
Exercise and Weight Management: The Role of Leptin—A Systematic Review and Update of Clinical Data from 2000–2022



Abstract
1. Introduction
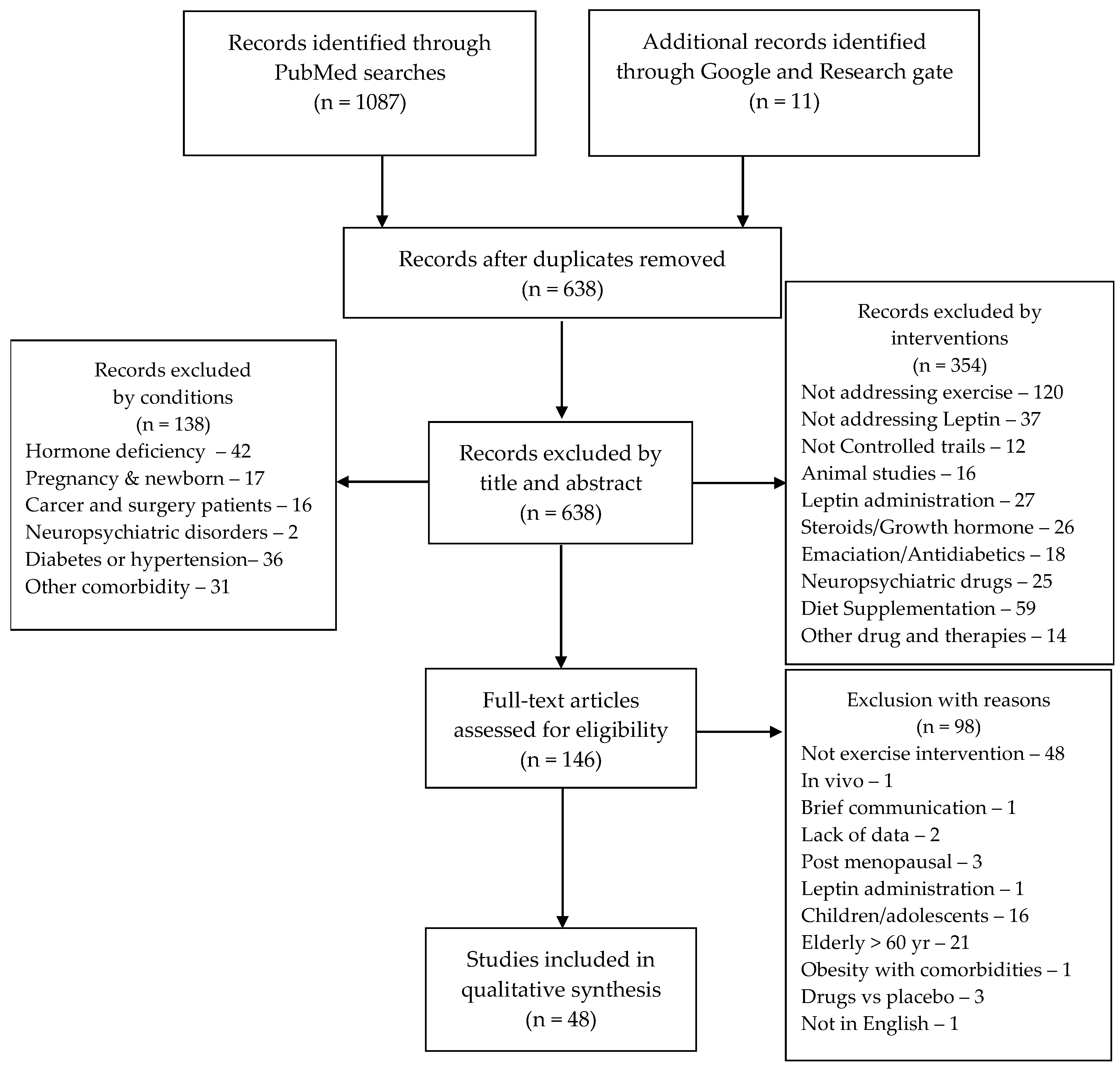
2. Materials and Methods
3. Results

{kind=link}
| Author and DOI | Subjects and Design | Main Outcomes |
|---|---|---|
| Hilton & Loucks [31]. DOI 10.1152/ajpendo.2000.278.1.e43 | Young women (age range, 20–22 yr, BMI, 24–26 kg/m2) engaged in either an exercise or diet intervention, with controlled energy intake, were evaluated for diurnal leptin levels during the estrous cycle. | Low energy availability suppressed the 24 h mean and amplitude. The negative effects of low energy intake on the leptin rhythm were more pronounced in sedentary women. |
| Zaccaria, M. et al. [32]. DOI 10.1007/s00421-002-0606-4 | Active men (age range, 32–49 yr, BMI, 21–27 kg/m2) engaged in either a half-marathon run (21.097 km—1400 kcal), a ski-alpinism race (about 45 km—5000 kcal), and an ultramarathon race (100 km—7000 kcal) were evaluated for energy expenditure and leptin levels. | Serum leptin decreased 3% in the ultramarathoners and 7% in the ski-alpinism athletes but did not change after the half-marathon. Variation in free fatty acids was inversely correlated with variation in leptin. |
| Tsofliou, F. et al. [33]. DOI 10.1038/sj.ijo.0802406 | Adult women (age range 42–58 yr, BMI, 30–37 kg/m2) were tested for blood leptin before and after a 20 min brisk walk, a 58.5 g chocolate-based snack, and control (sitting, TV-watching) conditions. | There was no effect of any intervention or effect over time on serum leptin concentrations. No associations were found between serum leptin and blood glucose or plasma free fatty acids. |
| Kraemer, R. R. et al. [34]. DOI 10.1385/ENDO:21:3:261 | Active men (age range, 25–32 yr, BMI, 10–12 kg/m2) completed an intermittent treadmill workout at the progressive intensity of 60% for 10 min, 75% for 10 min, 90% for 5 min, and 100% VO2max for 2 min; with samples collected 40 and 10 min prior, and immediately after exercise. | Exercise increased leptin concentrations after 5 min at 90% of VO2max, then declined to resting values during recovery. |
| Tsai, A. C. et al. [35]. DOI 10.1016/S0955-2863(03)00105-0 | Adult women (age range 22–55 yr, BMI, 17.4–40.3 kg/m2) underwent a 9-day energy deficit period with either pre-planned diet or activity schedule and a food-repletion period. Those on dieting would consume 75% of baseline energy intake and maintained the level of activity. Those in exercise achieved the same energy deficit by performing extra daily physical activity while maintaining the baseline energy intake. Samples were collected from baseline and after treatments. | Blood leptin concentration similarly reduced after exercise and food restriction. |
| Desgorces, F. D. et al. [36]. DOI 10.1007/s00421-003-1030-0 | Male rowers (age range, 20–23 yr) were evaluated for leptin levels before, 120 min after and 24 h after a 90 min rowing session at 70–75% of VO2max throughout the season. | Resting leptin levels were lower after the program. After the program, the past-task leptin levels were reduced after 120 min of recovery and returned to baseline levels after 24 h of recovery. |
| Rubin, M. R. et al. [37]. DOI 10.1249/01.MSS.0000155402.93987.C0 | Young men (age range, 20–26 yr, BMI, 25–31 kg/m2) underwent a resistance training session with the first of six sets at 10 RM (80–85% of the 1 RM) with a 2 min rest between sets, and weight was adjusted to allow completion of that set and subsequent sets. | There was no effect of exercise on plasma leptin concentrations. |
| Kondo, T. et al. [38]. DOI 10.1507/endocrj.53.189 | Young women (age range, 18–23 yr; BMI > 25 kg/m2) undergoing a 7-month, moderate aerobic exercise program were tested for leptin, adipokine, among other metabolites. | Leptin levels were higher and adipokine levels were lower in women with obesity compared to controls. Adiponectin levels were negatively correlated with BMI and leptin. |
| Ara, I. et al. [39]. DOI 10.1017/BJN20061956 | Healthy men (age range, 21–25 yr; mean BMI, 24 kg/m2) were examined for serum free leptin index in a 6-week resistance training program. | Resistance training did not alter basal leptin levels. Changes in leptin were not related to leptin levels, leptin receptor, or free testosterone. |
| Moro, C. et al. [40]. DOI 10.1038/oby.2007.267 | Sedentary subjects (age range, 26.2–28.4 yr; BMI, 27–28 kg/m2) performed two exercise sessions: 1, 30 min cycling at 30% of VO2max followed by 30 min at 50%; and 2, 30 min at 30% followed by 30 min at 70% of VO2max. | Plasma leptin concentrations were higher in women than in men, and exercise did not induce any changes in plasma levels of leptin. |
| Kyriazis, G. A. et al. [41]. DOI 10.1097/JSM.0b013e31802e9c38 | Men with obesity (age range, 22–26; BMI, >30 kg/m2) submitted to a 60 min treadmill test at 50–60% of VO2max and controls were evaluated for leptin levels. | No changes were observed for leptin, ghrelin or insulin levels during exercise or following recovery. |
| König, D. et al. [42]. DOI 10.1159/000119416 | Adult men and women (age range, 40–55 yr; BMI, 29–33 kg/m2) participated in a 6-week, increasing physical activity and either a meal-replacement or a low-calorie diet. | Leptin and insulin levels were reduced in both groups. |
| Volpe, S. L. et al. [43]. DOI 10.1080/07315724.2008.10719691 | Overweight women and men (age range, 37–52 yr; BMI, 27–33 kg/m2) were investigated for the effect of 6-month diet and/or exercise program on leptin levels. | Decreases in leptin levels were observed in the diet, and diet and exercise groups. In the exercise group, a decrease in leptin levels was observed only in males. |
| Trapp, E. G. et al. [44]. DOI 10.1038/sj.ijo.0803781 | Young women (age range 18–22 yr; BMI, 21–25 kg/m2) were evaluated for subcutaneous and metabolic parameters pre- post a 15-week, high-intensity intermittent cycling program. | There was a decrease in basal leptin levels after the program. |
| Liu, T. C. et al. [45]. DOI 10.1080/02640410801885925 | Elite kayak athletes (age range, 20–22 yr; BMI, 23–25 kg/m2) were examined for leptin levels in a partial detrained vs. total detrained condition. | Both partial and total detraining result in elevated basal leptin levels, while the totally detrained athletes had greater increases in leptin levels than the partially detrained athletes. |
| Jürimäe, J. et al. [46]. DOI 10.1249/MSS.0b013e31818313e6 | Male rowers (age range, 18–22 yr; body fat, 7–14%) were tested for their hormonal response to a 2 h rowing session and follow up. | Leptin levels decreased by 20% during the 30 min follow up. Basal leptin levels correlated with energy expenditure, energy intake and basal testosterone values. |
| Cheng, MHY et al. [47]. DOI 10.1016/j.appet.2008.09.015 | Active men (age range, 19–29; Body fat, 5–13%) were tested for their hormonal response to meal consumption followed by exercise, exercise followed by meal, or meal consumption only. | No effects were observed in plasma leptin levels. |
| Hagobian, T. A. et al. [48]. DOI 10.1152/ajpregu.90671.2008 | Young adults (age range, 16–38 yr; BMI, 22–32 kg/m2) were evaluated for sex differences in hormones and appetite in response to four exercise bouts with or without energy imbalance. | Lower leptin levels were associated with lower appetite following exercise with induced energy deficit. |
| Guadalupe-Grau, A. et al. [49]. DOI 10.1152/japplphysiol.91469.2008 | Young men and women (age range, 20–26 yr; BMI, 20–28 kg/m2) were examined for a sex dimorphism in serum osteocalcin and leptin responses to a 9-week resistance, plus plyometric, training program. | Serum leptin levels correlated with the percentage of muscle phenotype, IIx (myosin heavy chain composition), in men. Serum leptin concentration was reduced with training in women. Changes in leptin levels were associated with fat mass in men but not in women. |
| Bergouignan, A. et al. [50]. DOI 10.1210/jc.2009-1005 | Inactive women (age range, 30–35 yr; BMI, 18–22 kg/m2) were evaluated pre-post to the following conditions: a strict 60-day bed rest, or combined aerobic/resistive exercise training concomitantly to bed rest. | No changes in plasma leptin levels were found. |
| Sartor, F. et al. [51]. DOI 10.1007/s00421-010-1571-y | Healthy adults (age range, 27–47 yr; BMI, 28–36 kg/m2) were engaged to a 75% energy expenditure reduction in calorie intake or diet, plus a 2-week HIIT with dietary intake increased via the activity correction factor to compensate for energetic costs. | Plasma leptin was reduced in both groups. |
| Cooper, J. A. et al. [52]. DOI 10.1016/j.appet.2010.10.009 | Healthy men (age range, 18–45 yr; BMI, 18–30 kg/m2) were crossed over a 3-day, high-saturated fat or high-monounsaturated fat diet with a cycling session at 45% of VO2max to reach the 24 h EE of 1.8 × resting metabolic rate; and the corresponding sedentary conditions. | Average 24 h leptin levels were lower during exercise vs. sedentary conditions. |
| Kraemer, R. R. et al. [53]. DOI 10.1249/MSS.0b013e3182114ab9 | Healthy men (age range, 20–25 yr; BMI, 20–28 kg/m2) performed 90 min of treadmill exercise at 60% of VO2max with blood samples collected twice before, every 18 min during exercise, and every 20 min during 1 h of recovery. | Plasma leptin concentrations remained stable across the measurements. |
| Moran, C. N. et al. [54]. DOI 10.1016/j.metabol.2009.12.026 | Healthy women (age range 28–42 yr; BMI, 23–33 kg/m2) with and without a family history of diabetes accomplished a 7-week exercise session performed at 65–80% of maximum HR, with pre-post blood and abdominal subcutaneous adipose tissue samples analyzed. | Leptin mRNA decreased similarly in both groups. Plasma leptin decreased more in those with family history. The expression of the long leptin receptor mRNA increased only in those with family history. Changes in plasma leptin and leptin mRNA correlated with changes in insulin sensitivity. |
| Balaguera-Cortes, L. et al. [55]. DOI 10.1139/H11-121 | Healthy men (age range, 19–23 yr; BMI, 21–26 kg/m2) completed 3 trials as: 45 min of resistance exercise, running, or resting control, followed by a breakfast ad libitum. | There was no effect showing exercise effects on pre-prandial plasma concentrations of leptin. There was a time effect, however, with lower leptin concentrations observed post-meal compared with baseline. |
| Rosa, G. et al. [56]. DOI 10.14310/horm.2002.1311 | Adults (age range, 22–32 yr; BMI, 22–28 kg/m2) participated in three trail sessions: control, indoor cycling class followed by strength training, and resistance training with a 5-day rest in between, and blood samples were collected in each session. | There was a similar reduction in leptin levels after both exercise sessions. |
| Arikawa, A. Y. et al. [57]. DOI 10.1249/MSS.0b013e3182059eda | Inactive women (age range, 18–30 yr; BMI, 18–40 kg/m2) were evaluated for changes in inflammatory markers during a 16-week aerobic exercise program. | There was no effect of exercise on the levels of leptin or adiponectin. |
| Kelly, K. R. et al. [58]. DOI 10.1249/MSS.0b013e318228bf85 | Adults with obesity (age range, 52–58 yr; BMI, 32–35 kg/m2) were tested for adiponectin and leptin after a 7-day, high-intensity exercise program. | There were increases in adiponectin and the ratio of adiponectin to leptin, and a decrease in leptin after the program. |
| Numao, S. et al. [59]. DOI 10.1159/000346205 | Obese males (age range, 46–50 yr; BMI, 28–30 kg/m2) were tested for circulating leptin before and after a 12-week exercise training program. | There was a decrease in circulating leptin levels after training. |
| Zaccaria, M. et al. [60]. DOI 10.3275/8656 | Young men (age range, 22–32 yr; BMI, 20–25 kg/m2) were assessed for leptin levels during a 4 h treadmill running task. | Plasma leptin levels decreased within 2 h after the task. There were negative correlations between leptin, norepinephrine and energy expenditure. |
| Martins, C. et al. [61]. DOI 10.1249/MSS.0b013e31827d1618 | Sedentary adults (age range, 28–35 yr; BMI, 28–34 kg/m2) underwent a 12-week program: 5-day treadmill walking or running at 75% HRmax, for 500-kcal energy deficit per session, per week. | There was a reduction in the fasting and postprandial leptin concentrations after exercise intervention. |
| Morishima, T. et al. [62]. DOI 10.1111/cpf.12069 | Sedentary adults (age range, 28–35 yr; BMI, 24–27 kg/m2) accomplished 4-week training 3 days per week at 55% HRmax, under hypoxic and normoxic conditions. | A reduction of postprandial leptin response was observed in both groups. |
| Mendham, A. E. et al. [63]. DOI 10.1007/s00421-014-2953-3 | Sedentary men (age range, 42–54; BMI, 24–31 kg/m2) were tested before and after an 8-week cycling, small-sided game training of 3 days/week, and under control conditions. | Both exercise conditions decreased the concentration of plasma leptin. |
| Debevec, T. et al. [64]. DOI 10.1371/journal.pone.0098874 | Healthy men (age range, 21–28 yr; BMI, 19–25 kg/m2) underwent a 10-day hypoxic confinement at 4000 m with daily moderate intensity exercise or control conditions. | No differences between groups or testing periods were noticed for plasma leptin concentrations. |
| Ahmadizad, S. et al. [65]. DOI 10.1016/j.clinbiochem.2013.12.019 | Sedentary men (age range, 22–24 yr; BMI, 25–30 kg/m2) were allocated to undergo an 8-week resistance training with different protocols of periodization. | No changes were observed in plasma leptin concentrations. |
| Sim, A. Y. et al. [66]. DOI 10.1249/MSS.0000000000000687 | Inactive men (age range, 24–39 yr; BMI, 25–29 kg/m2) were randomized into either HIIT or moderate-intensity continuous exercise training at three sessions per week, during 12 weeks. | Leptin concentrations were lower after HIIT but not after moderate control conditions. |
| Kim, Y. S. et al. [67]. DOI 10.1111/cen.12601 | Young men (age range, 22–29 yr; BMI, 26–32 kg/m2) were examined for changes in adipocytokines following an 8-week treadmill running exercise at 65–75% VO2max (~600 Kcal), four times per week. | Leptin levels decreased after program. Adiponectin over total adiponectin ratio increased. |
| King, J. A. et al. [68]. DOI 10.1016/j.physbeh.2014.12.050 | Young men (age range, 20–24 yr; BMI, 20–25 kg/m2) were investigated for corrective changes in appetite-regulatory parameters on the day after a single bout of exercise. | Circulating leptin levels were reduced on the day after exercise. The AUC for plasma leptin showed reduced circulating levels the day after. |
| Douglas, J. A. et al. [69]. DOI 10.1016/j.appet.2015.05.006 | Young men (age range, 18–23 yr; BMI, 21–25 kg/m2) were examined for appetite, food intake and regulatory hormones in response to a single exercise bout. | Fasting leptin levels did not differ following the single exercise bout. |
| Racil, G. et al. [70]. DOI 10.1139/apnm-2015-0384 | Young women (age range, 15–18 yr; BMI, >25 kg/m2) were evaluated for biochemical parameters pre-post 2 HIIT programs (with and without plyometrics). | There was a decrease in leptin and an increase in adiponectin levels after both training programs. The plasma leptin/adiponectin ratio was reduced in both programs, but changes were greater in the plyometric + HIIT group. |
| Kong, Z. et al. [71]. DOI 10.1155/2016/4073618 | Young men and women (age ranger, 19–21 yr; BMI, 24–28 kg/m2) were analyzed for hormonal responses to a 5-week HIIT compared to moderate-intensity training. | No changes were observed in leptin levels among women. |
| Cameron, J. D. et al. [72]. DOI 10.3945/ajcn.115.115584 | Young men (age range, 18–28 yr; body weight, 71–95 kg) participated in 2 experimental conditions: 25% daily needs energy deficits induced by diet only and by exercise only, and tested before and after 3 and 4 days of the intervention. | There was no condition effect for plasma leptin concentrations. |
| Vardar, S. A. et al. [73]. DOI 10.1080/13813455.2017.1369998 | Young women (age range, 18–25 yr; BMI, 25–35 kg/m2) completed 19 days of high-intensity training through six sessions of 4–6 repeats of a Wingate test. Plasma adipokine levels were measured before exercise, and at 5 and 90 min after exercise, on the first and last training days. | Plasma leptin concentrations decreased 5 min after exercise and remained reduced following 90 min in both the first and last training days. |
| Caldeira, R. S. et al. [74]. DOI 10.1016/j.clnesp.2018.08.005 | Twenty men (are range, 18–35 yr; BMI, 18–29 kg/m2) performed 5 weeks of HIIT (5 km: 1 min running at 100% speed/1 min passive recovery), or running 5 km at 70% of the VO2max, 3 days a week. Leptin was analyzed at baseline, 24 h, and 48 h after an exercise session pre- and post-five weeks. | Plasma leptin concentrations were reduced in both exercise groups. LR increased post-five weeks of HIIT but continuous intensity exercise training. |
| Tremblay, A. et al. [75]. DOI 10.1139/apnm-2019-0019 | Older adult men and women (age range, 50–70 yr; BMI, 25–35 kg/m2) were tested for plasma leptin and ghrelin levels responses to a 12-month, high-resistance and low-aerobic exercise, low-resistance and high-aerobic exercise, and low-resistance and low-aerobic exercise. | Leptin levels decreased during the first 3 months and then plateaued. |
| Sommer, C. et al. [76]. DOI 10.1210/jc.2017-02126 | Adult men (age range, 40–65 yr; BMI, 21–27 kg/m2) were analyzed for circulating levels of leptin receptors (LR) and related genes, as well as mRNA pathways in adipose tissue and skeletal muscle pre-post a 12-week intensive exercise program. | Plasma LR levels were correlated with leptin, fasting GLU and insulin sensitivity. High adipocyte LR expression was associated with the upregulation of oxidative phosphorylation, adipogenesis, fatty acid metabolism and peroxisomes, and downregulation of inflammatory response. In muscle, high-LR expression was associated with the upregulation of the pathways of oxidative phosphorylation, and KRAS and mTORC1 signaling, and downregulation of an inflammatory response. |
| Middelbeek, R. J. W. et al. [77]. DOI 10.1038/s41387-020-00144-x | Sedentary men (age range, 40–55 yr; BMI, 18.5–30 kg/m2) performed 6 sessions of 6 × 30 s all-out cycle ergometer sprints with 4 min of recovery between sprints, or cycle ergometer exercise at 60% VO2max gradually increasing in duration from 40 to 60 min, over 2 weeks. | Both training programs reduced the concentrations of plasma leptin. |
| Zaman, G. S. et al. [78]. DOI 10.1155/2021/6684167 | Men and women (age range, 30–60 yr; any BMI) were checked for the impact of 12-week resistance exercise on biomedical profile at high altitude. | Leptin and Interleukin-6 decreased, while adiponectin increased. |
4. Discussion
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bull, F.C.; Al-Ansari, S.S.; Biddle, S.; Borodulin, K.; Buman, M.P.; Cardon, G.; Carty, C.; Chaput, J.P.; Chastin, S.; Chou, R.; et al. World Health Organization 2020 guidelines on physical activity and sedentary behaviour. Br. J. Sports Med. 2020, 54, 1451–1462. [Google Scholar] [CrossRef]
- Kercher, V.M.; Kercher, K.; Levy, P.; Bennion, T.; Alexander, C.; Amaral, P.C.; Batrakoulis, A.; Chávez, L.F.J.G.; Cortés-Almanzar, P.; Haro, J.L.; et al. 2023 Fitness Trends from Around the Globe. ACSM’s Health Fit. J. 2023, 27, 19–30. [Google Scholar] [CrossRef]
- Batrakoulis, A. Role of Mind—Body Fitness in Obesity. Diseases 2023, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Batrakoulis, A.; Jamurtas, A.Z.; Metsios, G.S.; Perivoliotis, K.; Liguori, G.; Feito, Y.; Riebe, D.; Thompson, W.R.; Angelopoulos, T.J.; Krustrup, P.; et al. Comparative Efficacy of 5 Exercise Types on Cardiometabolic Health in Overweight and Obese Adults: A Systematic Review and Network Meta-Analysis of 81 Randomized Controlled Trials. Circ. Cardiovasc. Qual. Outcomes 2022, 15, e008243. [Google Scholar] [CrossRef]
- Flores-Cordero, J.A.; Pérez-Pérez, A.; Jiménez-Cortegana, C.; Alba, G.; Flores-Barragán, A.; Sánchez-Margalet, V. Obesity as a Risk Factor for Dementia and Alzheimer’s Disease: The Role of Leptin. Int. J. Mol. Sci. 2022, 23, 5202. [Google Scholar] [CrossRef]
- Liu, L.; Shi, Z.; Ji, X.; Zhang, W.; Luan, J.; Zahr, T.; Qiang, L. Adipokines, adiposity, and atherosclerosis. Cell. Mol. Life Sci. 2022, 79, 272. [Google Scholar] [CrossRef] [PubMed]
- Considine, R.V.; Sinha, M.K.; Heiman, M.L.; Kriauciunas, A.; Stephens, T.W.; Nyce, M.R.; Ohannesian, J.P.; Marco, C.C.; McKee, L.J.; Bauer, T.L.; et al. Serum Immunoreactive-Leptin Concentrations in Normal-Weight and Obese Humans. Endocrinologist 1996, 6, 349. [Google Scholar] [CrossRef]
- Al-Sultan, A.; Al-Elq, A. Leptin levels in normal weight and obese Saudi adults. J. Fam. Community Med. 2006, 13, 97–102. [Google Scholar]
- Tentolouris, N.; Tsigos, C.; Perea, D.; Koukou, E.; Kyriaki, D.; Kitsou, E.; Daskas, S.; Daifotis, Z.; Makrilakis, K.; Raptis, S.A.; et al. Differential effects of high-fat and high-carbohydrate isoenergetic meals on cardiac autonomic nervous system activity in lean and obese women. Metabolism 2003, 52, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Poppitt, S.D.; Leahy, F.E.; Keogh, G.F.; Wang, Y.; Mulvey, T.B.; Stojkovic, M.; Chan, Y.K.; Choong, Y.S.; McArdle, B.H.; Cooper, G.J.S. Effect of high-fat meals and fatty acid saturation on postprandial levels of the hormones ghrelin and leptin in healthy men. Eur. J. Clin. Nutr. 2006, 60, 77–84. [Google Scholar] [CrossRef]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Phillips, S.A. Improvements in vascular health by a low-fat diet, but not a high-fat diet, are mediated by changes in adipocyte biology. Nutr. J. 2011, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Banks, A.S.; Davis, S.M.; Bates, S.H.; Myers, M.G. Activation of downstream signals by the long form of the leptin receptor. J. Biol. Chem. 2000, 275, 14563–14572. [Google Scholar] [CrossRef]
- Banks, R.E.; Forbes, M.A.; Patel, P.M.; Storr, M.; Hallam, S.; Clarke, D.; Novick, D.; Ingham, E.; Bowmer, C.; Southgate, J.; et al. Subcutaneous administration of recombinant glycosylated interleukin 6 in patients with cancer: Pharmacokinetics, pharmacodynamics and immunomodulatory effects. Cytokine 2000, 12, 388–396. [Google Scholar] [CrossRef]
- Bjørbæk, C.; Uotani, S.; Da Silva, B.; Flier, J.S. Divergent signaling capacities of the long and short isoforms of the leptin receptor. J. Biol. Chem. 1997, 272, 32686–32695. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.; Harvey, J. The neuronal actions of leptin and the implications for treating alzheimer’s disease. Pharmaceuticals 2021, 14, 52. [Google Scholar] [CrossRef]
- Kiernan, K.; MacIver, N.J. The Role of the Adipokine Leptin in Immune Cell Function in Health and Disease. Front. Immunol. 2021, 11, 622468. [Google Scholar] [CrossRef]
- Gorska, E.; Popko, K.; Stelmaszczyk-Emmel, A.; Ciepiela, O.; Kucharska, A.; Wasik, M. Leptin receptors. Eur. J. Med. Res. 2010, 15 (Suppl. 2), 50. [Google Scholar] [CrossRef]
- Voigtmann, F.; Wolf, P.; Landgraf, K.; Stein, R.; Kratzsch, J.; Schmitz, S.; Jamra, R.A.; Blüher, M.; Meiler, J.; Beck-Sickinger, A.G.; et al. Identification of a novel leptin receptor (LEPR) variant and proof of functional relevance directing treatment decisions in patients with morbid obesity. Metabolism 2021, 116, 154438. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.F.; Rocha, D.Q.C.; McIntyre, R.S.; Mesquita, L.M.; Köhler, C.A.; Hyphantis, T.N.; Sales, P.M.G.; Machado-Vieira, R.; Berk, M. Adipokines as emerging depression biomarkers: A systematic review and meta-analysis. J. Psychiatr. Res. 2014, 59, 28–37. [Google Scholar] [CrossRef]
- Reyes-Barrera, J.; Sainz-Escárrega, V.H.; Medina-Urritia, A.X.; Jorge-Galarza, E.; Osorio-Alonso, H.; Torres-Tamayo, M.; Leal-Escobar, G.; Posadas-Romero, C.; Torre-Villalvazo, I.; Juárez-Rojas, J.G. Dysfunctional adiposity index as a marker of adipose tissue morpho-functional abnormalities and metabolic disorders in apparently healthy subjects. Adipocyte 2021, 10, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Stieg, M.R.; Sievers, C.; Farr, O.; Stalla, G.K.; Mantzoros, C.S. Leptin: A hormone linking activation of neuroendocrine axes with neuropathology. Psychoneuroendocrinology 2015, 51, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.; Broxmeyer, H.E. Role for Leptin and Leptin Receptors in Stem Cells during Health and Diseases. Stem Cell Rev. Rep. 2021, 17, 511–522. [Google Scholar] [CrossRef]
- Fernández-Formoso, G.; Pérez-Sieira, S.; González-Touceda, D.; Dieguez, C.; Tovar, S. Leptin, 20 years of searching for glucose homeostasis. Life Sci. 2015, 140, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Cassioli, E.; Rossi, E.; Squecco, R.; Baccari, M.C.; Maggi, M.; Vignozzi, L.; Comeglio, P.; Gironi, V.; Lelli, L.; Rotella, F.; et al. Reward and psychopathological correlates of eating disorders: The explanatory role of leptin. Psychiatry Res. 2020, 290, 113071. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Duff, W.; Chizen, D.; Zello, G.A.; Chilibeck, P.D. The effect of a low glycemic index pulse-based diet on insulin sensitivity, insulin resistance, bone resorption and cardiovascular risk factors during bed rest. Nutrients 2019, 11, 2012. [Google Scholar] [CrossRef] [PubMed]
- Kolaczynski, J.W.; Considine, R.V.; Ohannesian, J.; Marco, C.; Opentanova, I.; Nyce, M.R.; Myint, M.; Caro, J.F. A Link with Ketogenesis but Not Ketones Themselves. Diabetes 1996, 45, 1511–1515. [Google Scholar] [CrossRef]
- Boyle, C.N.; Zheng, Y.; Lutz, T.A. Mediators of Amylin Action in Metabolic Control. J. Clin. Med. 2022, 11, 2207. [Google Scholar] [CrossRef]
- Grasso, P. Harnessing the Power of Leptin: The Biochemical Link Connecting Obesity, Diabetes, and Cognitive Decline. Front. Aging Neurosci. 2022, 14, 861350. [Google Scholar] [CrossRef]
- Andreoli, M.F.; Donato, J.; Cakir, I.; Perello, M. Leptin resensitisation: A reversion of leptin-resistant states. J. Endocrinol. 2019, 241, R81–R96. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. J. Clin. Epidemiol. 2021, 134, 178–189. [Google Scholar] [CrossRef]
- Hilton, L.K.; Loucks, A.B. Low energy availability, not exercise stress, suppresses the diurnal rhythm of leptin in healthy young women. Am. J. Physiol.-Endocrinol. Metab. 2000, 278, E43–E49. [Google Scholar] [CrossRef]
- Zaccaria, M.; Ermolao, A.; Roi, G.S.; Englaro, P.; Tegon, G.; Varnier, M. Leptin reduction after endurance races differing in duration and energy expenditure. Eur. J. Appl. Physiol. 2002, 87, 108–111. [Google Scholar] [CrossRef]
- Tsofliou, F.; Pitsiladis, Y.P.; Malkova, D.; Wallace, A.M.; Lean, M.E.J. Moderate physical activity permits acute coupling between serum leptin and appetite–satiety measures in obese women. Int. J. Obes. 2003, 27, 1332–1339. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, R.R.; Durand, R.J.; Acevedo, E.O.; Johnson, L.G.; Synovitz, L.B.; Kraemer, G.R.; Gimpel, T.; Castracane, V.D. Effects of High-Intensity Exercise on Leptin and Testosterone Concentrations in Well-Trained Males. Endocrine 2003, 21, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.C.; Sandretto, A.; Chung, Y.-C. Dieting is more effective in reducing weight but exercise is more effective in reducing fat during the early phase of a weight-reducing program in healthy humans. J. Nutr. Biochem. 2003, 14, 541–549. [Google Scholar] [CrossRef]
- Desgorces, F.D.F.; Chennaoui, M.; Gomez-Merino, D.; Drogou, C.; Guezennec, C.Y. Leptin response to acute prolonged exercise after training in rowers. Eur. J. Appl. Physiol. 2004, 91, 677–681. [Google Scholar] [CrossRef]
- Rubin, M.R.; Kraemer, W.J.; Maresh, C.M.; Volek, J.S.; Ratamess, N.A.; Vanheest, J.L.; Silvestre, R.; French, D.N.; Sharman, M.J.; Judelson, D.A.; et al. High-affinity growth hormone binding protein and acute heavy resistance exercise. Med. Sci. Sports Exerc. 2005, 37, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kobayashi, I.; Murakami, M. Effect of exercise on circulating adipokine levels in obese young women. Endocr. J. 2006, 53, 189–195. [Google Scholar] [CrossRef]
- Ara, I.; Perez-Gomez, J.; Vicente-Rodriguez, G.; Chavarren, J.; Dorado, C.; Calbert, J.A.L. Serum free testosterone, leptin and soluble leptin receptor changes in a 6-week strength-training programme. Br. J. Nutr. 2006, 96, 1053–1059. [Google Scholar] [CrossRef]
- Moro, C.; Pillard, F.; De Glisezinski, I.; Crampes, F.; Thalamas, C.; Harant, I.; Marques, M.A.; Lafontan, M.; Berlan, M. Sex differences in lipolysis-regulating mechanisms in overweight subjects: Effect of exercise intensity. Obesity 2007, 15, 2245–2255. [Google Scholar] [CrossRef]
- Kyriazis, G.A.; Caplan, J.D.; Lowndes, J.; Carpenter, R.L.; Dennis, K.E.; Sivo, S.A.; Angelopoulos, T.J. Moderate exercise-induced energy expenditure does not alter leptin levels in sedentary obese men. Clin. J. Sport Med. 2007, 17, 49–51. [Google Scholar] [CrossRef] [PubMed]
- König, D.; Deibert, P.; Frey, I.; Landmann, U.; Berg, A. Effect of meal replacement on metabolic risk factors in overweight and obese subjects. Ann. Nutr. Metab. 2008, 52, 74–78. [Google Scholar] [CrossRef]
- Volpe, S.L.; Kobusingye, H.; Bailur, S.; Stanek, E. Effect of Diet and Exercise on Body Composition, Energy Intake and Leptin Levels in Overweight Women and Men. J. Am. Coll. Nutr. 2008, 27, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Trapp, E.G.; Chisholm, D.J.; Freund, J.; Boutcher, S.H. The effects of high-intensity intermittent exercise training on fat loss and fasting insulin levels of young women. Int. J. Obes. 2008, 32, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.C.; Liu, Y.Y.; Lee, S.D.; Huang, C.Y.; Chien, K.Y.; Cheng, I.S.; Lin, C.Y.; Kuo, C.H. Effects of short-term detraining on measures of obesity and glucose tolerance in elite athletes. J. Sports Sci. 2008, 26, 919–925. [Google Scholar] [CrossRef]
- Jürimäe, J.; Rämson, R.; Mäestu, J.; Purge, P.; Jürimäe, T.; Arciero, P.J.; Von Duvillard, S.P. Plasma visfatin and ghrelin response to prolonged sculling in competitive male rowers. Med. Sci. Sports Exerc. 2009, 41, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.H.Y.; Bushnell, D.; Cannon, D.T.; Kern, M. Appetite regulation via exercise prior or subsequent to high-fat meal consumption. Appetite 2009, 52, 193–198. [Google Scholar] [CrossRef]
- Hagobian, T.A.; Sharoff, C.G.; Stephens, B.R.; Wade, G.N.; Silva, J.E.; Chipkin, S.R.; Braun, B. Effects of exercise on energy-regulating hormones and appetite in men and women. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2009, 296, R233–R242. [Google Scholar] [CrossRef]
- Guadalupe-Grau, A.; Perez-Gomez, J.; Olmedillas, H.; Chavarren, J.; Dorado, C.; Santana, A.; Serrano-Sanchez, J.A.; Calbet, J.A.L. Strength training combined with plyometric jumps in adults: Sex differences in fat-bone axis adaptations. J. Appl. Physiol. 2009, 106, 1100–1111. [Google Scholar] [CrossRef]
- Bergouignan, A.; Momken, I.; Schoeller, D.A.; Normand, S.; Zahariev, A.; Lescure, B.; Simon, C.; Blanc, S. Regulation of energy balance during long-term physical inactivity induced by bed rest with and without exercise training. J. Clin. Endocrinol. Metab. 2010, 95, 1045–1053. [Google Scholar] [CrossRef]
- Sartor, F.; De Morree, H.M.; Matschke, V.; Marcora, S.M.; Milousis, A.; Thom, J.M.; Kubis, H.P. High-intensity exercise and carbohydrate-reduced energy-restricted diet in obese individuals. Eur. J. Appl. Physiol. 2010, 110, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.A.; Watras, A.C.; Paton, C.M.; Wegner, F.H.; Adams, A.K.; Schoeller, D.A. Impact of exercise and dietary fatty acid composition from a high-fat diet on markers of hunger and satiety. Appetite 2011, 56, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, R.R.; Francois, M.R.; Sehgal, K.; Sirikul, B.; Valverde, R.A.; Castracane, V.D. Amylin and selective glucoregulatory peptide alterations during prolonged exercise. Med. Sci. Sports Exerc. 2011, 43, 1451–1456. [Google Scholar] [CrossRef]
- Moran, C.N.; Barwell, N.D.; Malkova, D.; Cleland, S.J.; McPhee, I.; Packard, C.J.; Zammit, V.A.; Gill, J.M.R. Effects of diabetes family history and exercise training on the expression of adiponectin and leptin and their receptors. Metabolism 2011, 60, 206–214. [Google Scholar] [CrossRef]
- Balaguera-Cortes, L.; Wallman, K.E.; Fairchild, T.J.; Guelfi, K.J. Energy intake and appetite-related hormones following acute aerobic and resistance exercise. Appl. Physiol. Nutr. Metab. 2011, 36, 958–966. [Google Scholar] [CrossRef]
- Rosa, G.; Dantas, E.H.M.; de Mello, D.B. The response of serum leptin, cortisol and zinc concentrations to concurrent training. Hormones 2011, 10, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Arikawa, A.Y.; Thomas, W.; Schmitz, K.H.; Kurzer, M.S. Sixteen weeks of exercise reduces C-reactive protein levels in young women. Med. Sci. Sports Exerc. 2011, 43, 1002–1009. [Google Scholar] [CrossRef]
- Kelly, K.R.; Blaszczak, A.; Haus, J.M.; Patrick-Melin, A.; Fealy, C.E.; Solomon, T.P.; Kalinski, M.I.; Kirwan, J.P. A 7-d exercise program increases high-molecular weight adiponectin in obese adults. Med. Sci. Sports Exerc. 2012, 44, 69–74. [Google Scholar] [CrossRef]
- Numao, S.; Sasai, H.; Nomata, Y.; Matsuo, T.; Eto, M.; Tsujimoto, T.; Tanaka, K. Effects of exercise training on circulating retinol-binding protein 4 and cardiovascular disease risk factors in obese men. Obes. Facts 2012, 5, 845–855. [Google Scholar] [CrossRef]
- Zaccaria, M.; Ermolao, A.; Brugin, E.; Bergamin, M. Plasma leptin and energy expenditure during prolonged, moderate intensity, treadmill exercise. J. Endocrinol. Investig. 2013, 36, 396–401. [Google Scholar] [CrossRef]
- Martins, C.; Kulseng, B.; Rehfeld, J.F.; King, N.A.; Blundell, J.E. Effect of chronic exercise on appetite control in overweight and obese individuals. Med. Sci. Sports Exerc. 2013, 45, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Morishima, T.; Kurihara, T.; Hamaoka, T.; Goto, K. Whole body, regional fat accumulation, and appetite-related hormonal response after hypoxic training. Clin. Physiol. Funct. Imaging 2014, 34, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Mendham, A.E.; Duffield, R.; Marino, F.; Coutts, A.J. Small-sided games training reduces CRP, IL-6 and leptin in sedentary, middle-aged men. Eur. J. Appl. Physiol. 2014, 114, 2289–2297. [Google Scholar] [CrossRef] [PubMed]
- Debevec, T.; Simpson, E.J.; Macdonald, I.A.; Eiken, O.; Mekjavic, I.B. Exercise training during normobaric hypoxic confinement does not alter hormonal appetite regulation. PLoS ONE 2014, 9, e98874. [Google Scholar] [CrossRef] [PubMed]
- Ahmadizad, S.; Ghorbani, S.; Ghasemikaram, M.; Bahmanzadeh, M. Effects of short-term nonperiodized, linear periodized and daily undulating periodized resistance training on plasma adiponectin, leptin and insulin resistance. Clin. Biochem. 2014, 47, 417–422. [Google Scholar] [CrossRef]
- Sim, A.Y.; Wallman, K.E.; Fairchild, T.J.; Guelfi, K.J. Effects of high-intensity intermittent exercise training on appetite regulation. Med. Sci. Sports Exerc. 2015, 47, 2441–2449. [Google Scholar] [CrossRef]
- Kim, Y.S.; Nam, J.S.; Yeo, D.W.; Kim, K.R.; Suh, S.H.; Ahn, C.W. The effects of aerobic exercise training on serum osteocalcin, adipocytokines and insulin resistance on obese young males. Clin. Endocrinol. 2015, 82, 686–694. [Google Scholar] [CrossRef] [PubMed]
- King, J.A.; Garnham, J.O.; Jackson, A.P.; Kelly, B.M.; Xenophontos, S.; Nimmo, M.A. Appetite-regulatory hormone responses on the day following a prolonged bout of moderate-intensity exercise. Physiol. Behav. 2015, 141, 23–31. [Google Scholar] [CrossRef]
- Douglas, J.A.; King, J.A.; McFarlane, E.; Baker, L.; Bradley, C.; Crouch, N.; Hill, D.; Stensel, D.J. Appetite, appetite hormone and energy intake responses to two consecutive days of aerobic exercise in healthy young men. Appetite 2015, 92, 57–65. [Google Scholar] [CrossRef]
- Racil, G.; Zouhal, H.; Elmontassar, W.; Abderrahmane, A.B.; De Sousa, M.V.; Chamari, K.; Amri, M.; Coquart, J.B. Plyometric exercise combined with high-intensity interval training improves metabolic abnormalities in young obese females more so than interval training alone. Appl. Physiol. Nutr. Metab. 2016, 41, 103–109. [Google Scholar] [CrossRef]
- Kong, Z.; Sun, S.; Liu, M.; Shi, Q. Short-Term High-Intensity Interval Training on Body Composition and Blood Glucose in Overweight and Obese Young Women. J. Diabetes Res. 2016, 2016, 4073618. [Google Scholar] [CrossRef]
- Cameron, J.D.; Goldfield, G.S.; Riou, M.È.; Finlayson, G.S.; Blundell, J.E.; Doucet, É. Energy depletion by diet or aerobic exercise alone: Impact of energy deficit modality on appetite parameters. Am. J. Clin. Nutr. 2016, 103, 1008–1016. [Google Scholar] [CrossRef]
- Vardar, S.A.; Karaca, A.; Güldiken, S.; Palabıyık, O.; Süt, N.; Demir, A.M. High-intensity interval training acutely alters plasma adipokine levels in young overweight/obese women. Arch. Physiol. Biochem. 2018, 124, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, R.S.; Panissa, V.L.G.; Inoue, D.S.; Campos, E.Z.; Monteiro, P.A.; Giglio, B.D.M.; Pimentel, G.D.; Hofmann, P.; Lira, F.S. Impact to short-term high intensity intermittent training on different storages of body fat, leptin and soluble leptin receptor levels in physically active non-obese men: A pilot investigation. Clin. Nutr. ESPEN 2018, 28, 186–192. [Google Scholar] [CrossRef]
- Tremblay, A.; Dutheil, F.; Drapeau, V.; Metz, L.; Lesour, B.; Chapier, R.; Pereira, B.; Verney, J.; Baker, J.S.; Vinet, A.; et al. Long-term effects of high-intensity resistance and endurance exercise on plasma leptin and ghrelin in overweight individuals: The RESOLVE Study. Appl. Physiol. Nutr. Metab. 2019, 44, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Lee, S.; Gulseth, H.L.; Jensen, J.; Drevon, C.A.; Birkeland, K.I. Soluble Leptin Receptor Predicts Insulin Sensitivity and Correlates with Upregulation of Metabolic Pathways in Men. J. Clin. Endocrinol. Metab. 2018, 103, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Middelbeek, R.J.W.; Motiani, P.; Brandt, N.; Nigro, P.; Zheng, J.; Virtanen, K.A.; Kalliokoski, K.K.; Hannukainen, J.C.; Goodyear, L.J. Exercise intensity regulates cytokine and klotho responses in men. Nutr. Diabetes 2021, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Zaman, G.S.; Abohashrh, M.; Ahmad, I.I.; Dera, A.A.; Alshahrani, M.S.; Ahmad, I.I.; Alam, M.M.; Mahmood, S.E.; Mansuri, N.; Irfan, S.; et al. The Impact of Body Resistance Training Exercise on Biomedical Profile at High Altitude: A Randomized Controlled Trial. Biomed Res. Int. 2021, 2021, 6684167. [Google Scholar] [CrossRef] [PubMed]
- Desgorces, F.D.; Chennaoui, M.; Gomez-Merino, D.; Drogou, C.; Bonneau, D.; Guezennec, C.Y. Leptin, catecholamines and free fatty acids related to reduced recovery delays after training. Eur. J. Appl. Physiol. 2004, 93, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Romero, F.; Rodríguez-Morán, M. Magnesium improves the beta-cell function to compensate variation of insulin sensitivity: Double-blind, randomized clinical trial. Eur. J. Clin. Investig. 2011, 41, 405–410. [Google Scholar] [CrossRef]
- Tsai, S.-H.H.; Cheng, H.-C.C.; Liu, H.-W.W. Effects of volume-matched resistance training with different loads on glycemic control, inflammation, and body composition in prediabetic older adults. Appl. Physiol. Nutr. Metab. 2021, 46, 1400–1406. [Google Scholar] [CrossRef]
- Khalil, R.B.; El Hachem, C. Adiponectin in eating disorders. Eat. Weight Disord. 2014, 19, 3–10. [Google Scholar] [CrossRef]
- Tang, N.; Zhang, X.; Chen, D.; Li, Z. The controversial role of adiponectin in appetite regulation of animals. Nutrients 2021, 13, 3387. [Google Scholar] [CrossRef]
- Yu, L.; Wang, J.; Xia, Y.; Hou, W.; Yao, X.; Guo, Y.; Wang, J.; Wei, H. Adiponectin Promotes Neurogenesis after Transient Cerebral Ischemia through STAT3 Mediated BDNF Upregulation in Astrocytes. Neurochem. Res. 2022, 48, 641–657. [Google Scholar] [CrossRef] [PubMed]
- de Assis, G.G.; Murawska-Ciałowicz, E. Leptin—A potential bridge between fat metabolism and the brain’s vulnerability to neuropsychiatric disorders: A systematic review. J. Clin. Med. 2021, 10, 5714. [Google Scholar] [CrossRef] [PubMed]
- Alghadir, A.H.; Gabr, S.A.; Almomani, M.; Almomani, F.; Tse, C. Adiponectin and Nitric Oxide Deficiency-Induced Cognitive Impairment in Fatigued Home-Resident in Mature and Older Adults: A Case-Control Study. Pain Res. Manag. 2022, 2022, 7480579. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Lim, J.H.; Kim, E.N.; Hong, Y.A.; Park, H.-J.; Chung, S.; Choi, B.S.; Kim, Y.-S.; Park, J.Y.; Kim, H.W.; et al. Adiponectin receptor agonist ameliorates cardiac lipotoxicity via enhancing ceramide metabolism in type 2 diabetic mice. Cell Death Dis. 2022, 13, 282. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Du, T.; Li, C.; Yang, G. STAT3 phosphorylation in central leptin resistance. Nutr. Metab. 2021, 18, 39. [Google Scholar] [CrossRef]
- Socol, C.T.; Chira, A.; Martinez-Sanchez, M.A.; Nuñez-Sanchez, M.A.; Maerescu, C.M.; Mierlita, D.; Rusu, A.V.; Ruiz-Alcaraz, A.J.; Trif, M.; Ramos-Molina, B. Leptin Signaling in Obesity and Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 4713. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Assis, G.G.; Murawska-Ciałowicz, E. Exercise and Weight Management: The Role of Leptin—A Systematic Review and Update of Clinical Data from 2000–2022. J. Clin. Med. 2023, 12, 4490. https://doi.org/10.3390/jcm12134490
de Assis GG, Murawska-Ciałowicz E. Exercise and Weight Management: The Role of Leptin—A Systematic Review and Update of Clinical Data from 2000–2022. Journal of Clinical Medicine. 2023; 12(13):4490. https://doi.org/10.3390/jcm12134490
Chicago/Turabian Stylede Assis, Gilmara Gomes, and Eugenia Murawska-Ciałowicz. 2023. "Exercise and Weight Management: The Role of Leptin—A Systematic Review and Update of Clinical Data from 2000–2022" Journal of Clinical Medicine 12, no. 13: 4490. https://doi.org/10.3390/jcm12134490
APA Stylede Assis, G. G., & Murawska-Ciałowicz, E. (2023). Exercise and Weight Management: The Role of Leptin—A Systematic Review and Update of Clinical Data from 2000–2022. Journal of Clinical Medicine, 12(13), 4490. https://doi.org/10.3390/jcm12134490