
Adapting Human-Based Transcutaneous Spinal Cord Stimulation to Develop a Clinically Relevant Animal Model


Abstract
1. Introduction
2. Materials and Methods
2.1. Intact Animals—Experiments
2.1.1. Transcutaneous Electrode Fabrication and Stimulation Set-Up
2.1.2. Stimulation and Recordings
2.1.3. Data Analysis
2.1.4. Statistical Analysis
2.2. Spinal Cord-Injured Animals—Procedures and Experiments
2.2.1. Surgical Procedures and Postoperative Care
2.2.2. Repeated Transcutaneous Stimulation in SCI Animals
2.2.3. Terminal Experiments and Analysis
2.3. Human Experiments
2.3.1. Stimulation and Recordings
2.3.2. Data Analysis and Statistics
3. Results
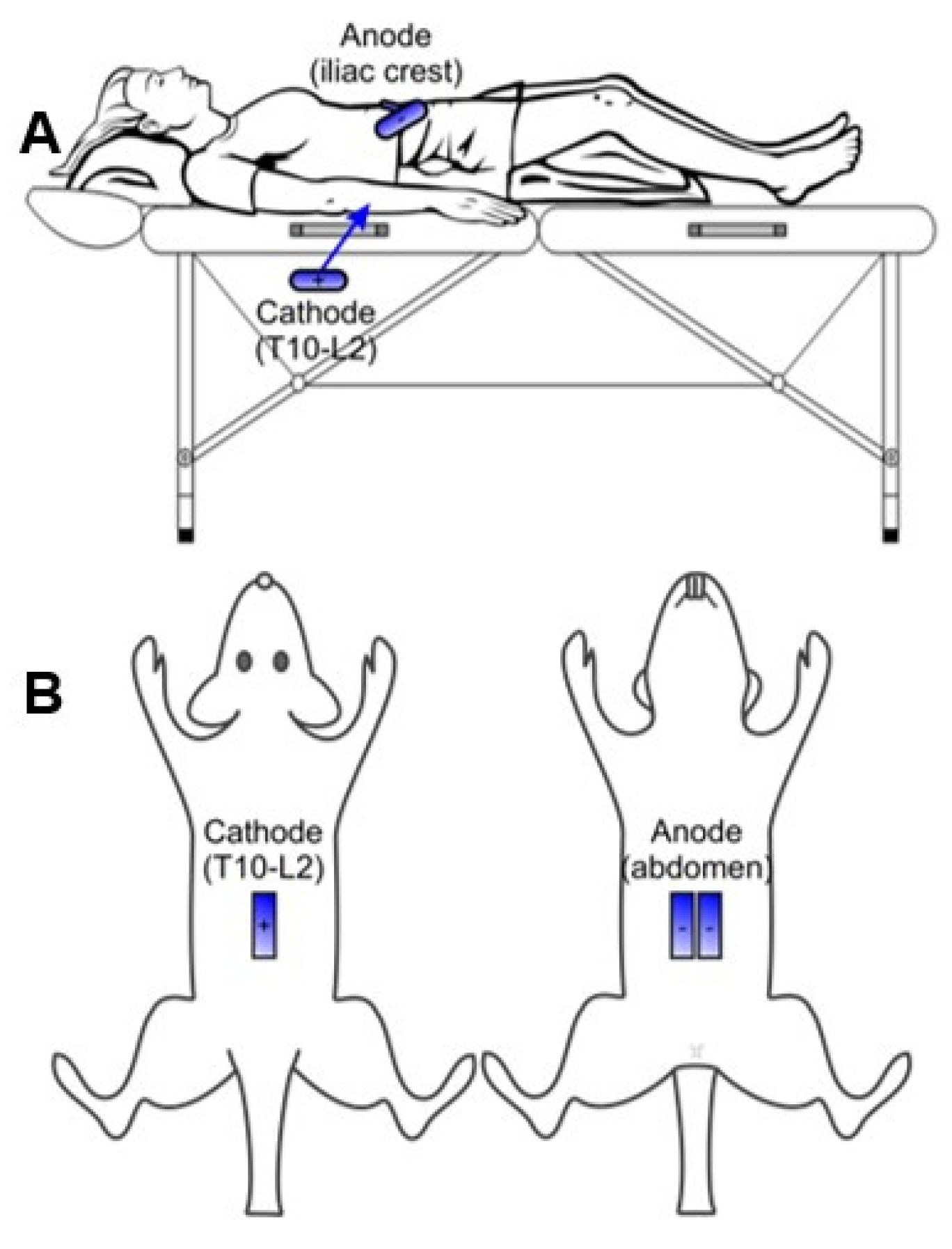
3.1. Adapting Electrode Configurations
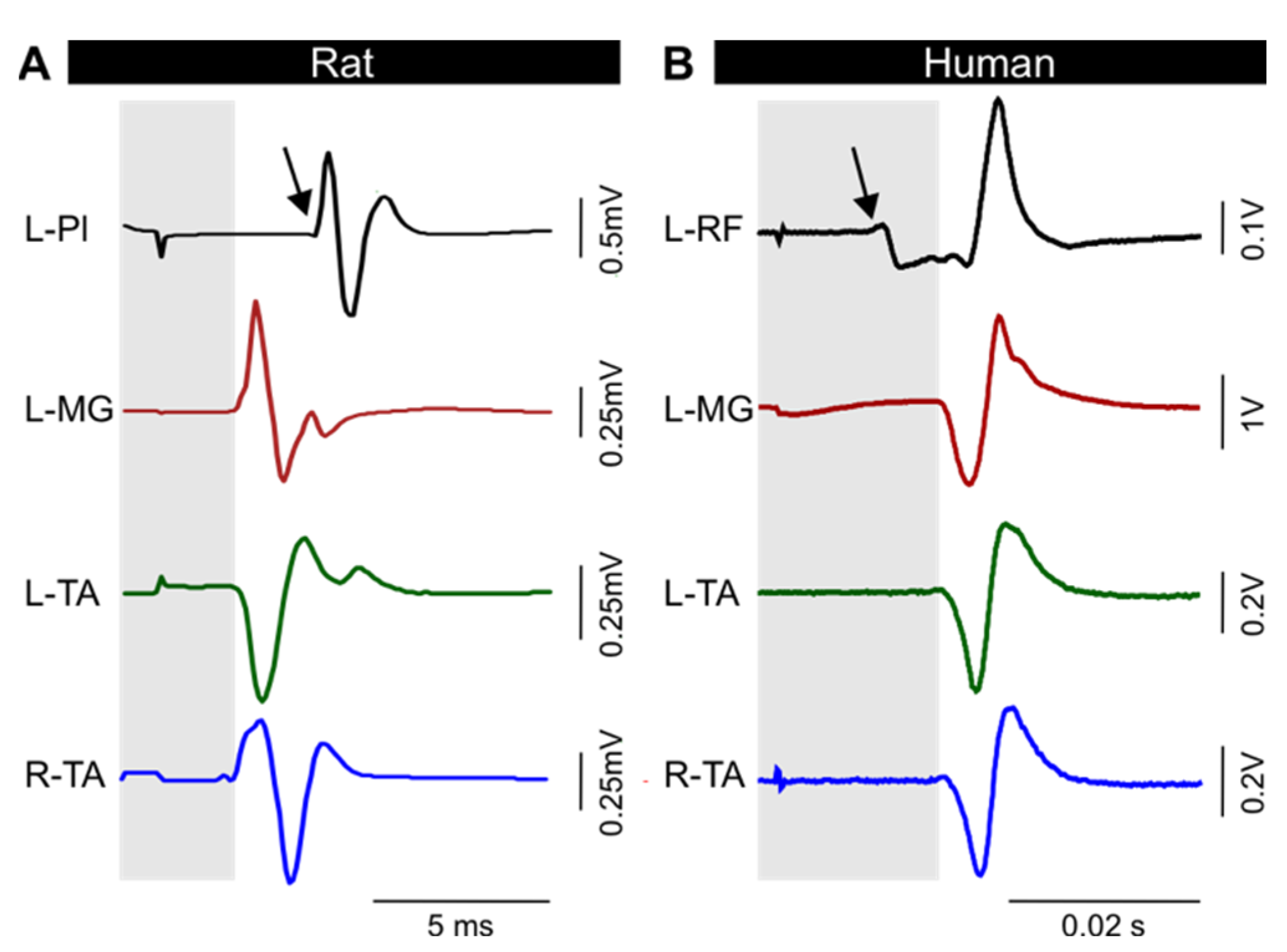
3.2. TEPs Are Characteristically Similar across Muscles and between Models
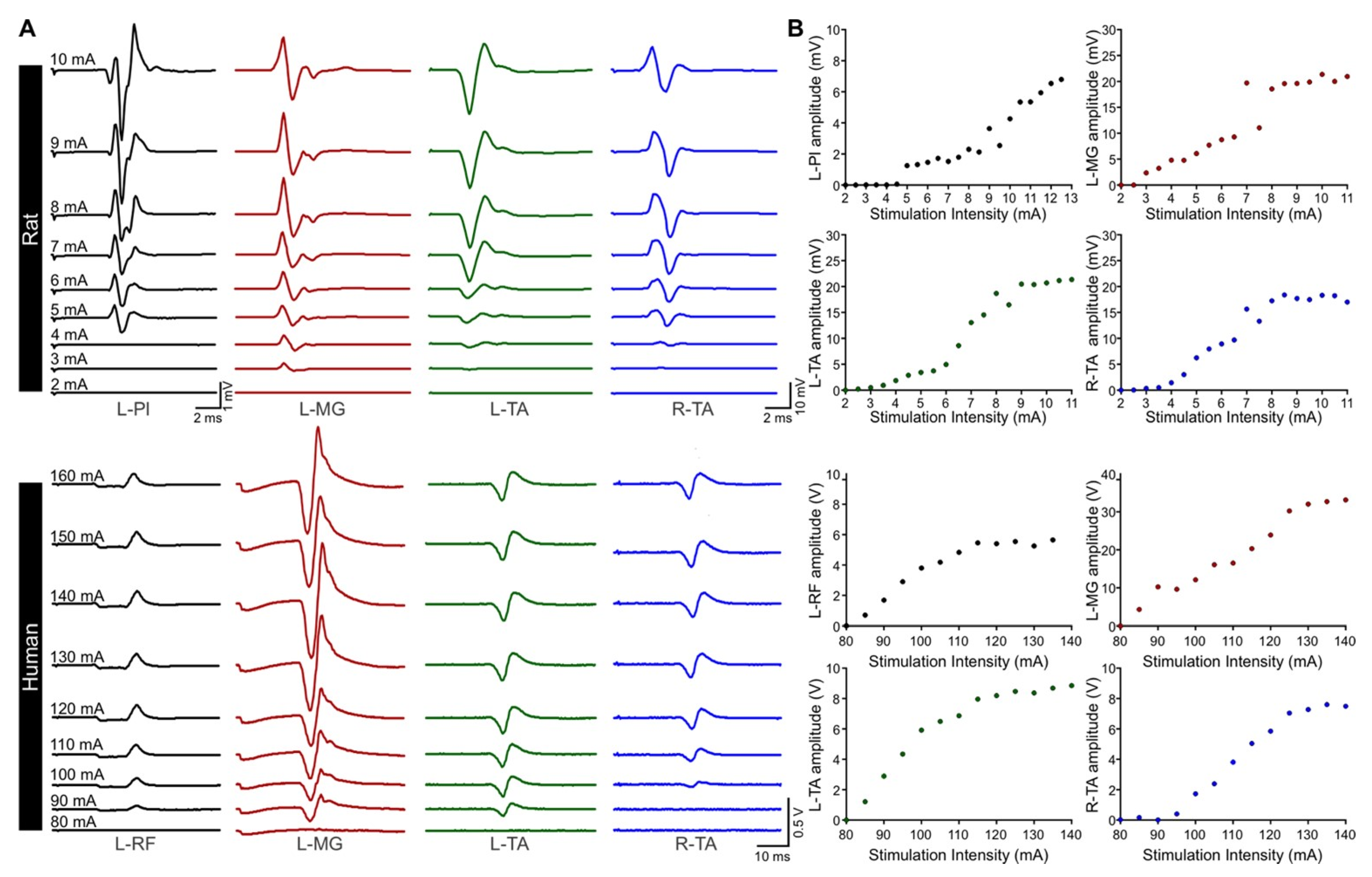
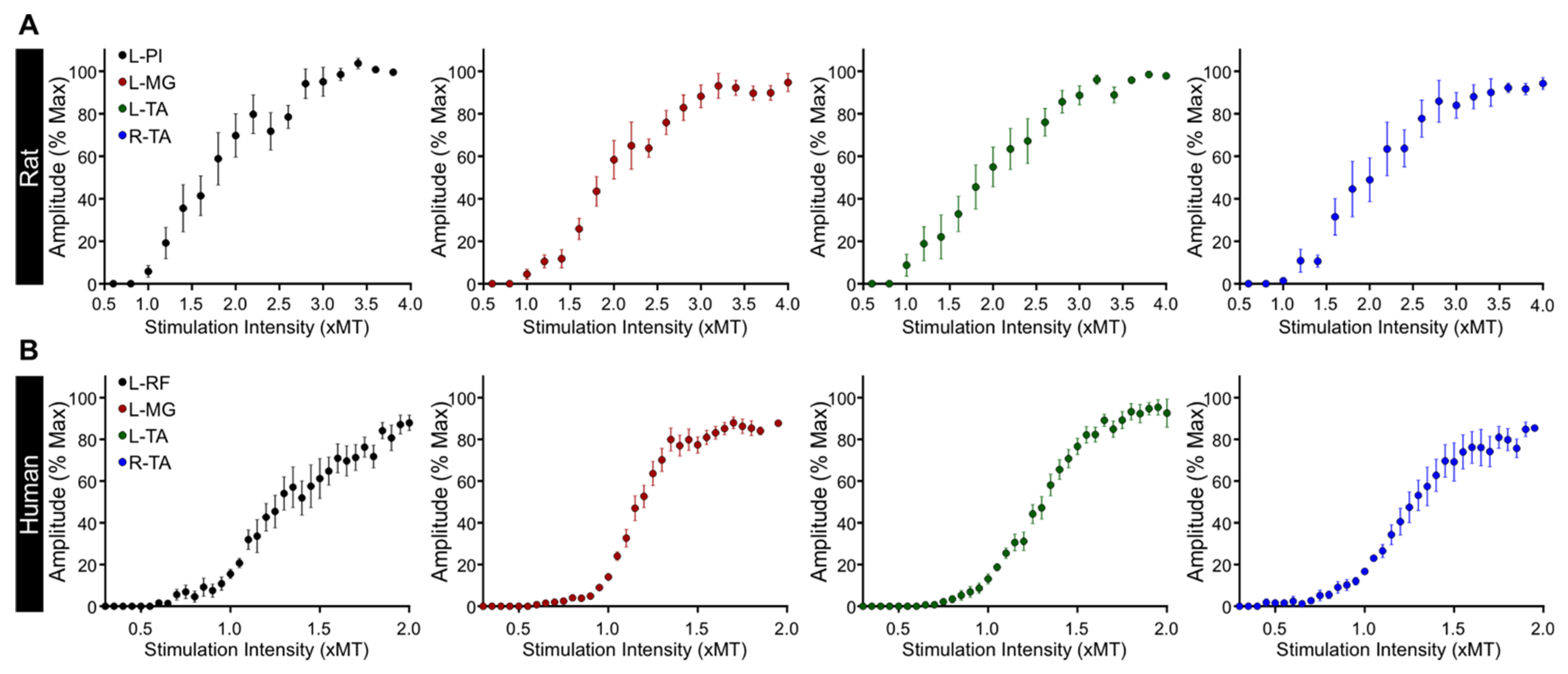
3.3. Excitability of Motor Pools in Response to tSCS
3.4. Latency and Duration of TEPs
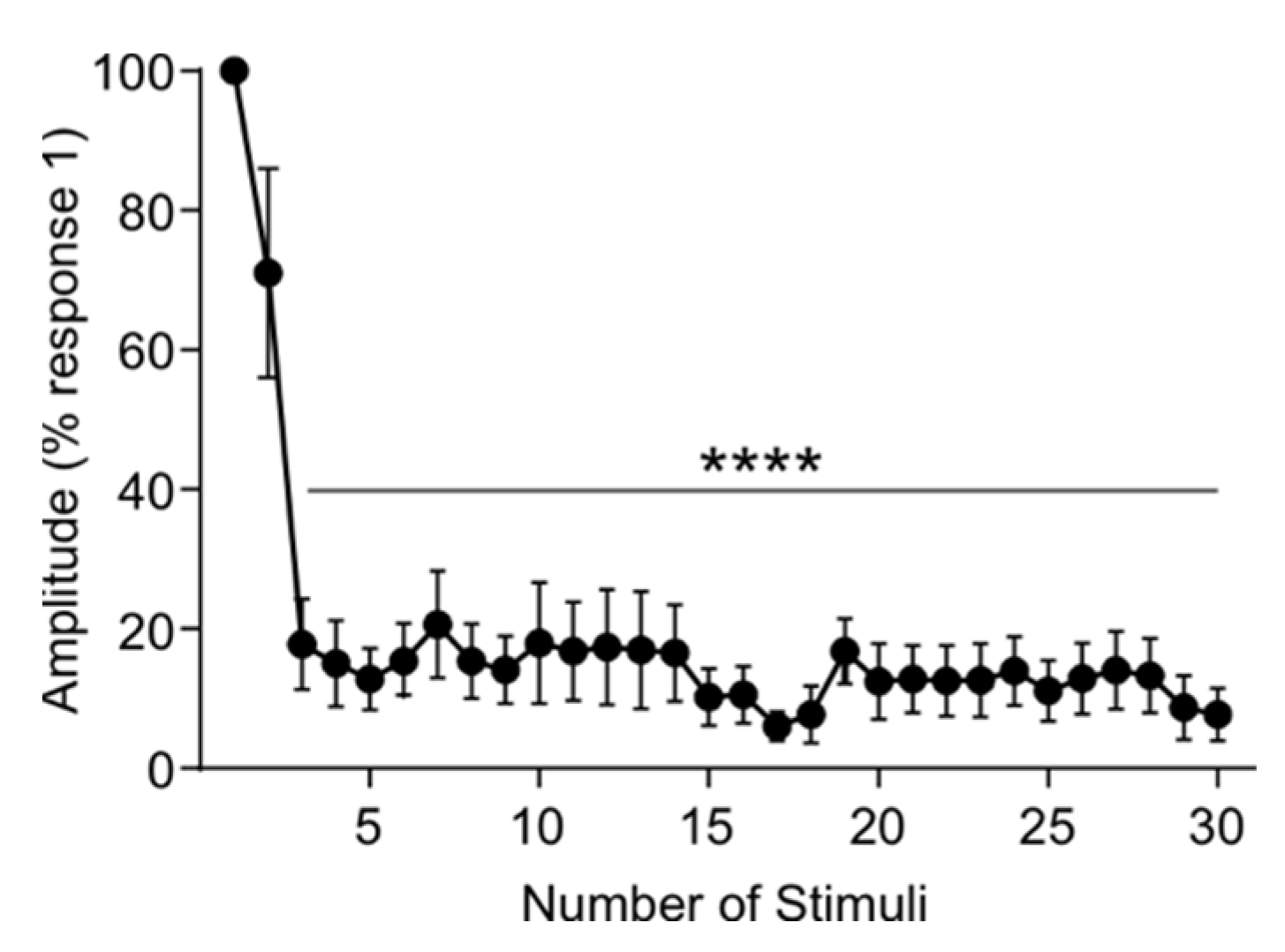
3.5. Rat tSCS Activates Primary Afferents
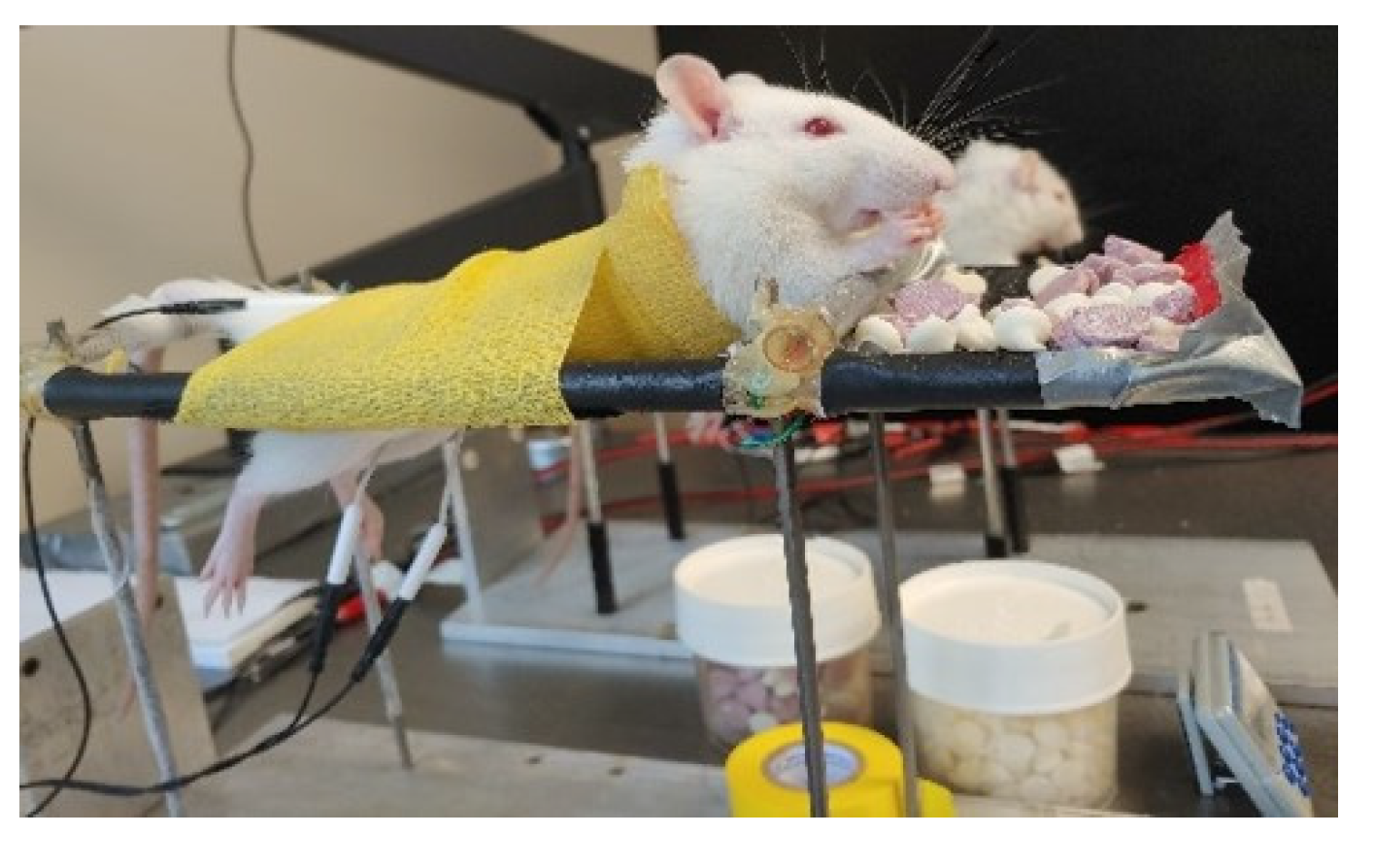
3.6. Repeated tSCS in the Awake Rat Is Feasible, Tolerable, and Safe
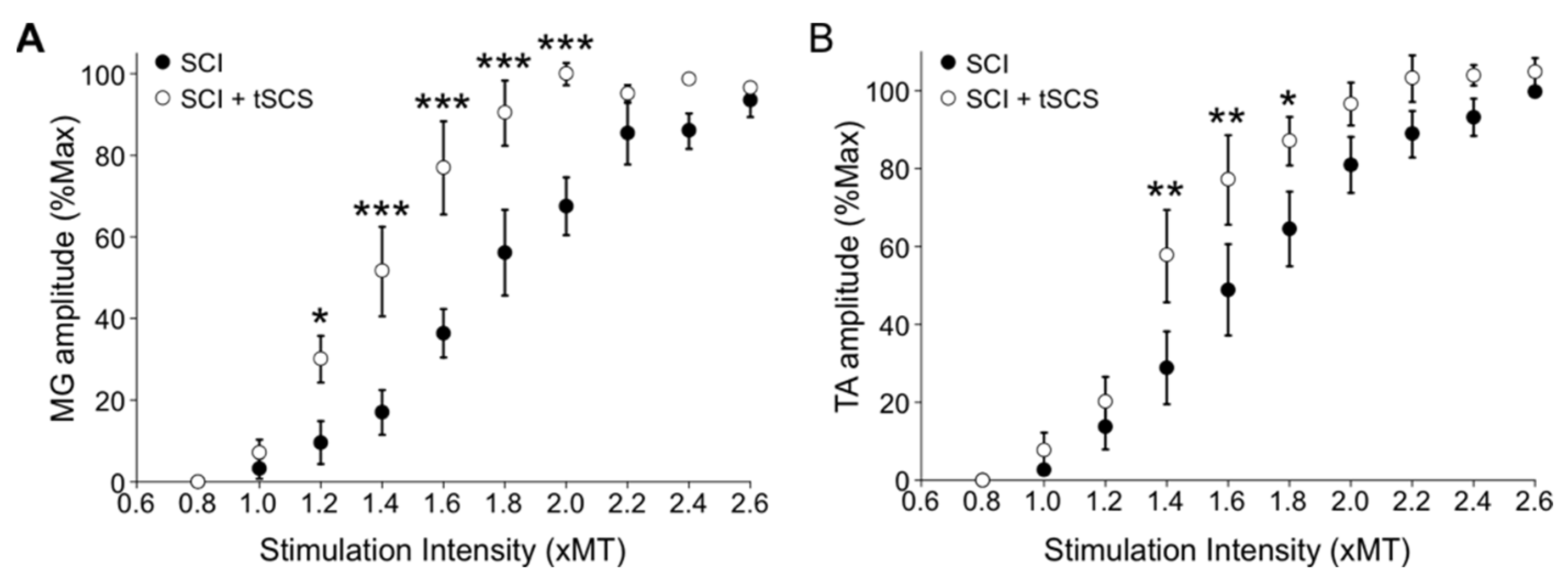
3.7. Repeated tSCS Increases Motor Output in Ankle Muscles of the Spinalized Rat
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hofstoetter, U.S.; Knikou, M.; Guertin, P.A.; Minassian, K. Probing the Human Spinal Locomotor Circuits by Phasic Step-Induced Feedback and by Tonic Electrical and Pharmacological Neuromodulation. Curr. Pharm. Des. 2017, 23, 1805–1820. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.; Knikou, M. A Review on Locomotor Training after Spinal Cord Injury: Reorganization of Spinal Neuronal Circuits and Recovery of Motor Function. Neural Plast. 2016, 2016, 1216258. [Google Scholar] [CrossRef] [PubMed]
- Einhorn, J.; Li, A.; Hazan, R.; Knikou, M. Cervicothoracic Multisegmental Transpinal Evoked Potentials in Humans. PLoS ONE 2013, 8, e76940. [Google Scholar] [CrossRef] [PubMed]
- Minassian, K.; Persy, I.; Rattay, F.; Dimitrijevic, M.R.; Ms, C.H.; Kern, H. Posterior root–muscle reflexes elicited by transcutaneous stimulation of the human lumbosacral cord. Muscle Nerve 2006, 35, 327–336. [Google Scholar] [CrossRef]
- Krenn, M.; Toth, A.; Danner, S.M.; Hofstoetter, U.S.; Minassian, K.; Mayr, W. Selectivity of transcutaneous stimulation of lumbar posterior roots at different spinal levels in humans. Biomed. Technol. 2013, 58 (Suppl. 1). [Google Scholar] [CrossRef]
- Knikou, M. Neurophysiological characterization of transpinal evoked potentials in human leg muscles. Bioelectromagnetics 2013, 34, 630–640. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Gorodnichev, R.; Machueva, E.; Pivovarova, E.; Semyenov, D.; Savochin, A.; Roy, R.R.; Edgerton, V.R. Novel and Direct Access to the Human Locomotor Spinal Circuitry. J. Neurosci. 2010, 30, 3700–3708. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Gorodnichev, R.; Moshonkina, T.; Sayenko, D.; Gad, P.; Edgerton, V.R. Transcutaneous electrical spinal-cord stimulation in humans. Ann. Phys. Rehabil. Med. 2015, 58, 225–231. [Google Scholar] [CrossRef]
- Gerasimenko, Y.P.; Lu, D.C.; Modaber, M.; Zdunowski, S.; Gad, P.; Sayenko, D.G.; Morikawa, E.; Haakana, P.; Ferguson, A.; Roy, R.R.; et al. Noninvasive Reactivation of Motor Descending Control after Paralysis. J. Neurotrauma 2015, 32, 1968–1980. [Google Scholar] [CrossRef]
- Gerasimenko, Y.; Gorodnichev, R.; Puhov, A.; Moshonkina, T.; Savochin, A.; Selionov, V.; Roy, R.R.; Lu, D.C.; Edgerton, V.R. Initiation and modulation of locomotor circuitry output with multisite transcutaneous electrical stimulation of the spinal cord in noninjured humans. J. Neurophysiol. 2015, 113, 834–842. [Google Scholar] [CrossRef]
- Gad, P.; Gerasimenko, Y.; Zdunowski, S.; Turner, A.; Sayenko, D.; Lu, D.C.; Edgerton, V.R. Weight Bearing Over-ground Stepping in an Exoskeleton with Non-invasive Spinal Cord Neuromodulation after Motor Complete Paraplegia. Front. Neurosci. 2017, 11, 333. [Google Scholar] [CrossRef] [PubMed]
- Sayenko, D.G.; Rath, M.; Ferguson, A.; Burdick, J.W.; Havton, L.A.; Edgerton, V.R.; Gerasimenko, Y.P. Self-Assisted Standing Enabled by Non-Invasive Spinal Stimulation after Spinal Cord Injury. J. Neurotrauma 2019, 36, 1435–1450. [Google Scholar] [CrossRef] [PubMed]
- Gad, P.; Lee, S.; Terrafranca, N.; Zhong, H.; Turner, A.; Gerasimenko, Y.; Edgerton, V.R. Non-Invasive Activation of Cervical Spinal Networks after Severe Paralysis. J. Neurotrauma 2018, 35, 2145–2158. [Google Scholar] [CrossRef] [PubMed]
- Inanici, F.; Samejima, S.; Gad, P.; Edgerton, V.R.; Hofstetter, C.P.; Moritz, C.T. Transcutaneous Electrical Spinal Stimulation Promotes Long-Term Recovery of Upper Extremity Function in Chronic Tetraplegia. IEEE Trans. Neural Syst. Rehabil. Eng. 2018, 26, 1272–1278. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.S.; Danner, S.M.; Mayr, W.; Bruce, J.A.; McKay, W.B.; Tansey, K.E. Spinal Rhythm Generation by Step-Induced Feedback and Transcutaneous Posterior Root Stimulation in Complete Spinal Cord–Injured Individuals. Neurorehabilit. Neural Repair 2015, 30, 233–243. [Google Scholar] [CrossRef]
- Murray, L.M.; Knikou, M. Transspinal stimulation increases motoneuron output of multiple segments in human spinal cord injury. PLoS ONE 2019, 14, e0213696. [Google Scholar] [CrossRef]
- Knikou, M.; Murray, L.M. Repeated transspinal stimulation decreases soleus H-reflex excitability and restores spinal inhibition in human spinal cord injury. PLoS ONE 2019, 14, e0223135. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; Freundl, B.; Danner, S.M.; Krenn, M.J.; Mayr, W.; Binder, H.; Minassian, K. Transcutaneous Spinal Cord Stimulation Induces Temporary Attenuation of Spasticity in Individuals with Spinal Cord Injury. J. Neurotrauma 2020, 37, 481–493. [Google Scholar] [CrossRef]
- Hofstoetter, U.S.; McKay, W.B.; Tansey, K.E.; Mayr, W.; Kern, H.; Minassian, K. Modification of spasticity by transcutaneous spinal cord stimulation in individuals with incomplete spinal cord injury. J. Spinal Cord Med. 2013, 37, 202–211. [Google Scholar] [CrossRef]
- Mekhael, W.; Begum, S.; Samaddar, S.; Hassan, M.; Toruno, P.; Ahmed, M.; Gorin, A.; Maisano, M.; Ayad, M.; Ahmed, Z. Repeated anodal trans-spinal direct current stimulation results in long-term reduction of spasticity in mice with spinal cord injury. J. Physiol. 2019, 597, 2201–2223. [Google Scholar] [CrossRef]
- Zareen, N.; Shinozaki, M.; Ryan, D.; Alexander, H.; Amer, A.; Truong, D.; Khadka, N.; Sarkar, A.; Naeem, S.; Bikson, M.; et al. Motor cortex and spinal cord neuromodulation promote corticospinal tract axonal outgrowth and motor recovery after cervical contusion spinal cord injury. Exp. Neurol. 2017, 297, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Truong, D.Q.; Bikson, M.; Martin, J.H. Transspinal direct current stimulation immediately modifies motor cortex sensorimotor maps. J. Neurophysiol. 2015, 113, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Cohen, L.G.; Wassermann, E.M.; Priori, A.; Lang, N.; Antal, A.; Paulus, W.; Hummel, F.; Boggio, P.S.; Fregni, F.; et al. Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 2008, 1, 206–223. [Google Scholar] [CrossRef] [PubMed]
- Priori, A.; Ciocca, M.; Parazzini, M.; Vergari, M.; Ferrucci, R. Transcranial cerebellar direct current stimulation and transcutaneous spinal cord direct current stimulation as innovative tools for neuroscientists. J. Physiol. 2014, 592, 3345–3369. [Google Scholar] [CrossRef]
- Matsumoto, H.; Ugawa, Y. Adverse events of tDCS and tACS: A review. Clin. Neurophysiol. Pract. 2017, 2, 19–25. [Google Scholar] [CrossRef]
- Murray, L.M.; Tahayori, B.; Knikou, M. Transspinal Direct Current Stimulation Produces Persistent Plasticity in Human Motor Pathways. Sci. Rep. 2018, 8, 717. [Google Scholar] [CrossRef]
- Sharif, H.; Alexander, H.; Azam, A.; Martin, J.H. Dual motor cortex and spinal cord neuromodulation improves rehabilitation efficacy and restores skilled locomotor function in a rat cervical contusion injury model. Exp. Neurol. 2021, 341, 113715. [Google Scholar] [CrossRef]
- Ellaway, P. Cumulative sum technique and its application to the analysis of peristimulus time histograms. Electroencephalogr. Clin. Neurophysiol. 1978, 45, 302–304. [Google Scholar] [CrossRef]
- Brinkworth, R.; Türker, K.S. A method for quantifying reflex responses from intra-muscular and surface electromyogram. J. Neurosci. Methods 2003, 122, 179–193. [Google Scholar] [CrossRef]
- Mishra, A.M.; Pal, A.; Gupta, D.; Carmel, J.B. Paired motor cortex and cervical epidural electrical stimulation timed to converge in the spinal cord promotes lasting increases in motor responses. J. Physiol. 2017, 595, 6953–6968. [Google Scholar] [CrossRef]
- Kumru, H.; Rodríguez-Cañón, M.; Edgerton, V.; García, L.; Flores, A.; Soriano, I.; Opisso, E.; Gerasimenko, Y.; Navarro, X.; García-Alías, G.; et al. Transcutaneous Electrical Neuromodulation of the Cervical Spinal Cord Depends Both on the Stimulation Intensity and the Degree of Voluntary Activity for Training. A Pilot Study. J. Clin. Med. 2021, 10, 3278. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Garcia-Alias, G.; Jin, B.; Keyes, J.; Zhong, H.; Roy, R.R.; Gerasimenko, Y.; Lu, D.C.; Edgerton, V.R. Electrical neuromodulation of the cervical spinal cord facilitates forelimb skilled function recovery in spinal cord injured rats. Exp. Neurol. 2017, 291, 141–150. [Google Scholar] [CrossRef]
- Fadeev, F.; Eremeev, A.; Bashirov, F.; Shevchenko, R.; Izmailov, A.; Markosyan, V.; Sokolov, M.; Kalistratova, J.; Khalitova, A.; Garifulin, R.; et al. Combined Supra- and Sub-Lesional Epidural Electrical Stimulation for Restoration of the Motor Functions after Spinal Cord Injury in Mini Pigs. Brain Sci. 2020, 10, 744. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M. Transpinal and Transcortical Stimulation Alter Corticospinal Excitability and Increase Spinal Output. PLoS ONE 2014, 9, e102313. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M.; Dixon, L.; Santora, D.; Ibrahim, M.M. Transspinal constant-current long-lasting stimulation: A new method to induce cortical and corticospinal plasticity. J. Neurophysiol. 2015, 114, 1486–1499. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, Y.; Shimoji, K.; Shimizu, H.; Kuribayashi, H.; Fujioka, H. Human spinal cord potentials evoked by different sources of stimulation and conduction velocities along the cord. J. Neurophysiol. 1982, 48, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- De Noordhout, A.M.; Rothwell, J.; Thompson, P.D.; Day, B.L.; Marsden, C.D. Percutaneous electrical stimulation of lumbosacral roots in man. J. Neurol. Neurosurg. Psychiatry 1988, 51, 174–181. [Google Scholar] [CrossRef]
- Danner, S.M.; Krenn, M.; Hofstoetter, U.S.; Toth, A.; Mayr, W.; Minassian, K. Body Position Influences Which Neural Structures Are Recruited by Lumbar Transcutaneous Spinal Cord Stimulation. PLoS ONE 2016, 11, e0147479. [Google Scholar] [CrossRef]
- Minassian, K.; Hofstoetter, U.; Rattay, F. Transcutaneous lumbar posterior root stimulation for motor control studies and modification of motor activity after spinal cord injury. In Restorative Neurology of Spinal Cord Injury; Dimitrijevic, M.R., Byron, A., Vrbova, G., McKay, W.B., Eds.; Oxford University Press: New York, NY, USA, 2011; pp. 226–255. [Google Scholar]
- Hofstoetter, U.S.; Freundl, B.; Binder, H.; Minassian, K. Common neural structures activated by epidural and transcutaneous lumbar spinal cord stimulation: Elicitation of posterior root-muscle reflexes. PLoS ONE 2018, 13, e0192013. [Google Scholar] [CrossRef]
- Lloyd, D.P.C. Monosynaptic reflex response of individual motoneurons as a function of frequency. J. Gen. Physiol. 1957, 40, 435–450. [Google Scholar] [CrossRef]
- Eccles, J.C.; Rall, W. Effects induced in a monosynaptic reflex path by its activation. J. Neurophysiol. 1951, 14, 353–376. [Google Scholar] [CrossRef]
- Rothwell, J.; Day, B.L.; Berardelli, A.; Marsden, C.D. Habituation and conditioning of the human long latency stretch reflex. Exp. Brain Res. 1986, 63, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Sotocinal, S.G.; Sorge, R.E.; Zaloum, A.; Tuttle, A.H.; Martin, L.; Wieskopf, J.S.; Mapplebeck, J.C.; Wei, P.; Zhan, S.; Zhang, S.; et al. The Rat Grimace Scale: A Partially Automated Method for Quantifying Pain in the Laboratory Rat via Facial Expressions. Mol. Pain 2011, 7, 55. [Google Scholar] [CrossRef] [PubMed]
- Sayenko, D.G.; Atkinson, D.A.; Dy, C.J.; Gurley, K.M.; Smith, V.L.; Angeli, C.; Harkema, S.J.; Edgerton, V.R.; Gerasimenko, Y.P. Spinal segment-specific transcutaneous stimulation differentially shapes activation pattern among motor pools in humans. J. Appl. Physiol. 2015, 118, 1364–1374. [Google Scholar] [CrossRef]
- Roy, F.D.; Gibson, G.; Stein, R.B. Effect of percutaneous stimulation at different spinal levels on the activation of sensory and motor roots. Exp. Brain Res. 2012, 223, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Krenn, M.; Hofstoetter, U.S.; Danner, S.M.; Minassian, K.; Mayr, W. Multi-Electrode Array for Transcutaneous Lumbar Posterior Root Stimulation. Artif. Organs 2015, 39, 834–840. [Google Scholar] [CrossRef] [PubMed]
- Knikou, M.; Murray, L. Neural interactions between transspinal evoked potentials and muscle spindle afferents in humans. J. Electromyogr. Kinesiol. 2018, 43, 174–183. [Google Scholar] [CrossRef]
- Sharpe, A.N.; Jackson, A. Upper-limb muscle responses to epidural, subdural and intraspinal stimulation of the cervical spinal cord. J. Neural Eng. 2014, 11, 016005. [Google Scholar] [CrossRef]
- Jackson, A.; Baker, S.N.; Fetz, E.E. Tests for presynaptic modulation of corticospinal terminals from peripheral afferents and pyramidal tract in the macaque. J. Physiol. 2006, 573, 107–120. [Google Scholar] [CrossRef]
- Thomas, P.K.; Sears, T.A.; Gilliatt, R.W. The range of conduction velocity in normal motor nerve fibres to the small muscles of the hand and foot. J. Neurol. Neurosurg. Psychiatry 1959, 22, 175–181. [Google Scholar] [CrossRef]
- Yap, C.B.; Hirota, T. Sciatic nerve motor conduction velocity study. J. Neurol. Neurosurg. Psychiatry 1967, 30, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Buchthal, F.; Rosenfalck, A. Evoked action potentials and conduction velocity in human sensory nerves. Brain Res. 1966, 3, 1–122. [Google Scholar] [CrossRef]
- Huseyinoglu, N.; Ozaydin, I.; Huseyinoglu, U.; Yayla, S.; Aksoy, O. Minimally Invasive Motor Nerve Conduction Study of the Rat Sciatic and Tail Nerves. Kafkas Üniversitesi Veteriner Fakültesi Dergisi 2013, 19, 943–948. [Google Scholar] [CrossRef][Green Version]
- Hofstoetter, U.S.; Danner, S.M.; Minassian, K. Paraspinal magnetic and transcutaneous electrical stimulation. In Encyclopedia of Computational Neuroscience; Jaeger, D., Jung, R., Eds.; Springer: New York, NY, USA, 2014. [Google Scholar]
- Knikou, M. The H-reflex as a probe: Pathways and pitfalls. J. Neurosci. Methods 2008, 171, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cote, M.-P.; Azzam, G.A.; Lemay, M.A.; Zhukareva, V.; Houlé, J.D. Activity-Dependent Increase in Neurotrophic Factors Is Associated with an Enhanced Modulation of Spinal Reflexes after Spinal Cord Injury. J. Neurotrauma 2011, 28, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Cote, M.-P.; Gandhi, S.; Zambrotta, M.; Houlé, J.D. Exercise Modulates Chloride Homeostasis after Spinal Cord Injury. J. Neurosci. 2014, 34, 8976–8987. [Google Scholar] [CrossRef]
- Beverungen, H.; Klaszky, S.C.; Klaszky, M.; Côté, M.-P. Rehabilitation Decreases Spasticity by Restoring Chloride Homeostasis through the Brain-Derived Neurotrophic Factor–KCC2 Pathway after Spinal Cord Injury. J. Neurotrauma 2020, 37, 846–859. [Google Scholar] [CrossRef]
- Ahmed, Z. Trans-spinal direct current stimulation modifies spinal cord excitability through synaptic and axonal mechanisms. Physiol. Rep. 2014, 2, e12157. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R2 | Max | m | S50 | Slope | S-Threshold | S-Max | ||
|---|---|---|---|---|---|---|---|---|
| Rats | L-Pl | 0.958 ± 0.013 | 2.29 ± 1.03 | 1.08 ± 0.21 | 6.03 ± 0.92 | 2.36 ± 0.46 | 4.21 ± 0.55 | 8.94 ± 1.27 |
| L-MG | 0.977 ± 0.003 | 14.62 ± 2.12 | 1.36 ± 0.32 | 5.17 ± 0.39 | 2.00 ± 0.31 | 3.17 ± 0.24 | 7.18 ± 0.66 | |
| L-TA | 0.984 ± 0.004 | 12.76 ± 2.85 | 1.42 ± 0.21 | 4.39 ± 0.48 | 1.59 ± 0.16 | 2.80 ± 0.41 | 5.98 ± 0.59 | |
| R-TA | 0.986 ± 0.003 | 14.94 ± 2.63 | 2.13 ± 0.60 | 4.35 ± 0.60 | 1.45 ± 0.31 | 2.91 ± 0.38 | 5.80 ± 0.87 | |
| Humans | L-RF | 0.923 ± 0.018 | 7.90 ± 1.94 | 0.08 ± 0.03 | 170.84 ± 14.07 | 52.37 ± 14.82 | 118.47 ± 10.88 | 223.21 ± 36.64 |
| L-MG | 0.947 ± 0.013 | 20.17 ± 2.46 | 0.08 ± 0.02 | 175.95 ± 14.07 | 31.56 ± 4.76 | 144.39 ± 11.56 | 207.51 ± 17.53 | |
| L-TA | 0.953 ± 0.005 | 7.16 ± 0.83 | 0.05 ± 0.01 | 170.63 ± 13.08 | 53.66 ± 8.82 | 116.97 ± 9.27 | 224.28 ± 20.29 | |
| R-TA | 0.950 ± 0.010 | 9.68 ± 1.52 | 0.05 ± 0.01 | 177.73 ± 16.29 | 58.01 ± 9.98 | 119.71 ± 11.12 | 235.74 ± 24.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malloy, D.C.; Knikou, M.; Côté, M.-P. Adapting Human-Based Transcutaneous Spinal Cord Stimulation to Develop a Clinically Relevant Animal Model. J. Clin. Med. 2022, 11, 2023. https://doi.org/10.3390/jcm11072023
Malloy DC, Knikou M, Côté M-P. Adapting Human-Based Transcutaneous Spinal Cord Stimulation to Develop a Clinically Relevant Animal Model. Journal of Clinical Medicine. 2022; 11(7):2023. https://doi.org/10.3390/jcm11072023
Chicago/Turabian StyleMalloy, Dillon C., Maria Knikou, and Marie-Pascale Côté. 2022. "Adapting Human-Based Transcutaneous Spinal Cord Stimulation to Develop a Clinically Relevant Animal Model" Journal of Clinical Medicine 11, no. 7: 2023. https://doi.org/10.3390/jcm11072023
APA StyleMalloy, D. C., Knikou, M., & Côté, M.-P. (2022). Adapting Human-Based Transcutaneous Spinal Cord Stimulation to Develop a Clinically Relevant Animal Model. Journal of Clinical Medicine, 11(7), 2023. https://doi.org/10.3390/jcm11072023