, 40–43. [Google Scholar] [CrossRef]
- Liebetanz, D.; Koch, R.; Mayenfels, S.; König, F.; Paulus, W.; Nitsche, M.A. Safety Limits of Cathodal Transcranial Direct Current Stimulation in Rats. Clin. Neurophysiol. 2009, 120, 1161–1167. [Google Scholar] [CrossRef]
- Lu, C.; Wei, Y.; Hu, R.; Wang, Y.; Li, K.; Li, X. Transcranial Direct Current Stimulation Ameliorates Behavioral Deficits and Reduces Oxidative Stress in 1-Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine-Induced Mouse Model of Parkinson’s Disease. Neuromodul. Technol. Neural Interface 2015, 18, 442–447. [Google Scholar] [CrossRef]
- Lee, S.-B.; Kim, H.-T.; Yang, H.O.; Jang, W. Anodal Transcranial Direct Current Stimulation Prevents Methyl-4-Phenyl-1,2,3,6-Tetrahydropyridine (MPTP)-Induced Neurotoxicity by Modulating Autophagy in an in Vivo Mouse Model of Parkinson’s Disease. Sci. Rep. 2018, 8, 15165. [Google Scholar] [CrossRef]
- Feng, X.-J.; Huang, Y.-T.; Huang, Y.-Z.; Kuo, C.-W.; Peng, C.-W.; Rotenberg, A.; Juan, C.-H.; Pei, Y.-C.; Chen, Y.-H.; Chen, K.-Y.; et al. Early Transcranial Direct Current Stimulation Treatment Exerts Neuroprotective Effects on 6-OHDA-Induced Parkinsonism in Rats. Brain Stimul. 2020, 13, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Jung, D.H.; Jung, Y.J.; Shin, H.K.; Choi, B.T. Transcranial Alternating Current Stimulation Rescues Motor Deficits in a Mouse Model of Parkinson’s Disease via the Production of Glial Cell Line-Derived Neurotrophic Factor. Brain Stimul. 2022, 15, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Kade, A.K.; Kravchenko, S.V.; Trofimenko, A.I.; Chaplygina, K.Y.; Ananeva, E.I.; Poliakov, P.P.; Lipatova, A.S. [The efficacy of tes-therapy for treatment of anxiety-like behavior and motor disorders in rats with an experimental model of parkinsonism]. Zhurnal Nevrologii i Psikhiatrii imeni S. S. Korsakova 2019, 119, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Dias, V.; Junn, E.; Mouradian, M.M. The Role of Oxidative Stress in Parkinson’s Disease. J. Park. Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Lynch-Day, M.A.; Mao, K.; Wang, K.; Zhao, M.; Klionsky, D.J. The Role of Autophagy in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009357. [Google Scholar] [CrossRef]
- Winkler, C.; Reis, J.; Hoffmann, N.; Gellner, A.-K.; Münkel, C.; Curado, M.R.; Furlanetti, L.; Garcia, J.; Döbrössy, M.D.; Fritsch, B. Anodal Transcranial Direct Current Stimulation Enhances Survival and Integration of Dopaminergic Cell Transplants in a Rat Parkinson Model. eNeuro 2017, 4, ENEURO.0063-17.2017. [Google Scholar] [CrossRef]
- Li, H.; Lei, X.; Yan, T.; Li, H.; Huang, B.; Li, L.; Xu, L.; Liu, L.; Chen, N.; Lü, L.; et al. The Temporary and Accumulated Effects of Transcranial Direct Current Stimulation for the Treatment of Advanced Parkinson’s Disease Monkeys. Sci. Rep. 2015, 5, 12178. [Google Scholar] [CrossRef]
- Mounayar, S.; Boulet, S.; Tandé, D.; Jan, C.; Pessiglione, M.; Hirsch, E.C.; Féger, J.; Savasta, M.; François, C.; Tremblay, L. A New Model to Study Compensatory Mechanisms in MPTP-Treated Monkeys Exhibiting Recovery. Brain J. Neurol. 2007, 130, 2898–2914. [Google Scholar] [CrossRef]
- Li, Y.; Tian, X.; Qian, L.; Yu, X.; Jiang, W. Anodal Transcranial Direct Current Stimulation Relieves the Unilateral Bias of a Rat Model of Parkinson’s Disease. In Proceedings of the 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 30 August–3 September 2011; pp. 765–768. [Google Scholar]
- Pagnin, D.; de Queiroz, V.; Pini, S.; Cassano, G.B. Efficacy of ECT in Depression: A Meta-Analytic Review. J. ECT 2004, 20, 13–20. [Google Scholar] [CrossRef]
- Kennedy, R.; Mittal, D.; O’Jile, J. Electroconvulsive Therapy in Movement Disorders: An Update. J. Neuropsychiatry Clin. Neurosci. 2003, 15, 407–421. [Google Scholar] [CrossRef]
- Garcia, E.; Sotelo, J. Electroconvulsive Shock Does Not Modify Striatal Contents of Dopamine in MPTP-Treated Mice. Neurochem. Res. 1993, 18, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Anastasia, A.; De Erausquin, G.A.; Wojnacki, J.; Mascó, D.H. Protection of Dopaminergic Neurons by Electroconvulsive Shock in an Animal Model of Parkinson’s Disease. J. Neurochem. 2007, 103, 1542–1552. [Google Scholar] [CrossRef] [PubMed]
- Strome, E.M.; Zis, A.P.; Doudet, D.J. Electroconvulsive Shock Enhances Striatal Dopamine D1 and D3 Receptor Binding and Improves Motor Performance in 6-OHDA-Lesioned Rats. J. Psychiatry Neurosci. 2007, 32, 193–202. [Google Scholar] [PubMed]
- Anastasía, A.; Wojnacki, J.; de Erausquin, G.A.; Mascó, D.H. Glial Cell-Line Derived Neurotrophic Factor Is Essential for Electroconvulsive Shock-Induced Neuroprotection in an Animal Model of Parkinson’s Disease. Neuroscience 2011, 195, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Landau, A.M.; Clark, C.; Jivan, S.; Doudet, D.J. Antiparkinsonian Mechanism of Electroconvulsive Therapy in MPTP-Lesioned Non-Human Primates. Neurodegener. Dis. 2012, 9, 128–138. [Google Scholar] [CrossRef]
- Yang, C.; Qiu, Y.; Qing, Y.; Xu, J.; Dai, W.; Hu, X.; Wu, X. Synergistic Effect of Electric Stimulation and Mesenchymal Stem Cells against Parkinson’s Disease. Aging 2020, 12, 16062–16071. [Google Scholar] [CrossRef]
- Huh, S.; Yu, H.S.; Kang, N.; Ahn, Y.M.; Kim, Y.S.; Kim, S.H. Electroconvulsive Seizure Normalizes Motor Deficits and Induces Autophagy Signaling in the MPTP-Induced Parkinson Disease Mouse Model. Psychiatry Investig. 2023, 20, 273–283. [Google Scholar] [CrossRef]
- Robertson, D.C.; Schmidt, O.; Ninkina, N.; Jones, P.A.; Sharkey, J.; Buchman, V.L. Developmental Loss and Resistance to MPTP Toxicity of Dopaminergic Neurones in Substantia Nigra Pars Compacta of Gamma-Synuclein, Alpha-Synuclein and Double Alpha/Gamma-Synuclein Null Mutant Mice. J. Neurochem. 2004, 89, 1126–1136. [Google Scholar] [CrossRef]
- Landry, M.; Moreno, A.; Patry, S.; Potvin, S.; Lemasson, M. Current Practices of Electroconvulsive Therapy in Mental Disorders: A Systematic Review and Meta-Analysis of Short and Long-Term Cognitive Effects. J. ECT 2021, 37, 119. [Google Scholar] [CrossRef]
- Yoo, S.-S.; Bystritsky, A.; Lee, J.-H.; Zhang, Y.; Fischer, K.; Min, B.-K.; McDannold, N.J.; Pascual-Leone, A.; Jolesz, F.A. Focused Ultrasound Modulates Region-Specific Brain Activity. Neuroimage 2011, 56, 1267–1275. [Google Scholar] [CrossRef]
- Mahdavi, K.; Jordan, K.; Haroon, J.; Habelhah, B.; Rindner, E.; Surya, R.; Venkatraman, V.; Becerra, S.; Spivak, N.; Bystritsky, A.; et al. A Pilot Study of Low-Intensity Focused Ultrasound for Treatment-Resistant Generalized Anxiety Disorder. Brain Stimul. 2023, 16, 369–370. [Google Scholar] [CrossRef]
- Wu, S.-K.; Tsai, C.-L.; Huang, Y.; Hynynen, K. Focused Ultrasound and Microbubbles-Mediated Drug Delivery to Brain Tumor. Pharmaceutics 2020, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Legon, W.; Sato, T.F.; Opitz, A.; Mueller, J.; Barbour, A.; Williams, A.; Tyler, W.J. Transcranial Focused Ultrasound Modulates the Activity of Primary Somatosensory Cortex in Humans. Nat. Neurosci. 2014, 17, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jiang, Y.; Lan, L.; Ge, X.; Cheng, R.; Zhan, Y.; Chen, G.; Shi, L.; Wang, R.; Zheng, N.; et al. Optically-Generated Focused Ultrasound for Noninvasive Brain Stimulation with Ultrahigh Precision. Light Sci. Appl. 2022, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Natera-Villalba, E.; Matarazzo, M.; Martinez-Fernandez, R. Update in the Clinical Application of Focused Ultrasound. Curr. Opin. Neurol. 2022, 35, 525. [Google Scholar] [CrossRef] [PubMed]
- Krishna, V.; Fishman, P.S.; Eisenberg, H.M.; Kaplitt, M.; Baltuch, G.; Chang, J.W.; Chang, W.-C.; Martinez Fernandez, R.; del Alamo, M.; Halpern, C.H.; et al. Trial of Globus Pallidus Focused Ultrasound Ablation in Parkinson’s Disease. N. Engl. J. Med. 2023, 388, 683–693. [Google Scholar] [CrossRef]
- Mahoney, J.J.; Thompson Lake, D.G.Y.; Ranjan, M.; Marton, J.L.; Carpenter, J.S.; Zheng, W.; Berry, J.H.; Farmer, D.L.; D’Haese, P.F.; Finomore, V.S.; et al. Low-Intensity Focused Ultrasound Targeting the Bilateral Nucleus Accumbens as a Potential Treatment for Substance Use Disorder: A First-in-Human Report. Biol. Psychiatry, 2023; in press. [Google Scholar] [CrossRef]
- Rezai, A.R.; Ranjan, M.; D’Haese, P.-F.; Haut, M.W.; Carpenter, J.; Najib, U.; Mehta, R.I.; Chazen, J.L.; Zibly, Z.; Yates, J.R.; et al. Noninvasive Hippocampal Blood-Brain Barrier Opening in Alzheimer’s Disease with Focused Ultrasound. Proc. Natl. Acad. Sci. USA 2020, 117, 9180–9182. [Google Scholar] [CrossRef]
- Rezai, A.R.; Ranjan, M.; Haut, M.W.; Carpenter, J.; D’Haese, P.-F.; Mehta, R.I.; Najib, U.; Wang, P.; Claassen, D.O.; Chazen, J.L.; et al. Focused Ultrasound-Mediated Blood-Brain Barrier Opening in Alzheimer’s Disease: Long-Term Safety, Imaging, and Cognitive Outcomes. J. Neurosurg. 2023, 139, 275–283. [Google Scholar] [CrossRef]
- Liu, X.; Naomi, S.S.M.; Sharon, W.L.; Russell, E.J. The Applications of Focused Ultrasound (FUS) in Alzheimer’s Disease Treatment: A Systematic Review on Both Animal and Human Studies. Aging Dis. 2022, 12, 1977–2002. [Google Scholar] [CrossRef]
- Gasca-Salas, C.; Fernández-Rodríguez, B.; Pineda-Pardo, J.A.; Rodríguez-Rojas, R.; Obeso, I.; Hernández-Fernández, F.; del Álamo, M.; Mata, D.; Guida, P.; Ordás-Bandera, C.; et al. Blood-Brain Barrier Opening with Focused Ultrasound in Parkinson’s Disease Dementia. Nat. Commun. 2021, 12, 779. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Pineda-Pardo, J.A.; Inoue, K.-I.; Gasca-Salas, C.; Balzano, T.; Del Rey, N.L.-G.; Reinares-Sebastián, A.; Esteban-García, N.; Rodríguez-Rojas, R.; Márquez, R.; et al. BBB Opening with Focused Ultrasound in Nonhuman Primates and Parkinson’s Disease Patients: Targeted AAV Vector Delivery and PET Imaging. Sci. Adv. 2023, 9, eadf4888. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Feng, L.; Liu, S.; Zhou, X.; Yin, T.; Liu, Z.; Yang, Z. Transcranial Magneto-Acoustic Stimulation Improves Neuroplasticity in Hippocampus of Parkinson’s Disease Model Mice. Neurotherapeutics 2019, 16, 1210–1224. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Niu, L.; Meng, L.; Lin, Z.; Zou, J.; Xia, X.; Huang, X.; Zhou, W.; Bian, T.; Zheng, H. Noninvasive Ultrasound Deep Brain Stimulation for the Treatment of Parkinson’s Disease Model Mouse. Research 2019, 2019, 1748489. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Niu, L.; Xia, X.; Lin, Z.; Liu, X.; Su, M.; Guo, R.; Meng, L.; Zheng, H. Wearable Ultrasound Improves Motor Function in an MPTP Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2019, 66, 3006–3013. [Google Scholar] [CrossRef]
- Xu, T.; Lu, X.; Peng, D.; Wang, G.; Chen, C.; Liu, W.; Wu, W.; Mason, T.J. Ultrasonic Stimulation of the Brain to Enhance the Release of Dopamine—A Potential Novel Treatment for Parkinson’s Disease. Ultrason. Sonochem. 2020, 63, 104955. [Google Scholar] [CrossRef]
- Zhou, H.; Meng, L.; Xia, X.; Lin, Z.; Zhou, W.; Pang, N.; Bian, T.; Yuan, T.; Niu, L.; Zheng, H. Transcranial Ultrasound Stimulation Suppresses Neuroinflammation in a Chronic Mouse Model of Parkinson’s Disease. IEEE Trans. Biomed. Eng. 2021, 68, 3375–3387. [Google Scholar] [CrossRef]
- Song, W.-S.; Sung, C.-Y.; Ke, C.-H.; Yang, F.-Y. Anti-Inflammatory and Neuroprotective Effects of Transcranial Ultrasound Stimulation on Parkinson’s Disease. Ultrasound Med. Biol. 2022, 48, 265–274. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, D.; Zhao, Y.; Yuan, Y.; Wang, W.; Wu, S.; Liang, X.; Wang, Z.; Liu, L. Assessment of Neuroprotective Effects of Low-Intensity Transcranial Ultrasound Stimulation in a Parkinson’s Disease Rat Model by Fractional Anisotropy and Relaxation Time T2∗ Value. Front. Neurosci. 2021, 15, 590354. [Google Scholar] [CrossRef]
- Haacke, E.M.; Cheng, N.Y.C.; House, M.J.; Liu, Q.; Neelavalli, J.; Ogg, R.J.; Khan, A.; Ayaz, M.; Kirsch, W.; Obenaus, A. Imaging Iron Stores in the Brain Using Magnetic Resonance Imaging. Magn. Reson. Imaging 2005, 23, 1–25. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, Y.; Li, X. Theoretical Analysis of Transcranial Magneto-Acoustical Stimulation with Hodgkin-Huxley Neuron Model. Front. Comput. Neurosci. 2016, 10, 35. [Google Scholar] [CrossRef]
- Allen, S.J.; Watson, J.J.; Shoemark, D.K.; Barua, N.U.; Patel, N.K. GDNF, NGF and BDNF as Therapeutic Options for Neurodegeneration. Pharmacol. Ther. 2013, 138, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Poduslo, J.F.; Curran, G.L. Permeability at the Blood-Brain and Blood-Nerve Barriers of the Neurotrophic Factors: NGF, CNTF, NT-3, BDNF. Mol. Brain Res. 1996, 36, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.-H.; Ting, C.-Y.; Lin, C.-Y.; Chan, H.-L.; Chang, Y.-C.; Chen, Y.-Y.; Liu, H.-L.; Yeh, C.-K. Noninvasive, Targeted and Non-Viral Ultrasound-Mediated GDNF-Plasmid Delivery for Treatment of Parkinson’s Disease. Sci. Rep. 2016, 6, 19579. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Hsieh, H.-Y.; Chen, C.-M.; Wu, S.-R.; Tsai, C.-H.; Huang, C.-Y.; Hua, M.-Y.; Wei, K.-C.; Yeh, C.-K.; Liu, H.-L. Non-Invasive, Neuron-Specific Gene Therapy by Focused Ultrasound-Induced Blood-Brain Barrier Opening in Parkinson’s Disease Mouse Model. J. Control. Release 2016, 235, 72–81. [Google Scholar] [CrossRef]
- Mead, B.P.; Kim, N.; Miller, G.W.; Hodges, D.; Mastorakos, P.; Klibanov, A.L.; Mandell, J.W.; Hirsh, J.; Suk, J.S.; Hanes, J.; et al. Novel Focused Ultrasound Gene Therapy Approach Noninvasively Restores Dopaminergic Neuron Function in a Rat Parkinson’s Disease Model. Nano Lett. 2017, 17, 3533–3542. [Google Scholar] [CrossRef]
- Yue, P.; Gao, L.; Wang, X.; Ding, X.; Teng, J. Ultrasound-Triggered Effects of the Microbubbles Coupled to GDNF- and Nurr1-Loaded PEGylated Liposomes in a Rat Model of Parkinson’s Disease. J. Cell. Biochem. 2018, 119, 4581–4591. [Google Scholar] [CrossRef]
- Karakatsani, M.E.; Wang, S.; Samiotaki, G.; Kugelman, T.; Olumolade, O.O.; Acosta, C.; Sun, T.; Han, Y.; Kamimura, H.A.S.; Jackson-Lewis, V.; et al. Amelioration of the Nigrostriatal Pathway Facilitated by Ultrasound-Mediated Neurotrophic Delivery in Early Parkinson’s Disease. J. Control. Release 2019, 303, 289–301. [Google Scholar] [CrossRef]
- Decressac, M.; Volakakis, N.; Björklund, A.; Perlmann, T. NURR1 in Parkinson Disease—From Pathogenesis to Therapeutic Potential. Nat. Rev. Neurol. 2013, 9, 629–636. [Google Scholar] [CrossRef]
- Long, L.; Cai, X.; Guo, R.; Wang, P.; Wu, L.; Yin, T.; Liao, S.; Lu, Z. Treatment of Parkinson’s Disease in Rats by Nrf2 Transfection Using MRI-Guided Focused Ultrasound Delivery of Nanomicrobubbles. Biochem. Biophys. Res. Commun. 2017, 482, 75–80. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Huang, R.-Y.; Liao, E.-C.; Lin, Y.-C.; Ho, Y.-J.; Chang, C.-W.; Chan, H.-L.; Huang, Y.-Z.; Hsieh, T.-H.; Fan, C.-H.; et al. A Preliminary Study of Parkinson’s Gene Therapy via Sono-Magnetic Sensing Gene Vector for Conquering Extra/Intracellular Barriers in Mice. Brain Stimul. 2020, 13, 786–799. [Google Scholar] [CrossRef]
- Ji, R.; Smith, M.; Niimi, Y.; Karakatsani, M.E.; Murillo, M.F.; Jackson-Lewis, V.; Przedborski, S.; Konofagou, E.E. Focused Ultrasound Enhanced Intranasal Delivery of Brain Derived Neurotrophic Factor Produces Neurorestorative Effects in a Parkinson’s Disease Mouse Model. Sci. Rep. 2019, 9, 19402. [Google Scholar] [CrossRef] [PubMed]
- Itoh, N.; Ohta, H. Roles of FGF20 in Dopaminergic Neurons and Parkinson’s Disease. Front. Mol. Neurosci. 2013, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Xie, J.; Guo, K.; Zhang, X.; Xia, F.; Zhao, X.; Song, L.; Zhuge, D.; Li, X.; Zhao, Y.; et al. Efficient Treatment of Parkinson’s Disease Using Ultrasonography-Guided RhFGF20 Proteoliposomes. Drug Deliv. 2018, 25, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Shimada, H.; Yoshimura, N.; Tsuji, A.; Kunisada, T. Differentiation of Dopaminergic Neurons from Human Embryonic Stem Cells: Modulation of Differentiation by FGF-20. J. Biosci. Bioeng. 2009, 107, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, I.J.; Boshoff, E.L.; Duty, S. Fibroblast Growth Factor-20 Protects against Dopamine Neuron Loss in Vitro and Provides Functional Protection in the 6-Hydroxydopamine-Lesioned Rat Model of Parkinson’s Disease. Neuropharmacology 2012, 63, 1268–1277. [Google Scholar] [CrossRef]
- Yan, Y.; Chen, Y.; Liu, Z.; Cai, F.; Niu, W.; Song, L.; Liang, H.; Su, Z.; Yu, B.; Yan, F. Brain Delivery of Curcumin Through Low-Intensity Ultrasound-Induced Blood-Brain Barrier Opening via Lipid-PLGA Nanobubbles. Int. J. Nanomed. 2021, 16, 7433–7447. [Google Scholar] [CrossRef]
- Zhang, N.; Yan, F.; Liang, X.; Wu, M.; Shen, Y.; Chen, M.; Xu, Y.; Zou, G.; Jiang, P.; Tang, C.; et al. Localized Delivery of Curcumin into Brain with Polysorbate 80-Modified Cerasomes by Ultrasound-Targeted Microbubble Destruction for Improved Parkinson’s Disease Therapy. Theranostics 2018, 8, 2264–2277. [Google Scholar] [CrossRef]
- Yavarpour-Bali, H.; Ghasemi-Kasman, M.; Pirzadeh, M. Curcumin-Loaded Nanoparticles: A Novel Therapeutic Strategy in Treatment of Central Nervous System Disorders. Int. J. Nanomed. 2019, 14, 4449–4460. [Google Scholar] [CrossRef]
- Pandey, N.; Strider, J.; Nolan, W.C.; Yan, S.X.; Galvin, J.E. Curcumin Inhibits Aggregation of Alpha-Synuclein. Acta Neuropathol. 2008, 115, 479–489. [Google Scholar] [CrossRef]
- Feng, Y.; An, R.; Zhang, Y.; Chen, M.; Wang, L.; Duan, Y.; Xing, C. AHNAK-Modified Microbubbles for the Intracranial Delivery of Triptolide: In-Vitro and in-Vivo Investigations. Int. J. Pharm. 2022, 629, 122351. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Gong, X.; Wang, L.; Liu, M.; Liu, Y.; Fu, X.; Wang, W.; Zhang, T.; Wang, X. Triptolide Promotes the Clearance of α-Synuclein by Enhancing Autophagy in Neuronal Cells. Mol. Neurobiol. 2017, 54, 2361–2372. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.-L.; Yang, Y.-X.; Ding, J.; Li, Y.-C.; Miao, Z.-H. Triptolide: Structural Modifications, Structure-Activity Relationships, Bioactivities, Clinical Development and Mechanisms. Nat. Prod. Rep. 2012, 29, 457–475. [Google Scholar] [CrossRef]
- Wang, Y.; Luo, K.; Li, J.; Liao, Y.; Liao, C.; Chen, W.-S.; Chen, M.; Ao, L. Focused Ultrasound Promotes the Delivery of Gastrodin and Enhances the Protective Effect on Dopaminergic Neurons in a Mouse Model of Parkinson’s Disease. Front. Cell. Neurosci. 2022, 16, 884788. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, X.; Wang, H.; Qu, M. Gastrodin Attenuates Pentylenetetrazole-Induced Seizures by Modulating the Mitogen-Activated Protein Kinase-Associated Inflammatory Responses in Mice. Neurosci. Bull. 2016, 33, 264–272. [Google Scholar] [CrossRef]
- Xhima, K.; Nabbouh, F.; Hynynen, K.; Aubert, I.; Tandon, A. Non-Invasive Delivery of an α-Synuclein Gene Silencing Vector with MR-Guided Focused Ultrasound. Mov. Disord. 2018, 33, 1567–1579. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Huang, C.-Y.; Chen, C.-M.; Liu, H.-L. Focused Ultrasound-Induced Blood-Brain Barrier Opening Enhanced α-Synuclein Expression in Mice for Modeling Parkinson’s Disease. Pharmaceutics 2022, 14, 444. [Google Scholar] [CrossRef]
- Kim, T.; Kim, H.J.; Choi, W.; Lee, Y.M.; Pyo, J.H.; Lee, J.; Kim, J.; Kim, J.; Kim, J.-H.; Kim, C.; et al. Deep Brain Stimulation by Blood-Brain-Barrier-Crossing Piezoelectric Nanoparticles Generating Current and Nitric Oxide under Focused Ultrasound. Nat. Biomed. Eng. 2023, 7, 149–163. [Google Scholar] [CrossRef]
- Kovacs, Z.I.; Kim, S.; Jikaria, N.; Qureshi, F.; Milo, B.; Lewis, B.K.; Bresler, M.; Burks, S.R.; Frank, J.A. Disrupting the Blood-Brain Barrier by Focused Ultrasound Induces Sterile Inflammation. Proc. Natl. Acad. Sci. USA 2017, 114, E75–E84. [Google Scholar] [CrossRef]
- Jordão, J.F.; Thévenot, E.; Markham-Coultes, K.; Scarcelli, T.; Weng, Y.-Q.; Xhima, K.; O’Reilly, M.; Huang, Y.; McLaurin, J.; Hynynen, K.; et al. Amyloid-β Plaque Reduction, Endogenous Antibody Delivery and Glial Activation by Brain-Targeted, Transcranial Focused Ultrasound. Exp. Neurol. 2013, 248, 16–29. [Google Scholar] [CrossRef]
- Leinenga, G.; Götz, J. Scanning Ultrasound Removes Amyloid-β and Restores Memory in an Alzheimer’s Disease Mouse Model. Sci. Transl. Med. 2015, 7, 278ra33. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.; Hynynen, K. Acute Inflammatory Response Following Increased Blood-Brain Barrier Permeability Induced by Focused Ultrasound Is Dependent on Microbubble Dose. Theranostics 2017, 7, 3989–4000. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Chaney, A.M.; Carlson, M.L.; Jackson, I.M.; Rao, A.; James, M.L. Neuroinflammation PET Imaging: Current Opinion and Future Directions. J. Nucl. Med. 2020, 61, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Lavisse, S.; Goutal, S.; Wimberley, C.; Tonietto, M.; Bottlaender, M.; Gervais, P.; Kuhnast, B.; Peyronneau, M.-A.; Barret, O.; Lagarde, J.; et al. Increased Microglial Activation in Patients with Parkinson Disease Using [18F]-DPA714 TSPO PET Imaging. Park. Relat. Disord. 2021, 82, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Sochocka, M.; Diniz, B.S.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef]
- Shah, F.; Hume, S.P.; Pike, V.W.; Ashworth, S.; McDermott, J. Synthesis of the Enantiomers of [N-Methyl-11C]PK 11195 and Comparison of Their Behaviours as Radioligands for PK Binding Sites in Rats. Nucl. Med. Biol. 1994, 21, 573–581. [Google Scholar] [CrossRef]
- Molendijk, M.L.; de Kloet, E.R. Forced Swim Stressor: Trends in Usage and Mechanistic Consideration. Eur. J. Neurosci. 2022, 55, 2813–2831. [Google Scholar] [CrossRef]
- Stephan, M.; Volkmann, P.; Rossner, M.J. Assessing Behavior and Cognition in Rodents, Nonhuman Primates, and Humans: Where Are the Limits of Translation? Dialogues Clin. Neurosci. 2019, 21, 249–259. [Google Scholar] [CrossRef]
- Rentsch, P.; Stayte, S.; Morris, G.P.; Vissel, B. Time Dependent Degeneration of the Nigrostriatal Tract in Mice with 6-OHDA Lesioned Medial Forebrain Bundle and the Effect of Activin A on l-Dopa Induced Dyskinesia. BMC Neurosci. 2019, 20, 5. [Google Scholar] [CrossRef]
- Masini, D.; Plewnia, C.; Bertho, M.; Scalbert, N.; Caggiano, V.; Fisone, G. A Guide to the Generation of a 6-Hydroxydopamine Mouse Model of Parkinson’s Disease for the Study of Non-Motor Symptoms. Biomedicines 2021, 9, 598. [Google Scholar] [CrossRef]
- Meredith, G.E.; Rademacher, D.J. MPTP Mouse Models of Parkinson’s Disease: An Update. J. Park. Dis. 2011, 1, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Navarro, J.A.; Casarejos, M.J.; Menéndez, J.; Solano, R.M.; Rodal, I.; Gómez, A.; de Yébenes, J.G.; Mena, M.A. Mortality, Oxidative Stress and Tau Accumulation during Ageing in Parkin Null Mice. J. Neurochem. 2007, 103, 98–114. [Google Scholar] [CrossRef] [PubMed]
- Correia Guedes, L.; Mestre, T.; Outeiro, T.F.; Ferreira, J.J. Are Genetic and Idiopathic Forms of Parkinson’s Disease the Same Disease? J. Neurochem. 2020, 152, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Blandini, F.; Armentero, M.-T. Animal Models of Parkinson’s Disease. FEBS J. 2012, 279, 1156–1166. [Google Scholar] [CrossRef]
- Sarre, S.; Yuan, H.; Jonkers, N.; Van Hemelrijck, A.; Ebinger, G.; Michotte, Y. In Vivo Characterization of Somatodendritic Dopamine Release in the Substantia Nigra of 6-Hydroxydopamine-Lesioned Rats. J. Neurochem. 2004, 90, 29–39. [Google Scholar] [CrossRef]
- Stott, S.R.W.; Barker, R.A. Time Course of Dopamine Neuron Loss and Glial Response in the 6-OHDA Striatal Mouse Model of Parkinson’s Disease. Eur. J. Neurosci. 2014, 39, 1042–1056. [Google Scholar] [CrossRef]
- Pol, F.; Salehinejad, M.A.; Baharlouei, H.; Nitsche, M.A. The Effects of Transcranial Direct Current Stimulation on Gait in Patients with Parkinson’s Disease: A Systematic Review. Transl. Neurodegener. 2021, 10, 22. [Google Scholar] [CrossRef]
- Takamiya, A.; Seki, M.; Kudo, S.; Yoshizaki, T.; Nakahara, J.; Mimura, M.; Kishimoto, T. Electroconvulsive Therapy for Parkinson’s Disease: A Systematic Review and Meta-Analysis. Mov. Disord. 2021, 36, 50–58. [Google Scholar] [CrossRef]
- Sveinbjornsdottir, S. The Clinical Symptoms of Parkinson’s Disease. J. Neurochem. 2016, 139, 318–324. [Google Scholar] [CrossRef]
- Andrade, C.; Arumugham, S.S.; Thirthalli, J. Adverse Effects of Electroconvulsive Therapy. Psychiatr. Clin. N. Am. 2016, 39, 513–530. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ugawa, Y. Adverse Events of TDCS and TACS: A Review. Clin. Neurophysiol. Pract. 2016, 2, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Loo, C.K.; McFarquhar, T.F.; Mitchell, P.B. A Review of the Safety of Repetitive Transcranial Magnetic Stimulation as a Clinical Treatment for Depression. Int. J. Neuropsychopharmacol. 2008, 11, 131–147. [Google Scholar] [CrossRef]
- Wassermann, E.M. Side Effects of Repetitive Transcranial Magnetic Stimulation. Depress. Anxiety 2000, 12, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Magsood, H.; Syeda, F.; Holloway, K.; Carmona, I.C.; Hadimani, R.L. Safety Study of Combination Treatment: Deep Brain Stimulation and Transcranial Magnetic Stimulation. Front. Hum. Neurosci. 2020, 14, 123. [Google Scholar] [CrossRef] [PubMed]
- Eun, J.D.; Bang, Y.M.; Youn, J.; Cho, J.W.; Kim, Y.-H.; Chang, W.H. Feasibility of Transcranial Direct Current Stimulation in Patients with Deep Brain Stimulation: A Case Report. Brain NeuroRehabil. 2020, 13, e13. [Google Scholar] [CrossRef]
- Ducharme, S.; Flaherty, A.W.; Seiner, S.J.; Dougherty, D.D.; Morales, O.G. Temporary Interruption of Deep Brain Stimulation for Parkinson’s Disease during Outpatient Electroconvulsive Therapy for Major Depression: A Novel Treatment Strategy. J. Neuropsychiatry Clin. Neurosci. 2011, 23, 194–197. [Google Scholar] [CrossRef]
- Cunningham, M.G.; Yadollahikhales, G.; Vitaliano, G.; van Horne, C. Administration of Electroconvulsive Therapy for Depression Associated with Deep Brain Stimulation in a Patient with Post-Traumatic Parkinson’s Disease: A Case Study. BMC Psychiatry 2016, 16, 399. [Google Scholar] [CrossRef]
- Volkaerts, L.; Roels, R.; Bouckaert, F. Motor Function Improvement After Electroconvulsive Therapy in a Parkinson’s Disease Patient With Deep Brain Stimulator. J. ECT 2020, 36, 66–68. [Google Scholar] [CrossRef]
- Bailine, S.; Kremen, N.; Kohen, I.; Linder, H.; Schwartz, G.J.; Mogilner, A.Y.; Pourfar, M. Bitemporal Electroconvulsive Therapy for Depression in a Parkinson Disease Patient with a Deep-Brain Stimulator. J. ECT 2008, 24, 171–172. [Google Scholar] [CrossRef]
- Nasr, S.; Murillo, A.; Katariwala, N.; Mothkur, V.; Wendt, B. Case Report of Electroconvulsive Therapy in a Patient with Parkinson Disease Concomitant with Deep Brain Stimulation. J. ECT 2011, 27, 89–90. [Google Scholar] [CrossRef]
- Sarica, C.; Fomenko, A.; Nankoo, J.-F.; Darmani, G.; Vetkas, A.; Yamamoto, K.; Lozano, A.M.; Chen, R. Toward Focused Ultrasound Neuromodulation in Deep Brain Stimulator Implanted Patients: Ex-Vivo Thermal, Kinetic and Targeting Feasibility Assessment. Brain Stimul. 2022, 15, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Jackson-Lewis, V.; Blesa, J.; Przedborski, S. Animal Models of Parkinson’s Disease. Park. Relat. Disord. 2012, 18, S183–S185. [Google Scholar] [CrossRef] [PubMed]


| TMS Type | Stimulated Regions | Primary Symptom Relief |
|---|---|---|
| hf-rTMS | Frontal, prefrontal, M1, DLPFC, SMA, inferior frontal gyrus, dorsal premotor cortex, occipital cortex |
|
| lf-rTMS | Vertex, frontal, prefrontal, M1, DLPFC, SMA |
|
| cTBS | SMA |
|
| iTBS | M1, DLPFC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muksuris, K.; Scarisbrick, D.M.; Mahoney, J.J., III; Cherkasova, M.V. Noninvasive Neuromodulation in Parkinson’s Disease: Insights from Animal Models. J. Clin. Med. 2023, 12, 5448. https://doi.org/10.3390/jcm12175448
Muksuris K, Scarisbrick DM, Mahoney JJ III, Cherkasova MV. Noninvasive Neuromodulation in Parkinson’s Disease: Insights from Animal Models. Journal of Clinical Medicine. 2023; 12(17):5448. https://doi.org/10.3390/jcm12175448
Chicago/Turabian StyleMuksuris, Katherine, David M. Scarisbrick, James J. Mahoney, III, and Mariya V. Cherkasova. 2023. "Noninvasive Neuromodulation in Parkinson’s Disease: Insights from Animal Models" Journal of Clinical Medicine 12, no. 17: 5448. https://doi.org/10.3390/jcm12175448
APA StyleMuksuris, K., Scarisbrick, D. M., Mahoney, J. J., III, & Cherkasova, M. V. (2023). Noninvasive Neuromodulation in Parkinson’s Disease: Insights from Animal Models. Journal of Clinical Medicine, 12(17), 5448. https://doi.org/10.3390/jcm12175448