
The Impact of Hormesis, Neuronal Stress Response, and Reproduction, upon Clinical Aging: A Narrative Review



Abstract
1. Introduction and Methods
“This definition implies that, with the passage of time and for a variety of causative factors, humans are subjected to damage which is not properly repaired. As a consequence, there is degeneration and loss of utility at all levels (molecular, cellular, tissue, organismic, and societal) with a resulting failure of the normal function of a human. In other words, it is a chronologically-dependent erosion of our functions, which makes it increasingly difficult for us to manage and operate within a given, always-changing environment”.
2. Results and Discussion
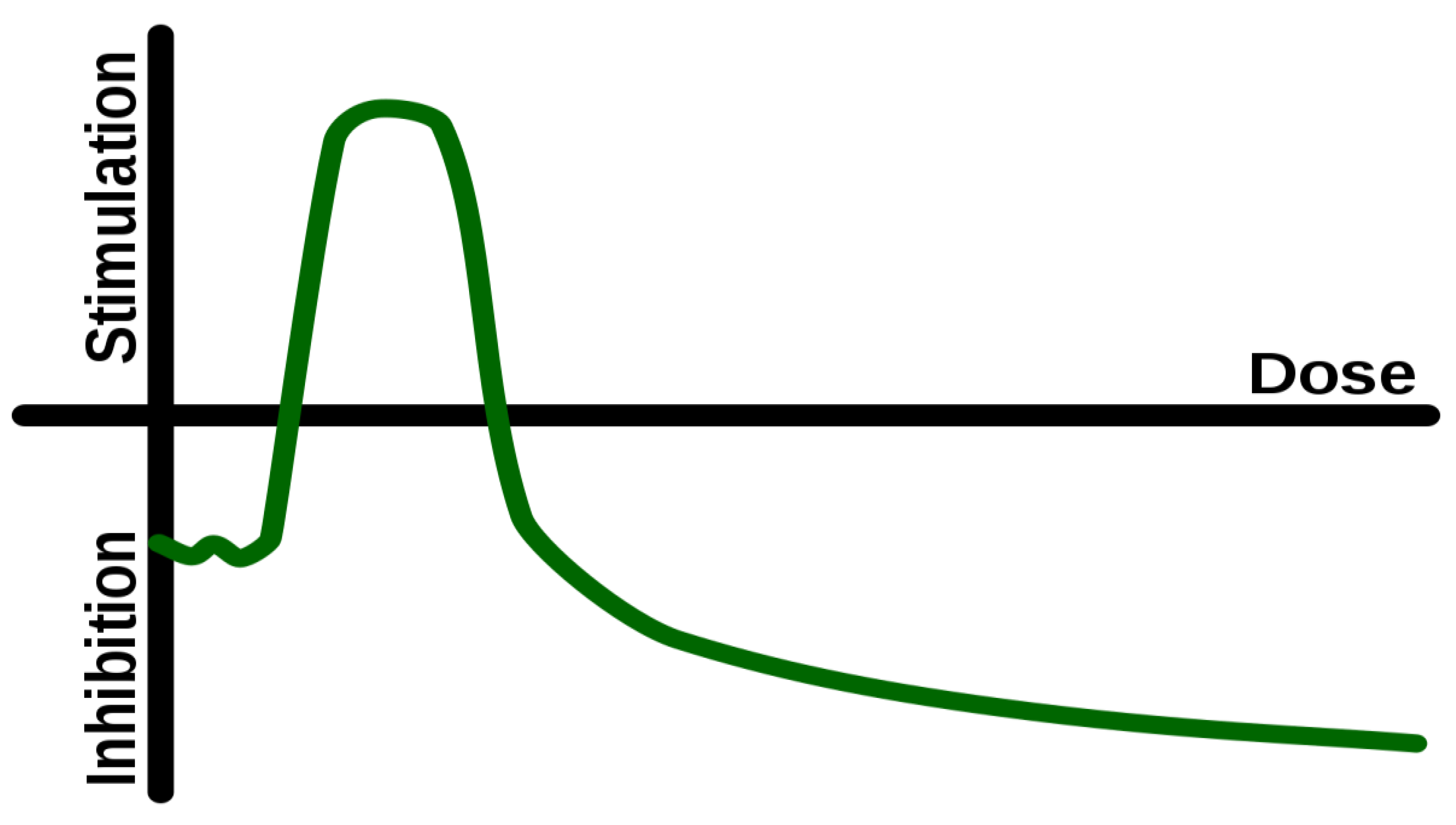
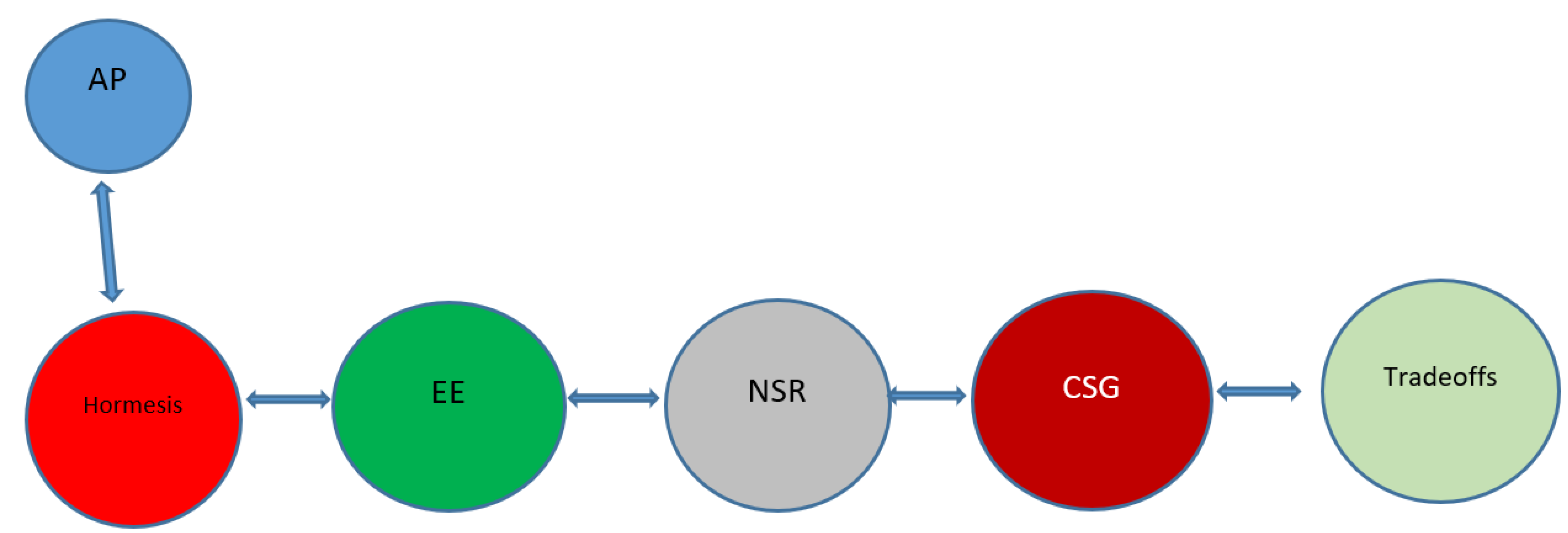
2.1. Hormesis
- Rattan explains the concept of ‘homeodynamic space’ as encompassing “three characteristics: stress response, damage control, and constant remodeling, which provide measurable biomarkers reflecting the survival ability, robustness, and resilience of a biological system. A biological definition of health thus involves measures of functionality, tolerance and adaptation” [21].
- The homeodynamic space refers to the range of physiological states and processes that an organism can tolerate and maintain without suffering harm or death. This range is determined by the organism’s capacity to adapt to and cope with stress [22].
- The concept of homeodynamic space is related to the concept of homeostasis. However, while homeostasis refers to a narrow range of physiological states that are optimal for survival, homeodynamic space refers to a much broader range of physiological states and processes that an organism can tolerate. It highlights the dynamic nature of physiological systems and the importance of resilience and adaptability in maintaining health and well-being.
A basic characteristic of a hormetic event is the novelty of information. Hormesis is present when the challenge is of sufficient magnitude and appropriate quality as to satisfy the definition of ‘novelty’. Novelty is defined as ‘the quality of being new, original, or unusual’, and this includes both unfamiliarity and unconventionality. Routine and monotony generally do not invoke a hormetic response. The assessment and response to the new challenge leads to adaptation and thus, eventually, improvement of function within a particular environment (the environment where the challenges have originated from). Therefore, when a certain stimulus appears ‘novel’ to us, then it is likely that this stimulus is eliciting a hormetic response. When the stimulus appears boring or monotonous, it is less likely that hormesis is taking place.
2.2. Autophagy and Hormesis
- Exercise. This can enhance autophagy in liver, muscles, pancreas and adipose tissue, as well as in the brain [29].
- Intermittent fasting (IF) is a nutritional hormetic stress. Alirezaei et al. [33] conducted a study to investigate the effects of food restriction and short-term fasting on autophagy. Their findings revealed that food restriction induces autophagy in mouse livers, challenging the conventional belief of the brain’s metabolic privilege. Moreover, their research suggests that sporadic fasting could be a cost-effective approach to promote a therapeutic neuronal response. In a separate study, Pietrocola et al. [34] emphasized the significance of autophagy in cancer treatment. They highlighted that impairment of autophagy reduces the effectiveness of chemotherapy and radiotherapy. These findings underscore the importance of understanding autophagic mechanisms to enhance cancer treatment strategies. Additionally, Kim and Lemasters [35] observed the occurrence of autophagy in liver cells during fasting, providing further insights into its role in cellular recycling. Their study demonstrated that liver cells form phagophores and autophagosomes, which encapsulate and capture mitochondria for recycling. This process leads to the breakdown of mitochondria and their contents, including DNA. In addition to physical stimuli, autophagy can be modulated by hormetins, i.e., substances that can induce health-beneficial physiological hormesis [36] and this is an appropriate opportunity to discuss some more details of hormetins (Table 1).
2.3. Environmental Enrichment
2.4. Neuronal Stress Response
2.5. Digital Information, Cognition, and Neuronal Stress Response
“…evidence that video game interventions could be considered for the elderly for improving performance and cognitive function, especially general cognitive scores and processing speed. Games with better interactivity and visual stimulation have better curative effects…”.
2.6. The Bidirectional Communication (Cross-Talk) between Neurons and the Germline
- Germline cells have the innate capacity to become neurons following suitable natural (or artificial) reprogramming by transcription factors, even though there are several conserved mechanisms that safeguard against this. This is an intriguing situation because it shows the direct relationship between the germline and the brain [103]. In addition, multipotent neural and glial precursors can be derived from multipotent adult germ line stem cells [104]. These multipotent neural precursors are able to mature and integrate within the existing neural network. It is necessary to mention that, although these effects have been found in experiments conducted in vitro, clinical tests in vivo are still lacking. It is, however, remarkable to realize that the germline acts as a source of fully functional neurons [105,106,107].
- A more detailed direct communication pathway between germline cells and the soma has been studied by Levi-Feber et al. [110] who showed that this pathway depends on the endoplasmic reticulum stress factor inositol requiring enzyme-1 (IRE-1).
- On certain situations, ectopic germline cells can be found in the brain, and could contribute to altered neuronal development, resulting in neurodevelopmental disorders. This indicates not only the close relationship between neurons and germ cells, but also the continual struggle for equilibrium, between these two [111].
- Furthermore, there is another fact that underlines the close relationship between neurons and germ cells. Progesterone, which modulates sperm function, acts (via intermediate steps) by interacting with “a sperm membrane receptor which resembles the neuronal GABA(A) receptor” in the brain [112].
- It was shown that eradication of germ cells in Drosophila, has a positive impact on its lifespan, possibly through modulation of the nutrient sensing insulin/insulin-like (IIS) growth factor signaling [113]. This strengthens the general argument that that somatic lifespan is under the control of the germ line, and vice versa.
- The repressor element 1-silencing transcription factor REST which modulates multipotent stem cells, is present in testes, but, intriguingly, regulates target genes in neurons [114]. REST activity has been associated with cognitive impairment and dementia, whereas a potent activity of REST is associated with modulating the balance of neuroprotection vs. neurodegeneration—i.e., acts in a hormetic way [115,116]. Thus, there exist a conserved mechanism of modulation of neural development regulated by REST which is present in spermatogonial cells, indicating another possible mechanism of neuro-germline communication.
- It is known that the germline may influence the function of distant somatic cells, including neurons. For instance, germline stem cells influence proteostasis and thus prevent abnormal protein accumulation in neurons [117]. Thus, at least theoretically, the risk of neurodegenerative diseases is reduced.
- Spermidine is a natural polyamine compound with effects on heart disease, brain degeneration, cancer, and inflammation, among others. It also extends lifespan and health span [43], and it modulates autophagy both in the germline and in the neuron [118]. It was originally found in sperm and this begs the question: how and why does a compound in semen benefit the neuron? Spermidine is a mediator of the complex relationship that exists between neurons and the germline. The concentration of spermidine (apart from its high concentration in the sperm) is also high in the human brain. This must be because it has important actions to perform there [119]. It has positive actions on neuronal mitochondria [118], improves autophagy in neurons [120], and in germline stem cells [121], protects against synaptic degeneration [122] and exhibits general neuroprotective actions [123]. There are studies linking consumption of spermidine with a reduced risk of cognitive impairment in humans [124].
- DNA damage in germ cells increases resilience in somatic cells [91].
- Neuronal stress causes an increased demand (by neurons) for repair resources, which are diverted from the germline [90].
- A direct communication pathway between germline cells and the soma depends on the endoplasmic reticulum stress factor inositol requiring enzyme-1 (IRE-1) [110].
- Ectopic germline cells can be found in the brain, and could contribute to altered neuronal development, resulting in neurodevelopmental disorders [111].
- Progesterone, which modulates sperm function, acts by interacting with a membrane receptor which resembles the neuronal GABA(A) receptor in the brain [112].
- Eradication of germ cells in Drosophila, has a positive impact on its lifespan, possibly through modulation of the nutrient sensing insulin/insulin-like (IIS) growth factor signaling [113].
- The repressor element 1-silencing transcription factor (REST) which modulates multipotent stem cells, is present in both neonatal and adult testes, and regulates target genes in neurons [114].
- The germline may influence the function of distant somatic cells, including neurons. For instance, germline stem cells influence proteostasis and thus control abnormal protein accumulation in neurons [117].
2.7. Neurons vs. Germline
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kyriazis, M. Aging as “Time-Related Dysfunction”: A Perspective. Front. Med. 2020, 7, 371. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, M. Clinical Effects of a ‘Human-Computer’ Interaction. 21 June 2016. Available online: https://ssrn.com/abstract=2798529 (accessed on 20 June 2023).
- Digital Watch. Available online: https://dig.watch/trends/digital-and-environment (accessed on 20 June 2023).
- Germline: National Human Genome Research Institute. Available online: https://www.genome.gov/genetics-glossary/germ-line (accessed on 20 June 2023).
- Gaddy, M.A.; Kuang, S.; Alfhili, M.A.; Lee, M.H. The soma-germline communication: Implications for somatic and reproductive aging. BMB Rep. 2021, 54, 253–259. [Google Scholar] [CrossRef]
- Sharma, A. Transgenerational epigenetics: Integrating soma to germline communication with gametic inheritance. Mech. Ageing Dev. 2017, 163, 15–22. [Google Scholar] [CrossRef]
- Qi, W.; Xu, F.; Heimbucher, T.; Baumeister, R. Protection of germline immortality by the soma via a secreted endoribonuclease. BioEssays 2021, 43, 2100195. [Google Scholar] [CrossRef] [PubMed]
- Conine, C.C.; Rando, O.J. Soma-to-germline RNA communication. Nat. Rev. Genet. 2022, 23, 73–88. [Google Scholar] [CrossRef]
- Levi-Ferber, M.; Shalash, R.; Le-Thomas, A.; Salzberg, Y.; Shurgi, M.; Benichou, J.I.; Ashkenazi, A.; Henis-Korenblit, S. Neuronal regulated ire-1-dependent mRNA decay controls germline differentiation in Caenorhabditis elegans. Elife 2021, 10, e65644. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, M. Biological ageing and clinical consequences of modern technology. Biogerontology 2017, 18, 711–715. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. Defining hormesis. Hum. Exp. Toxicol. 2002, 21, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Rattan, S. Hormesis in aging. Ageing Res. Rev. 2008, 7, 63–78. [Google Scholar] [CrossRef]
- Santoro, A.; Martucci, M.; Conte, M.; Capri, M.; Franceschi, C.; Salvioli, S. Inflammaging, hormesis and the rationale for anti-aging strategies. Ageing Res. Rev. 2020, 64, 101142. [Google Scholar] [CrossRef]
- Nitti, M.; Marengo, B.; Furfaro, A.L.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C.; Traverso, N. Hormesis and Oxidative Distress: Pathophysiology of Reactive Oxygen Species and the Open Question of Antioxidant Modulation and Supplementation. Antioxidants 2022, 11, 1613. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, T.; Sun, Z. Hormesis in Health and Chronic Diseases. Trends Endocrinol. Metab. 2019, 30, 944–958. [Google Scholar] [CrossRef] [PubMed]
- Mattson, M.P. Hormesis defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Agathokleous, E.; Kitao, M.; Calabrese, E.J. Hormesis: Highly Generalizable and Beyond Laboratory. Trends Plant Sci. 2020, 25, 1076–1086. [Google Scholar] [CrossRef]
- Pande, S.; Raisuddin, S. The Underexplored Dimensions of Nutritional Hormesis. Curr. Nutr. Rep. 2022, 11, 386–394. [Google Scholar] [CrossRef]
- Demirovic, D.; Rattan, S.I. Establishing cellular stress response profiles as biomarkers of homeodynamics, health and hormesis. Exp. Gerontol. 2013, 48, 94–98. [Google Scholar] [CrossRef]
- Calabrese, E.J.; Mattson, M.P. How does hormesis impact biology, toxicology, and medicine? Aging. Mech. Dis. 2017, 3, 13. [Google Scholar] [CrossRef]
- Rattan, S.I.S. Biological Health and Homeodynamic Space. In Explaining Health Across the Sciences; Healthy Ageing and Longevity; Sholl, J., Rattan, S.I., Eds.; Springer: Cham, Switzerland, 2020; Volume 12. [Google Scholar]
- Rattan, S.I. Aging is not a disease: Implications for intervention. Aging Dis. 2014, 5, 196–202. [Google Scholar] [CrossRef]
- Mehdi, M.M.; Solanki, P.; Singh, P. Oxidative stress, antioxidants, hormesis and calorie restriction: The current perspective in the biology of aging. Arch. Gerontol. Geriatr. 2021, 95, 104413. [Google Scholar] [CrossRef]
- Toussaint, O.; Remacle, J.; Dierick, J.F.; Pascal, T.; Frippiat, C.; Royer, V.; Chainiaux, F. Approach of evolutionary theories of ageing, stress, senescence-like phenotypes, calorie restriction and hormesis from the view point of far-from-equilibrium thermodynamics. Mech. Ageing Dev. 2002, 123, 937–946. [Google Scholar] [CrossRef]
- Testa, G.; Biasi, F.; Poli, G.; Chiarpotto, E. Calorie restriction and dietary restriction mimetics: A strategy for improving healthy aging and longevity. Curr Pharm Des. 2014, 20, 2950–2977. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, M. Challenging Aging: The Anti-Senescence Effects of Hormesis, Environmental Enrichment, and Information Exposure; Frontiers in Aging Sciences; Bentham Science Publishers: Sharjah, United Arab Emirates, 2016; Volume 1, ISSN 2468-5933. [Google Scholar]
- Kyriazis, M. Systems neuroscience in focus: From the human brain to the global brain? Front. Syst. Neurosci. 2015, 9, 7. [Google Scholar] [CrossRef]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of Aging: An Autophagic Perspective. Front. Endocrinol. 2019, 9, 790. [Google Scholar] [CrossRef]
- He, C.; Sumpter, R.; Levine, B. Exercise induces autophagy in peripheral tissues and in the brain. Autophagy 2012, 8, 1548–1551. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiang, M.; Bi, Y.; Zhu, H.; Zhou, Z. Autophagy and Apoptosis act as partners to induce germ cell death after heat stress in mice. PLoS ONE 2012, 7, e41412. [Google Scholar] [CrossRef]
- Penke, B.; Bogár, F.; Crul, T.; Sántha, M.; Tóth, M.E.; Vígh, L. Heat Shock Proteins and Autophagy Pathways in Neuroprotection: From Molecular Bases to Pharmacological Interventions. Int. J. Mol. Sci. 2018, 19, 325. [Google Scholar] [CrossRef]
- Kumsta, C.; Chang, J.T.; Schmalz, J.; Hansen, M. Hormetic heat stress and HSF-1 induce autophagy to improve survival and proteostasis in C. elegans. Nat. Commun. 2017, 8, 14337. [Google Scholar] [CrossRef]
- Alirezaei, M.; Kemball, C.C.; Flynn, C.T.; Wood, M.R.; Whitton, J.; Kiosses, W.B. Short-term fasting induces profound neuronal autophagy. Autophagy 2010, 6, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Pol, J.; Vacchelli, E.; Rao, S.; Enot, D.P.; Baracco, E.E.; Levesque, S.; Castoldi, F.; Jacquelot, N.; Yamazaki, T.; et al. Caloric Restriction Mimetics Enhance Anticancer Immunosurveillance. Cancer Cell 2016, 30, 147–160. [Google Scholar] [CrossRef]
- Kim, I.; Lemasters, J.J. Mitochondrial degradation by autophagy (mitophagy) in GFP-LC3 transgenic hepatocytes during nutrient deprivation. Am. J. Physiol. Cell Physiol. 2011, 300, C308–C317. [Google Scholar] [CrossRef]
- Rattan, S.I.S. Hormetins as drugs for healthy aging. In Anti-Aging Drugs: From Basic Research to Clinical Practice; Vaiserman, M., Ed.; The Royal Society of Chemistry: London, UK, 2017; pp. 170–180. [Google Scholar]
- Arabit, J.G.J.; Elhaj, R.; Schriner, S.E.; Sevrioukov, E.A.; Jafari, M. Rhodiola rosea Improves Lifespan, Locomotion, and Neurodegeneration in a Drosophila melanogaster Model of Huntington’s Disease. Biomed. Res. Int. 2018, 2018, 6726874. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Simoneau, A.R.; Jafari, M.; Zi, X. Rhodiola rosea extracts and salidroside decrease the growth of bladder cancer cell lines via inhibition of the mTOR pathway and induction of autophagy. Mol. Carcinog. 2012, 51, 257–267. [Google Scholar] [CrossRef]
- Yu, C.; Yu, C.; Jing, S.; Li, H.; Jiang, E.; Ju, W.; Chen, J. Effects of Schisandra total lignin on autophagy and apoptosis of mouse brain aging induced by D-galactose. J. Jilin Univ. (Med. Ed.) 2014, 40, 1210–1215. [Google Scholar]
- Zhao, X.; Liu, C.; Xu, M.; Li, X.; Bi, K.; Jia, Y. Total Lignans of Schisandra chinensis Ameliorates Aβ1-42-Induced Neurodegeneration with Cognitive Impairment in Mice and Primary Mouse Neuronal Cells. PLoS ONE 2016, 11, e0152772. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Yi, H.K. Schisandrin C enhances mitochondrial biogenesis and autophagy in C2C12 skeletal muscle cells: Potential involvement of anti-oxidative mechanisms. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 197–206. [Google Scholar] [CrossRef]
- Sun, J.; Jing, S.; Jiang, R.; Wang, C.; Zhang, C.; Chen, J.; Li, H. Metabolomics study of the therapeutic mechanism of Schisandra chinensis lignans on aging rats induced by d-galactose. Clin. Interv. Aging 2018, 13, 829–841. [Google Scholar] [CrossRef]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359, eaan2788. [Google Scholar] [CrossRef] [PubMed]
- Sigrist, S.J.; Carmona-Gutierrez, D.; Gupta, V.K.; Bhukel, A.; Mertel, S.; Eisenberg, T.; Madeo, F. Spermidine-triggered autophagy ameliorates memory during aging. Autophagy 2014, 10, 178–179. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, S.; Zhang, Y.; Lin, X.; Song, Y.; Xue, Z.; Qian, H.; Wang, S.; Wan, G.; Zheng, X.; et al. Induction of autophagy by spermidine is neuroprotective via inhibition of caspase 3-mediated Beclin 1 cleavage. Cell Death Dis. 2017, 8, e2738. [Google Scholar] [CrossRef] [PubMed]
- Luan, Y.; Ren, X.; Zheng, W.; Zeng, Z.; Guo, Y.; Hou, Z.; Guo, W.; Chen, X.; Li, F.; Chen, J.F. Chronic Caffeine Treatment Protects Against α-Synucleinopathy by Reestablishing Autophagy Activity in the Mouse Striatum. Front. Neurosci. 2018, 12, 301. [Google Scholar] [CrossRef]
- Hung, J.Y.; Hsu, Y.L.; Li, C.; Ko, Y.C.; Ni, W.C.; Huang, M.S.; Kuo, P.L. 6-Shogaol, an active constituent of dietary ginger, induces autophagy by inhibiting the AKT/mTOR pathway in human non-small cell lung cancer A549 cells. J. Agric. Food Chem. 2009, 57, 9809–9816. [Google Scholar] [CrossRef] [PubMed]
- Li, T.Y.; Chiang, B.H. 6-shogaol induces autophagic cell death then triggered apoptosis in colorectal adenocarcinoma HT-29 cells. Biomed. Pharmacother. 2017, 93, 208–217. [Google Scholar] [CrossRef]
- Xiao, K.; Jiang, J.; Guan, C.; Dong, C.; Wang, G.; Bai, L.; Sun, J.; Hu, C.; Bai, C. Curcumin induces autophagy via activating the AMPK signaling pathway in lung adenocarcinoma cells. J. Pharmacol. Sci. 2013, 123, 102–109. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Li, X.H.; Zeng, X.C.; Li, J.; Zhou, J.; Xiao, B.; Hu, K. Curcumin protects neuronal cells against status-epilepticus-induced hippocampal damage through induction of autophagy and inhibition of necroptosis. Can. J. Physiol. Pharmacol. 2017, 95, 501–509. [Google Scholar] [CrossRef]
- Brimson, J.M.; Prasanth, M.I.; Malar, D.S.; Thitilertdecha, P.; Kabra, A.; Tencomnao, T.; Prasansuklab, A. Plant Polyphenols for Aging Health: Implication from Their Autophagy Modulating Properties in Age-Associated Diseases. Pharmaceuticals 2021, 14, 982. [Google Scholar] [CrossRef] [PubMed]
- Stavoe, A.K.H.; Holzbaur, E.L.F. Autophagy in Neurons. Annu. Rev. Cell Dev. Biol. 2019, 35, 477–500. [Google Scholar] [CrossRef]
- Ji, S.; Xiong, M.; Chen, H.; Liu, Y.; Zhou, L.; Hong, Y.; Wang, M.; Wang, C.; Fu, X.; Sun, X. Cellular rejuvenation: Molecular mechanisms and potential therapeutic interventions for diseases. Signal Transduct Target Ther. 2023, 8, 116. [Google Scholar] [CrossRef]
- Martinelli, S.; Anderzhanova, E.A.; Bajaj, T.; Wiechmann, S.; Dethloff, F.; Weckmann, K.; Heinz, D.; Ebert, T.; Hartmann, J.; Geiger, T.M.; et al. Stress-primed secretory autophagy promotes extracellular BDNF maturation by enhancing MMP9 secretion. Nat. Commun. 2021, 12, 4643. [Google Scholar] [CrossRef]
- Peker, N.; Gozuacik, D. Autophagy as a Cellular Stress Response Mechanism in the Nervous System. J. Mol. Biol. 2020, 432, 2560–2588. [Google Scholar] [CrossRef]
- Cappucci, U.; Noro, F.; Casale, A.; Pimpinell, S. The Hsp70 chaperone is a major player in stress-induced transposable element activation. Proc. Natl. Acad. Sci. USA 2019, 116, 17943–17950. [Google Scholar] [CrossRef] [PubMed]
- Leon, M.; Woo, C. Environmental Enrichment and Successful Aging. Front. Behav. Neurosci. 2018, 12, 155. [Google Scholar] [CrossRef]
- Colavitta, M.F.; Grasso, L.; Barrantes, F.J. Environmental Enrichment in Murine Models and Its Translation to Human Factors Improving Conditions in Alzheimer Disease. J. Prev. Alzheimers Dis. 2023, 10, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Birch, A.M.; Kelly, Á.M. Lifelong environmental enrichment in the absence of exercise protects the brain from age-related cognitive decline. Neuropharmacology 2019, 145, 59–74. [Google Scholar] [CrossRef] [PubMed]
- Balietti, M.; Conti, F. Environmental enrichment and the aging brain: Is it time for standardization? Neurosci. Biobehav. Rev. 2022, 139, 104728. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Haase, M.; Best, L.; Groth, M.; Lindner, J.; Witte, O.W.; Kaleta, C.; Frahm, C. Restoring Age-Related Cognitive Decline through Environmental Enrichment: A Transcriptomic Approach. Cells 2022, 11, 3864. [Google Scholar] [CrossRef]
- Barone, I.; Novelli, E.; Strettoi, E. Long-term preservation of cone photoreceptors and visual acuity in rd10 mutant mice exposed to continuous environmental enrichment. Mol. Vis. 2014, 20, 1545–1556. [Google Scholar]
- Levine, J.N.; Chen, H.; Gu, Y.; Cang, J. Environmental Enrichment Rescues Binocular Matching of Orientation Preference in the Mouse Visual Cortex. J. Neurosci. 2017, 37, 5822–5833. [Google Scholar] [CrossRef]
- Gurfein, B.T.; Davidenko, O.; Premenko-Lanier, M.; Milush, J.M.; Acree, M.; Dallman, M.F.; Touma, C.; Palme, R.; York, V.A.; Fromentin, G.; et al. Environmental enrichment alters splenic immune cell and enhances secondary influenza vaccine responses in mice. Mol. Med. 2014, 20, 179–190. [Google Scholar] [CrossRef]
- Vitalo, A.G.; Gorantla, S.; Fricchione, J.G.; Scichilone, J.M.; Camacho, J.; Niemi, S.M.; Denninger, J.W.; Benson, H.; Yarmush, M.L.; Levine, J.B. Environmental enrichment with nesting material accelerates wound healing in isolation-reared rats. Behav. Brain Res. 2012, 226, 606–612. [Google Scholar] [CrossRef]
- Bice, B.D.; Stephens, M.R.; Georges, S.J.; Venancio, A.R.; Bermant, P.C.; Warncke, A.V.; Affolter, K.E.; Hidalgo, J.R.; Angus-Hill, M.L. Environmental Enrichment Induces Pericyte and IgA-Dependent Wound Repair and Lifespan Extension in a Colon Tumor Model. Cell Rep. 2017, 19, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, D.; Aranda, M.L.; González Fleitas, M.F.; Chianelli, M.S.; Fernandez, D.C.; Sande, P.; Rosenstein, R.E. Environmental enrichment protects the retina from early diabetic damage in adult rats. PLoS ONE 2014, 9, e101829. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.C.; Mahato, N.K.; Nakazawa, M.; Law, T.D.; Thomas, J.S. The power of the mind: The cortex as a critical determinant of muscle strength/weakness. J. Neurophysiol. 2014, 112, 3219–3226. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, K.; Shen, X. Environmental enrichment attenuated sevoflurane-induced neurotoxicity through the PPAR-γ signaling pathway. Biomed. Res. Int. 2015, 2015, 107149. [Google Scholar] [CrossRef] [PubMed]
- Scarola, S.; Bardi, M. Environmental enrichment modulates inflammation during development in long-evans rats (Rattus norvegicus). Dev. Psychobiol. 2021, 63, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Matur, E.; Akyaz, İ.; Eraslan, E.; Ergul Ekiz, E.; Eseceli, H.; Keten, M.; Metiner, K.; Aktaran Bala, D. The effects of environmental enrichment and transport stress on the weights of lymphoid organs, cell-mediated immune response, heterophil functions and antibody production in laying hens. Anim. Sci. J. 2016, 87, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Wolinsky, F.; Unverzagt, F.W.; Smith, D.M.; Jones, R.; Wright, E.; Tennstedt, S.L. The effects of the ACTIVE cognitive training trial on clinically relevant declines in health-related quality of life. J. Gerontol. B 2006, 61, S281–S287. [Google Scholar] [CrossRef]
- Rebok, G.W.; Ball, K.; Guey, L.T.; Jones, R.N.; Kim, H.Y.; King, J.W.; Marsiske, M.; Morris, J.N.; Tennstedt, S.L.; Unverzagt, F.W.; et al. ACTIVE Study Group. Ten-year effects of the advanced cognitive training for independent and vital elderly cognitive training trial on cognition and everyday functioning in older adults. J. Am. Geriatr. Soc. 2014, 62, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Joëls, M.; Karst, H.; Sarabdjitsingh, R.A. The stressed brain of humans and rodents. Acta Physiol. 2018, 223, e13066. [Google Scholar] [CrossRef]
- Korneeva, N.L. Integrated Stress Response in Neuronal Pathology and in Health. Biochemistry 2022, 87 (Suppl. S1), S111–S127. [Google Scholar] [CrossRef]
- Farley, M.M.; Watkins, T.A. Intrinsic Neuronal Stress Response Pathways in Injury and Disease. Annu. Rev. Pathol. 2018, 13, 93–116. [Google Scholar] [CrossRef]
- Kim, K.W.; Jin, Y. Neuronal responses to stress and injury in C. elegans. FEBS Lett. 2015, 589, 1644–1652. [Google Scholar] [CrossRef]
- Schulz, A.; Sekine, Y.; Oyeyemi, M.J.; Abrams, A.J.; Basavaraju, M.; Han, S.M.; Groth, M.; Morrison, H.; Strittmatter, S.M.; Hammarlund, M. The stress-responsive gene GDPGP1/mcp-1 regulates neuronal glycogen metabolism and survival. J. Cell Biol. 2020, 219, e201807127. [Google Scholar] [CrossRef] [PubMed]
- Freeland, K.; Boxer, L.M.; Latchman, D.S. The cyclic AMP response element in the Bcl-2 promoter confers inducibility by hypoxia in neuronal cells. Brain Res. Mol. Brain Res. 2001, 92, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Korte, M. The impact of the digital revolution on human brain and behavior: Where do we stand? Dialogues Clin. Neurosci. 2020, 22, 101–111. [Google Scholar] [CrossRef]
- Loh, K.K.; Kanai, R. How Has the Internet Reshaped Human Cognition? Neuroscientist 2016, 22, 506–520. [Google Scholar] [CrossRef]
- Menghini, F.; van Rijsbergen, N.; Treves, A. Modelling adaptation aftereffects in associative memory. Neurocomputing 2007, 70, 2000–2004. [Google Scholar] [CrossRef]
- Small, G.W.; Lee, J.; Kaufman, A.; Jalil, J.; Siddarth, P.; Gaddipati, H.; Moody, T.D.; Bookheimer, S.Y. Brain health consequences of digital technology use. Dialogues Clin. Neurosci. 2020, 22, 179–187. [Google Scholar] [CrossRef]
- Advanced Cognitive Training in Vital Elderly—ACTIVE—Study. IU School of Medicine. 16 November 2017. Available online: https://medicine.iu.edu/news/2017/11/brain-exercise-dementia-prevention (accessed on 2 June 2023).
- Abd-Alrazaq, A.; Abuelezz, I.; AlSaad, R.; Al-Jafar, E.; Ahmed, A.; Aziz, S.; Nashwan, A.; Sheikh, J. Serious Games for Learning Among Older Adults with Cognitive Impairment: Systematic Review and Meta-analysis. J. Med. Internet Res. 2023, 25, e43607. [Google Scholar] [CrossRef]
- Abd-Alrazaq, A.; Alhuwail, D.; Ahmed, A.; Househ, M. Effectiveness of Serious Games for Improving Executive Functions Among Older Adults with Cognitive Impairment: Systematic Review and Meta-analysis. JMIR Serious Games 2022, 10, e36123. [Google Scholar] [CrossRef]
- Yang, C.; Han, X.; Jin, M.; Xu, J.; Wang, Y.; Zhang, Y.; Xu, C.; Zhang, Y.; Jin, E.; Piao, C. The Effect of Video Game-Based Interventions on Performance and Cognitive Function in Older Adults: Bayesian Network Meta-analysis. JMIR Serious Games 2021, 9, e27058. [Google Scholar] [CrossRef] [PubMed]
- Clemenson, G.D.; Stark, S.M.; Rutledge, S.M.; Stark, C.E.L. Enriching hippocampal memory function in older adults through video games. Behav. Brain Res. 2020, 390, 112667. [Google Scholar] [CrossRef]
- Ramnath, U.; Rauch, L.; Lambert, E.V.; Kolbe-Alexander, T. Efficacy of interactive video gaming in older adults with memory complaints: A cluster-randomized exercise intervention. PLoS ONE 2021, 16, e0252016. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Jolly, C.; Bublys, K.; Immler, S. Trade-off between somatic and germline repair in a vertebrate supports the expensive germ line hypothesis. Proc. Natl. Acad. Sci. USA 2020, 117, 8973–8979. [Google Scholar] [CrossRef] [PubMed]
- Ermolaeva, M.A.; Segref, A.; Dakhovnik, A. DNA damage in germ cells induces an innate immune response that triggers systemic stress resistance. Nature 2013, 501, 416–420. [Google Scholar] [CrossRef]
- Shemesh, N.; Shai, N.; Ben-Zvi, A. Germline stem cell arrest inhibits the collapse of somatic proteostasis early in Caenorhabditis elegans adulthood. Aging Cell 2013, 12, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Ermolaeva, M.; Schumacher, B. The innate immune system as mediator of systemic DNA damage responses. Commun. Integr. Biol. 2013, 6, e26926. [Google Scholar] [CrossRef]
- Khodakarami, A.; Saez, I.; Mels, J.; Vilchez, D. Mediation of organismal aging and somatic proteostasis by the germline. Front. Mol. Biosci. 2015, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, P.; Metcalfe, N.B. The deteriorating soma and the indispensable germline: Gamete senescence and offspring fitness. Proc. Biol. Sci. 2019, 286, 20192187. [Google Scholar] [CrossRef]
- Avise, J.C. The evolutionary biology of aging, sexual reproduction and DNA repair. Evolution 1993, 47, 1293–1301. [Google Scholar] [CrossRef]
- Gracida, X.; Eckmann, C.R. Fertility and germline stem cell maintenance under different diets requires nhr-114/HNF4 in C. elegans. Curr. Biol. 2013, 23, 607–613. [Google Scholar] [CrossRef]
- Heininger, K. Aging is a deprivation syndrome driven by a germ-soma conflict. Ageing Res. Rev. 2002, 1, 481–536. [Google Scholar] [CrossRef]
- Douglas, P.M.; Dillin, A. The disposable soma theory of aging in reverse. Cell Res. 2014, 24, 7–8. [Google Scholar] [CrossRef]
- Qian, Y.; Ng, C.L.; Schulz, C. CSN maintains the germline cellular microenvironment and controls the level of stem cell genes via distinct CRLs in testes of Drosophila melanogaster. Dev. Biol. 2015, 398, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U. Paternal contributions to offspring health: Role of sperm small RNAs in intergenerational transmission of epigenetic information. Front. Cell Dev. Biol. 2019, 7, 215. [Google Scholar] [CrossRef]
- Eaton, S.A.; Jayasooriah, N.; Buckland, M.E.; Martin, D.I.K.; Cropley, J.E.; Suter, C.M. Roll over Weismann: Extracellular vesicles in the transgenerational transmission of environmental effects. Epigenomics 2015, 7, 1165–1171. [Google Scholar] [CrossRef]
- Marchal, I.; Tursun, B. Induced Neurons from Germ Cells in Caenorhabditis elegans. Front. Neurosci. 2021, 15, 771687. [Google Scholar] [CrossRef] [PubMed]
- Glaser, T.; Opitz, T.; Kischlat, T. Adult germ line stem cells as a source of functional neurons and glia. Stem Cells 2008, 26, 2434–2443. [Google Scholar] [CrossRef]
- Kim, B.J.; Lee, Y.A.; Kim, K.J. Effects of paracrine factors on CD24 expression and neural differentiation of male germline stem cells. Int. J. Mol. Med. 2015, 36, 255–262. [Google Scholar] [CrossRef]
- Streckfuss-Bömeke, K.; Vlasov, A.; Hülsmann, S. Generation of functional neurons and glia from multipotent adult mouse germ-line stem cells. Stem Cell Res. 2009, 2, 139–154. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Y.; Hai, Y. Efficient Conversion of Spermatogonial Stem Cells to Phenotypic and Functional Dopaminergic Neurons via the PI3K/Akt and P21/Smurf2/Nolz1 Pathway. Mol. Neurobiol. 2015, 52, 1654–1669. [Google Scholar] [CrossRef]
- Wang, X.; Chen, T.; Zhang, Y.; Li, B.; Xu, Q.; Song, C. Isolation and Culture of Pig Spermatogonial Stem Cells and Their in Vitro Differentiation into Neuron-Like Cells and Adipocytes. Int. J. Mol. Sci. 2015, 16, 26333–26346. [Google Scholar] [CrossRef]
- Teichert, A.M.; Pereira, S.; Coles, B. The neural stem cell lineage reveals novel relationships among spermatogonial germ stem cells and other pluripotent stem cells. Stem Cells Dev. 2014, 23, 767–778. [Google Scholar] [CrossRef]
- Levi-Ferber, M.; Salzberg, Y.; Safra, M.; Haviv-Chesner, A.; Bülow, H.E.; Henis-Korenblit, S. It’s all in your mind: Determining germ cell fate by neuronal IRE-1 in C. elegans. PLoS Genet. 2014, 10, e1004747. [Google Scholar] [CrossRef]
- Bonefas, K.M.; Iwase, S. Soma-to-germline transformation in chromatin-linked neurodevelopmental disorders? FEBS J. 2022, 289, 2301–2317. [Google Scholar] [CrossRef]
- Calogero, A.E.; Burrello, N.; Barone, N.; Palermo, I.; Grasso, U.; D’Agata, R. Effects of progesterone on sperm function: Mechanisms of action. Hum. Reprod. 2000, 15 (Suppl. S1), 28–45. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Min, K.J.; D’Alterio, C.; Villa-Cuesta, E.; Cumbers, J.; Lehmann, R.; Jones, D.L.; Tatar, M. Drosophila germ-line modulation of insulin signaling and lifespan. Proc. Natl. Acad. Sci. USA 2008, 105, 6368–6373. [Google Scholar] [CrossRef] [PubMed]
- Kimura, R.; Inoue, Y.U.; Kikkawa, T.; Tatehana, M.; Morimoto, Y.; Inada, H.; Oki, S.; Inoue, T.; Osumi, N. Detection of REST expression in the testis using epitope-tag knock-in mice generated by genome editing. Dev. Dyn. 2022, 251, 525–535. [Google Scholar] [CrossRef]
- Cheng, Y.; Yin, Y.; Zhang, A.; Bernstein, A.M.; Kawaguchi, R.; Gao, K.; Potter, K.; Gilbert, H.Y.; Ao, Y.; Ou, J.; et al. Transcription factor network analysis identifies REST/NRSF as an intrinsic regulator of CNS regeneration in mice. Nat. Commun. 2022, 13, 4418. [Google Scholar] [CrossRef]
- Zullo, J.M.; Drake, D.; Aron, L.; O’Hern, P.; Dhamne, S.C.; Davidsohn, N.; Mao, C.A.; Klein, W.H.; Rotenberg, A.; Bennett, D.A.; et al. Regulation of lifespan by neural excitation and REST. Nature 2019, 574, 359–364. [Google Scholar] [CrossRef]
- Calculli, G.; Lee, H.J.; Shen, K.; Pham, U.; Herholz, M.; Trifunovic, A.; Dillin, A.; Vilchez, D. Systemic regulation of mitochondria by germline proteostasis prevents protein aggregation in the soma of C. elegans. Sci. Adv. 2021, 7, eabg3012. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.H.; Yan, J.L.; Wang, Q.J.; Chen, H.C.; Ma, X.Z.; Yin, J.; Gao, L.P. Spermidine ameliorates the neuronal aging by improving the mitochondrial function in vitro. Exp. Gerontol. 2018, 108, 77–86. [Google Scholar] [CrossRef]
- Ghosh, I.; Sankhe, R.; Mudgal, J.; Arora, D.; Nampoothiri, M. Spermidine, an autophagy inducer, as a therapeutic strategy in neurological disorders. Neuropeptides 2020, 83, 102083. [Google Scholar] [CrossRef]
- Yang, N.; Liu, X.; Niu, X.; Wang, X.; Jiang, R.; Yuan, N.; Wang, J.; Zhang, C.; Lim, K.L.; Lu, L. Activation of Autophagy Ameliorates Age-Related Neurogenesis Decline and Neurodysfunction in Adult Mice. Stem Cell Rev. Rep. 2022, 18, 626–641. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Tian, G.G.; Pei, X.; Hu, X.; Wu, J. Spermidine induces cytoprotective autophagy of female germline stem cells in vitro and ameliorates aging caused by oxidative stress through upregulated sequestosome-1/p62 expression. Cell Biosci. 2021, 11, 107. [Google Scholar] [CrossRef]
- Bhukel, A.; Madeo, F.; Sigrist, S. Spermidine boosts autophagy to protect against synapse ageing. Autophagy 2017, 13, 444–445. [Google Scholar] [CrossRef]
- Sharma, S.; Kumar, P.; Deshmukh, R. Neuroprotective potential of spermidine against rotenone induced Parkinson’s disease in rats. Neurochem. Int. 2018, 116, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, S.; Hofer, S.J.; Zimmermann, A.; Pechlaner, R.; Dammbrueck, C.; Pendl, T.; Marcello, G.M.; Pogatschnigg, V.; Bergmann, M.; Müller, M.; et al. Dietary spermidine improves cognitive function. Cell Rep. 2021, 35, 108985. [Google Scholar] [CrossRef]
- Babajanyan, S.G.; Koonin, E.V.; Allahverdyan, A.E. Thermodynamic selection: Mechanisms and scenarios. New J. Phys. 2022, 24, 053006. [Google Scholar] [CrossRef] [PubMed]
- Teulière, J.; Bhattacharya, D.; Bapteste, E. Ancestral germen/soma distinction in microbes: Expanding the disposable soma theory of aging to all unicellular lineages. Ageing Res. Rev. 2020, 60, 101064. [Google Scholar] [CrossRef]
- Lorenzini, A.; Stamato, T.; Sell, C. The disposable soma theory revisited: Time as a resource in the theories of aging. Cell Cycle 2011, 10, 3853–3856. [Google Scholar] [CrossRef][Green Version]
- Drenos, F.; Kirkwood, T.B. Modelling the disposable soma theory of ageing. Mech. Ageing Dev. 2005, 126, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, M. Neurons vs. Germline: A War of Hormetic Tradeoffs. Curr. Aging Sci. 2017, 10, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Kyriazis, M. Reversal of informational entropy and the acquisition of germ-like immortality by somatic cells. Curr. Aging Sci. 2014, 7, 9–16. [Google Scholar] [CrossRef]
- Aiello, L.C.; Wheeler, P. The expensive-tissue hypothesis. Curr. Anthropol. 1995, 36, 199–221. [Google Scholar] [CrossRef]
- Roff, D.A.; Mostowy, S.; Fairbairn, D.J. The evolution of trade-offs: Testing predictions on response to selection and environmental variation. Evolution 2002, c56, 84–95. [Google Scholar]
- Lesch, R.; Kotrschal, K.; Kitchener, A.C.; Fitch, W.T.; Kotrschal, A. The expensive-tissue hypothesis may help explain brain-size reduction during domestication. Commun. Integr. Biol. 2022, 15, 190–192. [Google Scholar] [CrossRef]
- Huang, C.H.; Yu, X.; Liao, W.B. The Expensive-Tissue Hypothesis in Vertebrates: Gut Microbiota Effect, a Review. Int. J. Mol. Sci. 2018, 19, 1792. [Google Scholar] [CrossRef]
- Nengovhela, A.; Ivy, C.M.; Scott, G.; Denys, C.; Taylor, P.J. Counter-gradient variation and the expensive tissue hypothesis explain parallel brain size reductions at high elevation in cricetid and murid rodents. Sci. Rep. 2023, 13, 5617. [Google Scholar] [CrossRef]
- Maklakov, A.A.; Immler, S. The Expensive Germline and the Evolution of Ageing. Curr. Biol. 2016, 26, R577–R586. [Google Scholar] [CrossRef]
- Teplyuk, N.M. Near-to-perfect homeostasis: Examples of universal aging rule which germline evades. J. Cell Biochem. 2012, 113, 388–396. [Google Scholar] [CrossRef]
- Kyriazis, M. The Indispensable Soma Hypothesis. Available online: www.indispensablesoma.info (accessed on 5 August 2023).
- Calabrese, E.J.; Calabrese, V. Enhancing health span: Muscle stem cells and hormesis. Biogerontology 2022, 23, 151–167. [Google Scholar] [CrossRef]
- Calabrese, E.J. Hormesis Mediates Acquired Resilience: Using Plant-Derived Chemicals to Enhance Health. Annu. Rev. Food Sci. Technol. 2021, 12, 355–381. [Google Scholar] [CrossRef]
- Patrick, R.P.; Johnson, T.L. Sauna use as a lifestyle practice to extend healthspan. Exp. Gerontol. 2021, 154, 111509. [Google Scholar] [CrossRef]
- Ji, L.L.; Kang, C.; Zhang, Y. Exercise-induced hormesis and skeletal muscle health. Free. Radic. Biol. Med. 2016, 98, 113–122. [Google Scholar] [CrossRef]
- Cook, R.; Calabrese, E.J. The importance of hormesis to public health. Cien. Saude. Colet. 2007, 12, 955–963. [Google Scholar] [CrossRef][Green Version]
- Mattson, M.P. Dietary factors, hormesis and health. Ageing Res. Rev. 2008, 7, 43–48. [Google Scholar] [CrossRef]
- Bukowski, J.A.; Lewis, R.J. Hormesis and health: A little of what you fancy may be good for you. South Med. J. 2000, 93, 371–374. [Google Scholar] [CrossRef]
- Poumadère, M. Hormesis: Public health policy, organizational safety and risk communication. Hum. Exp. Toxicol. 2003, 22, 39–41; discussion 43–49. [Google Scholar] [CrossRef]
- Rattan, S.I. Ageing, gerontogenes, and hormesis. Indian J. Exp. Biol. 2000, 38, 1–5. [Google Scholar]
- Forcina, L.; Franceschi, C.; Musarò, A. The hormetic and hermetic role of IL-6. Ageing Res. Rev. 2022, 80, 101697. [Google Scholar] [CrossRef]
- Lajqi, T.; Stojiljkovic, M.; Wetzker, R. Toxin-induced hormesis may restrain aging. Biogerontology 2019, 20, 571–581. [Google Scholar] [CrossRef]
- Martel, J.; Chang, S.H.; Wu, C.; Peng, H.H.; Hwang, T.L.; Ko, Y.F.; Young, J.D.; Ojcius, D.M. Recent advances in the field of caloric restriction mimetics and anti-aging molecules. Ageing Res. Rev. 2021, 66, 101240. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Iavicoli, I.; Calabrese, V. Hormesis: Why it is important to biogerontologists. Biogerontology 2012, 13, 215–235. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S. Possible role of NF-κB in hormesis during ageing. Biogerontology 2012, 13, 637–646. [Google Scholar] [CrossRef]
- Jacome Burbano, M.S.; Gilson, E. The Power of Stress: The Telo-Hormesis Hypothesis. Cells 2021, 10, 1156. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Hormetin Supplements | Description | Reference |
|---|---|---|
| Rhodiola | Adaptogen, antioxidant | [37,38] |
| Schisandra | Adaptogen, liver conditions, tonic | [39,40,41,42] |
| Spermidine | Biological modulator, longevity (See section on spermidine) | [43,44,45] |
| Caffeine | Cognitive enhancer | [46] |
| Ginger | General health | [47,48] |
| Turmeric (Curcumin) | Anti-inflammation, antioxidant | [49,50,51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriazis, M.; Swas, L.; Orlova, T. The Impact of Hormesis, Neuronal Stress Response, and Reproduction, upon Clinical Aging: A Narrative Review. J. Clin. Med. 2023, 12, 5433. https://doi.org/10.3390/jcm12165433
Kyriazis M, Swas L, Orlova T. The Impact of Hormesis, Neuronal Stress Response, and Reproduction, upon Clinical Aging: A Narrative Review. Journal of Clinical Medicine. 2023; 12(16):5433. https://doi.org/10.3390/jcm12165433
Chicago/Turabian StyleKyriazis, Marios, Lama Swas, and Tetiana Orlova. 2023. "The Impact of Hormesis, Neuronal Stress Response, and Reproduction, upon Clinical Aging: A Narrative Review" Journal of Clinical Medicine 12, no. 16: 5433. https://doi.org/10.3390/jcm12165433
APA StyleKyriazis, M., Swas, L., & Orlova, T. (2023). The Impact of Hormesis, Neuronal Stress Response, and Reproduction, upon Clinical Aging: A Narrative Review. Journal of Clinical Medicine, 12(16), 5433. https://doi.org/10.3390/jcm12165433