
Caveolin-3 and Arrhythmias: Insights into the Molecular Mechanisms


Abstract
1. Introduction
2. Caveolin-3 and Electrical Signal Propagation
2.1. The Sodium Current (INa)
2.2. The K+ Current (Ik)
2.3. The Ca+ Current (ICa)

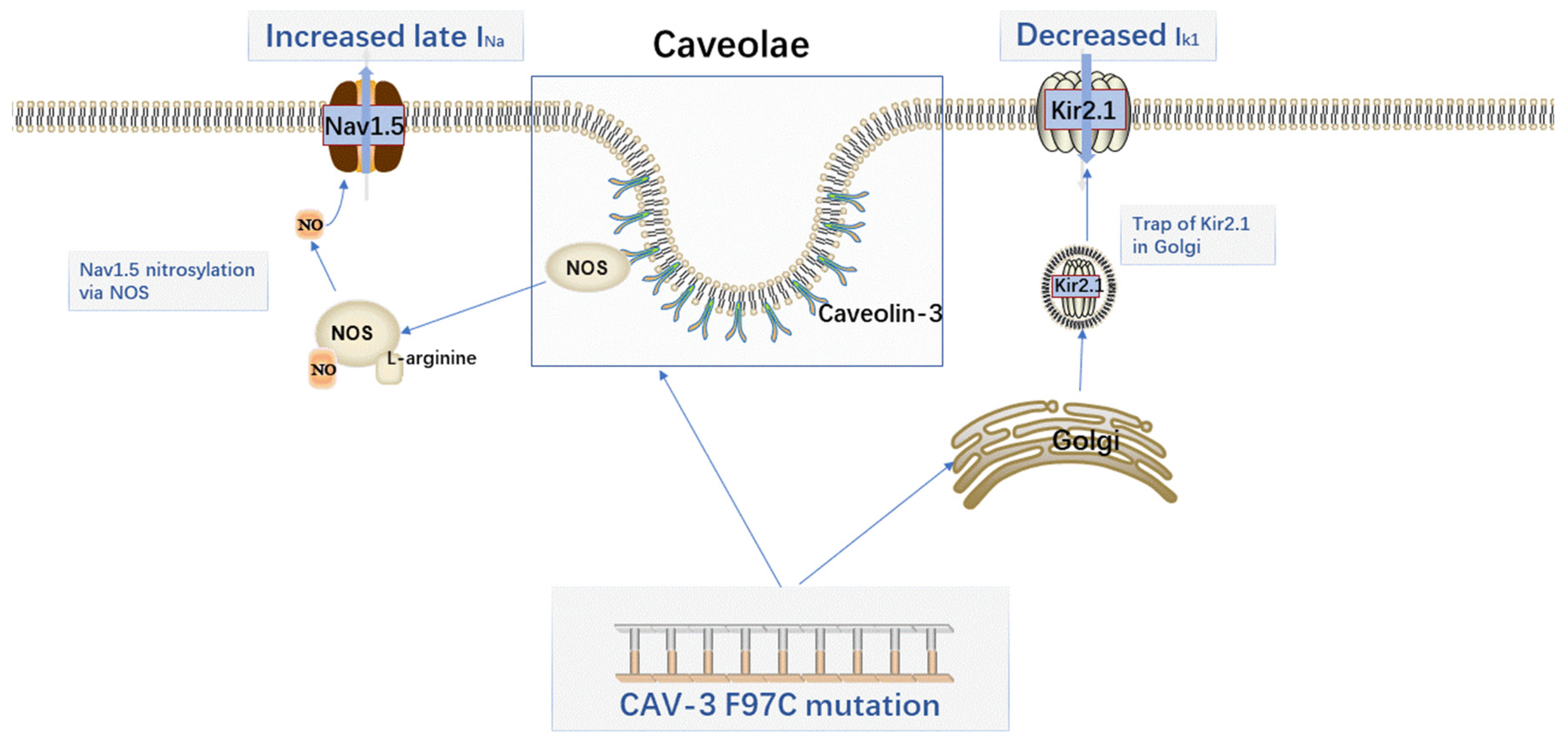{kind=link}
| CAV-3 Expression | Functional Alteration | Arrhythmia Implications | Related Mechanisms | Ref. |
|---|---|---|---|---|
| CAV-3 mutation (V14L, T78M, L79R) | Increased late sodium current | LQT3 | NOS-dependent S-nitrosylation of SCN5A | Cheng. J. et al. [30] |
| Decreased Kir2.x current density | LQT9 | Downstream Ang1R signaling involves the activation of PKC | Tyan. L. et al. [42] | |
| Caveolin-3 overexpression | Reduced diastolic spontaneous Ca2+ waves | Ventricular arrhythmias | Inhibition of RYR2 hyperphosphorylation | Zhang. ZH. et al. [64] |
| CAV-3 mutation (S141R) | Increased HCN4 current density | LQTS | NA | NA |
| Caveolin-3 downregulated expression | Activated ICl, swell | Atrial fibrillation | NA | NA |
2.4. The Hyperpolarization-Activated Cyclic Nucleotide Channel 4 Current (IHCN4)
2.5. The Volume-Activated Cl-Channel Current (ICl,swell)
3. Caveolin-3 and Intercellular Communication
3.1. Caveolin-3 and ConnexIn 43
3.2. Caveolin-3 and Dystrophin
3.3. Caveolin-3 and Adiponectin Receptor
4. Caveolin-3 and Metabolic Perturbation
4.1. Caveolin-3 and Insulin Resistance
4.2. Caveolin-3 and Adiposity
5. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamada, E. The fine structure of the gall bladder epithelium of the mouse. J. Biophys. Biochem. Cytol. 1955, 1, 445–458. [Google Scholar] [CrossRef]
- Rothberg, K.G.; Heuser, J.E.; Donzell, W.C.; Ying, Y.S.; Glenney, J.R.; Anderson, R.G. Caveolin, a protein component of caveolae membrane coats. Cell 1992, 68, 673–682. [Google Scholar] [CrossRef]
- Song, K.S.; Scherer, P.E.; Tang, Z.; Okamoto, T.; Li, S.; Chafel, M.; Chu, C.; Kohtz, D.S.; Lisanti, M.P. Expression of caveolin-3 in skeletal, cardiac, and smooth muscle cells. Caveolin-3 is a component of the sarcolemma and co-fractionates with dystrophin and dystrophin-associated glycoproteins. J. Biol. Chem. 1996, 271, 15160–15165. [Google Scholar] [CrossRef]
- Balijepalli, R.C.; Kamp, T.J. Caveolae, ion channels and cardiac arrhythmias. Prog. Biophys. Mol. Biol. 2008, 98, 149–160. [Google Scholar] [CrossRef]
- Kim, J.H.; Peng, D.; Schlebach, J.P.; Hadziselimovic, A.; Sanders, C.R. Modest effects of lipid modifications on the structure of caveolin-3. Biochemistry 2014, 53, 4320–4322. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Schlebach, J.P.; Lu, Z.; Peng, D.; Reasoner, K.C.; Sanders, C.R. A pH-Mediated Topological Switch within the N-Terminal Domain of Human Caveolin-3. Biophys. J. 2016, 110, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.; Ying, Y.; Albanesi, J.; Anderson, R.G. Mechanism of caveolin filament assembly. Proc. Natl. Acad. Sci. USA 2002, 99, 11193–11198. [Google Scholar] [CrossRef] [PubMed]
- Koga, A.; Oka, N.; Kikuchi, T.; Miyazaki, H.; Kato, S.; Imaizumi, T. Adenovirus-mediated overexpression of caveolin-3 inhibits rat cardiomyocyte hypertrophy. Hypertension 2003, 42, 213–219. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tsutsumi, Y.M.; Horikawa, Y.T.; Jennings, M.M.; Kidd, M.W.; Niesman, I.R.; Yokoyama, U.; Head, B.P.; Hagiwara, Y.; Ishikawa, Y.; Miyanohara, A.; et al. Cardiac-specific overexpression of caveolin-3 induces endogenous cardiac protection by mimicking ischemic preconditioning. Circulation 2008, 118, 1979–1988. [Google Scholar] [CrossRef]
- Parton, R.G. Caveolae: Structure, Function, and Relationship to Disease. Annu. Rev. Cell Dev. Biol. 2018, 34, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Israeli-Rosenberg, S.; Chen, C.; Li, R.; Deussen, D.N.; Niesman, I.R.; Okada, H.; Patel, H.H.; Roth, D.M.; Ross, R.S. Caveolin modulates integrin function and mechanical activation in the cardiomyocyte. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2015, 29, 374–384. [Google Scholar] [CrossRef]
- Yarbrough, T.L.; Lu, T.; Lee, H.C.; Shibata, E.F. Localization of cardiac sodium channels in caveolin-rich membrane domains: Regulation of sodium current amplitude. Circ. Res. 2002, 90, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Martens, J.R.; Sakamoto, N.; Sullivan, S.A.; Grobaski, T.D.; Tamkun, M.M. Isoform-specific localization of voltage-gated K+ channels to distinct lipid raft populations. Targeting of Kv1.5 to caveolae. J. Biol. Chem. 2001, 276, 8409–8414. [Google Scholar] [CrossRef] [PubMed]
- Bossuyt, J.; Taylor, B.E.; James-Kracke, M.; Hale, C.C. The cardiac sodium-calcium exchanger associates with caveolin-3. Ann. N. Y. Acad. Sci. 2002, 976, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Balijepalli, R.C.; Foell, J.D.; Hall, D.D.; Hell, J.W.; Kamp, T.J. Localization of cardiac L-type Ca(2+) channels to a caveolar macromolecular signaling complex is required for beta(2)-adrenergic regulation. Proc. Natl. Acad. Sci. USA 2006, 103, 7500–7505. [Google Scholar] [CrossRef] [PubMed]
- Vatta, M.; Ackerman, M.J.; Ye, B.; Makielski, J.C.; Ughanze, E.E.; Taylor, E.W.; Tester, D.J.; Balijepalli, R.C.; Foell, J.D.; Li, Z.; et al. Mutant caveolin-3 induces persistent late sodium current and is associated with long-QT syndrome. Circulation 2006, 114, 2104–2112. [Google Scholar] [CrossRef]
- Cronk, L.B.; Ye, B.; Kaku, T.; Tester, D.J.; Vatta, M.; Makielski, J.C.; Ackerman, M.J. Novel mechanism for sudden infant death syndrome: Persistent late sodium current secondary to mutations in caveolin-3. Hear. Rhythm 2007, 4, 161–166. [Google Scholar] [CrossRef]
- .Catteruccia, M.; Sanna, T.; Santorelli, F.M.; Tessa, A.; Di Giacopo, R.; Sauchelli, D.; Verbo, A.; Monaco, M.L.; Servidei, S. Rippling muscle disease and cardiomyopathy associated with a mutation in the CAV3 gene. Neuromuscul. Disord. 2009, 19, 779–783. [Google Scholar] [CrossRef]
- Traverso, M.; Gazzerro, E.; Assereto, S.; Sotgia, F.; Biancheri, R.; Stringara, S.; Giberti, L.; Pedemonte, M.; Wang, X.; Scapolan, S.; et al. Caveolin-3 T78M and T78K missense mutations lead to different phenotypes in vivo and in vitro. Lab. Investig. 2008, 88, 275–283. [Google Scholar] [CrossRef][Green Version]
- Hayashi, T.; Arimura, T.; Ueda, K.; Shibata, H.; Hohda, S.; Takahashi, M.; Hori, H.; Koga, Y.; Oka, N.; Imaizumi, T.; et al. Identification and functional analysis of a caveolin-3 mutation associated with familial hypertrophic cardiomyopathy. Biochem. Biophys. Res. Commun. 2004, 313, 178–184. [Google Scholar] [CrossRef]
- Ohsawa, Y.; Tokoet, H.; Katsura, M.; Morimoto, K.; Yamada, H.; Ichikawa, Y.; Murakami, T.; Ohkuma, S.; Komuro, I.; Sunada, Y. Overexpression of P104L mutant caveolin-3 in mice develops hypertrophic cardiomyopathy with enhanced contractility in association with increased endothelial nitric oxide synthase activity. Hum. Mol. Genet. 2004, 13, 151–157. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bruno, G.; Puoti, G.; Oliva, M.; Colavito, D.; Allegorico, L.; Napolitano, F.; Sampaoloet, S. A novel missense mutation in CAV3 gene in an Italian family with persistent hyperCKemia, myalgia and hypercholesterolemia: Double-trouble. Clin. Neurol. Neurosurg. 2020, 191. [Google Scholar] [CrossRef] [PubMed]
- Gal, D.B.; Wojciak, J.; Perera, J.; Tanel, R.E.; Patel, A.R. Atrial standstill in a pediatric patient with associated caveolin-3 mutation. Hear. Case Rep. 2017, 3, 513–516. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Head, B.P.; Patel, H.H.; Roth, D.M.; Lai, N.C.; Niesman, I.R.; Farquhar, M.G.; Insel, P.A. G-protein-coupled receptor signaling components localize in both sarcolemmal and intracellular caveolin-3-associated microdomains in adult cardiac myocytes. J. Biol. Chem. 2005, 280, 31036–31044. [Google Scholar] [CrossRef] [PubMed]
- Lariccia, V.; Nasti, A.A.; Alessandrini, F.; Pesaresi, M.; Gratteri, S.; Tagliabracci, A.; Amoroso, S. Identification and functional analysis of a new putative caveolin-3 variant found in a patient with sudden unexplained death. J. Biomed. Sci. 2014, 21, 58. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, M.J.; Siu, B.L.; Sturner, W.Q.; Tester, D.J.; Valdivia, C.R.; Makielski, J.C.; Towbin, J.A. Postmortem molecular analysis of SCN5A defects in sudden infant death syndrome. JAMA 2001, 286, 2264–2269. [Google Scholar] [CrossRef] [PubMed]
- Ahern, G.P.; Hsu, S.F.; Klyachko, V.A.; Jackson, M.B. Induction of persistent sodium current by exogenous and endogenous nitric oxide. J. Biol. Chem. 2000, 275, 28810–28815. [Google Scholar] [CrossRef] [PubMed]
- Venema, V.J.; Ju, H.; Zou, R.; Venema, R.C. Interaction of neuronal nitric-oxide synthase with caveolin-3 in skeletal muscle. Identification of a novel caveolin scaffolding/inhibitory domain. J. Biol. Chem. 1997, 272, 28187–28190. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.H.; Pundi, K.N.; Van Norstrand, D.W.; Valdivia, C.R.; Tester, D.J.; Medeiros-Domingo, A.; Makielski, J.C.; Ackerman, M.J. Sudden infant death syndrome-associated mutations in the sodium channel beta subunits. Heart Rhythm 2010, 7, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Valdivia, C.R.; Vaidyanathan, R.; Balijepalli, R.C.; Ackerman, M.J.; Makielski, J.C. Caveolin-3 suppresses late sodium current by inhibiting nNOS-dependent S-nitrosylation of SCN5A. J. Mol. Cell. Cardiol. 2013, 61, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Tyan, L.; Foell, J.D.; Vincent, K.P.; Woon, M.T.; Mesquitta, W.T.; Lang, D.; Best, J.M.; Ackerman, M.J.; McCulloch, A.D.; Glukhov, A.V.; et al. Long QT syndrome caveolin-3 mutations differentially modulate K(v) 4 and Ca(v) 1.2 channels to contribute to action potential prolongation. J. Physiol. 2019, 597, 1531–1551. [Google Scholar] [CrossRef] [PubMed]
- Palygin, O.A.; Pettus, J.M.; Shibata, E.F. Regulation of caveolar cardiac sodium current by a single Gsalpha histidine residue. Am. J. Physiology. Heart Circ. Physiol. 2008, 294, H1693–H1699. [Google Scholar] [CrossRef] [PubMed]
- Miake, J.; Marbán, E.; Nuss, H.B. Functional role of inward rectifier current in heart probed by Kir2.1 overexpression and dominant-negative suppression. J. Clin. Investig. 2003, 111, 1529–1536. [Google Scholar] [CrossRef]
- Liu, G.X.; Derst, C.; Schlichthörl, G.; Heinen, S.; Seebohm, G.; Brüggemann, A.; Kummer, W.; Veh, R.W.; Daut, J.; Preisig-Müller, R. Comparison of cloned Kir2 channels with native inward rectifier K+ channels from guinea-pig cardiomyocytes. J. Physiol. 2001, 532, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Anumonwo, J.M.; Lopatin, A.N. Cardiac strong inward rectifier potassium channels. J. Mol. Cell. Cardiol. 2010, 48, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, R.; Van Ert, H.; Haq, K.T.; Morotti, S.; Esch, S.; McCune, E.C.; Grandi, E.; Eckhardt, L.L. Inward Rectifier Potassium Channels (Kir2.x) and Caveolin-3 Domain-Specific Interaction: Implications for Purkinje Cell-Dependent Ventricular Arrhythmias. Circ. Arrhythmia Electrophysiol. 2018, 11, e005800. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, R.; Vega, A.L.; Song, C.; Zhou, Q.; Tan, B.H.; Berger, S.; Makielski, J.C.; Eckhardt, L.L. The interaction of caveolin 3 protein with the potassium inward rectifier channel Kir2.1: Physiology and pathology related to long qt syndrome 9 (LQT9). J. Biol. Chem. 2013, 288, 17472–17480. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Yu, T.; Yang, H.; Peng, Z. Exhaustive exercise-induced cardiac conduction system injury and changes of cTnT and Cx43. Int. J. Sports Med. 2015, 36, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, R.; Reilly, L.; Eckhardt, L.L. Caveolin-3 Microdomain: Arrhythmia Implications for Potassium Inward Rectifier and Cardiac Sodium Channel. Front. Physiol. 2018, 9, 1548. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Balbuena, D.; Guerrero-Serna, G.; Valdivia, C.R.; Caballero, R.; Diez-Guerra, F.J.; Jiménez-Vázquez, E.N.; Ramírez, R.J.; Monteiro da Rocha, A.; Herron, T.J.; Campbell, K.F.; et al. Cardiac Kir2.1 and Na(V)1.5 Channels Traffic Together to the Sarcolemma to Control Excitability. Circ. Res. 2018, 122, 1501–1516. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Dinh Cat, A.; Touyz, R.M. A new look at the renin-angiotensin system--focusing on the vascular system. Peptides 2011, 32, 2141–2150. [Google Scholar] [CrossRef]
- Tyan, L.; Turner, D.; Komp, K.R.; Medvedev, R.Y.; Lim, E.; Glukhov, A.V. Caveolin-3 is required for regulation of transient outward potassium current by angiotensin II in mouse atrial myocytes. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H787–H797. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.B.; Lokuta, A.J. Angiotensin II signal transduction pathways in the cardiovascular system. Trends Cardiovasc. Med. 1994, 4, 110–116. [Google Scholar] [CrossRef]
- Scholz, E.P.; Welke, F.; Joss, N.; Seyler, C.; Zhang, W.; Scherer, D.; Völkers, M.; Bloehs, R.; Thomas, D.; Katus, H.A.; et al. Central role of PKCα in isoenzyme-selective regulation of cardiac transient outward current Ito and Kv4.3 channels. J. Mol. Cell. Cardiol. 2011, 51, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Jansen, H.J.; Mackasey, M.; Moghtadaei, M.; Belke, D.D.; Egom, E.E.; Tuomi, J.M.; Rafferty, S.A.; Kirkby, A.W.; Rose, R.A. Distinct patterns of atrial electrical and structural remodeling in angiotensin II mediated atrial fibrillation. J. Mol. Cell. Cardiol. 2018, 124, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Kawai, M.; Hussain, M.; Orchard, C.H. Excitation-contraction coupling in rat ventricular myocytes after formamide-induced detubulation. Am. J. Physiol. 1999, 277, H603–H609. [Google Scholar] [CrossRef]
- Shan, J.; Xie, W.; Betzenhauser, M.; Reiken, S.; Chen, B.X.; Wronska, A.; Marks, A.R. Calcium leak through ryanodine receptors leads to atrial fibrillation in 3 mouse models of catecholaminergic polymorphic ventricular tachycardia. Circ. Res. 2012, 111, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Nikolaev, V.O.; Moshkov, A.; Lyon, A.R.; Miragoli, M.; Novak, P.; Paur, H.; Lohse, M.J.; Korchev, Y.E.; Harding, S.E.; Gorelik, J. Beta2-adrenergic receptor redistribution in heart failure changes cAMP compartmentation. Science 2010, 327, 1653–1657. [Google Scholar] [CrossRef] [PubMed]
- Beuckelmann, D.J.; Näbauer, M.; Erdmann, E. Characteristics of calcium-current in isolated human ventricular myocytes from patients with terminal heart failure. J. Mol. Cell. Cardiol. 1991, 23, 929–937. [Google Scholar] [CrossRef]
- Ouadid, H.; Séguin, J.; Richard, S.; Chaptal, P.A.; Nargeot, J. Properties and Modulation of Ca channels in adult human atrial cells. J. Mol. Cell. Cardiol. 1991, 23, 41–54. [Google Scholar] [CrossRef]
- Li, G.R.; Nattel, S. Properties of human atrial ICa at physiological temperatures and relevance to action potential. Am. J. Physiol. 1997, 272, H227–H235. [Google Scholar] [CrossRef] [PubMed]
- Markandeya, Y.S.; Fahey, J.M.; Pluteanu, F.; Cribbs, L.L.; Balijepalli, R.C. Caveolin-3 regulates protein kinase A modulation of the Ca(V)3.2 (alpha1H) T-type Ca2+ channels. J. Biol. Chem. 2011, 286, 2433–2444. [Google Scholar] [CrossRef] [PubMed]
- Markandeya, Y.S.; Phelan, L.J.; Woon, M.T.; Keefe, A.M.; Reynolds, C.R.; August, B.K.; Hacker, T.A.; Roth, D.M.; Patel, H.H.; Balijepalli, R.C. Caveolin-3 Overexpression Attenuates Cardiac Hypertrophy via Inhibition of T-type Ca2+ Current Modulated by Protein Kinase Cα in Cardiomyocytes. J. Biol. Chem. 2015, 290, 22085–22100. [Google Scholar] [CrossRef] [PubMed]
- Chemin, J.; Taiakina, V.; Monteil, A.; Piazza, M.; Guan, W.; Stephens, R.F.; Kitmitto, A.; Pang, Z.P.; Dolphin, A.C.; Perez-Reyes, E.; et al. Calmodulin regulates Ca(v)3 T-type channels at their gating brake. J. Biol. Chem. 2017, 292, 20010–20031. [Google Scholar] [CrossRef]
- Parton, R.G.; Way, M.; Zorzi, N.; Stang, E. Caveolin-3 associates with developing T-tubules during muscle differentiation. J. Cell Biol. 1997, 136, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Bryant, S.; Kimura, T.E.; Kong, C.H.; Watson, J.J.; Chase, A.; Suleiman, M.S.; James, A.F.; Orchard, C.H. Stimulation of ICa by basal PKA activity is facilitated by Caveolin-3 in cardiac ventricular myocytes. J. Mol. Cell. Cardiol. 2014, 68, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, F.; Engelman, J.A.; Volonte, D.; Zhang, X.L.; Minetti, C.; Li, M.; Hou, H., Jr.; Kneitz, B.; Edelmann, W.; Lisanti, M.P. Caveolin-3 null mice show a loss of caveolae, changes in the microdomain distribution of the dystrophin-glycoprotein complex, and t-tubule abnormalities. J. Biol. Chem. 2001, 276, 21425–21433. [Google Scholar] [CrossRef]
- Bryant, S.M.; Kong, C.H.T.; Watson, J.J.; Gadeberg, H.C.; James, A.F.; Cannell, M.B.; Orchard, C.H. Caveolin 3-dependent loss of t-tubular I(Ca) during hypertrophy and heart failure in mice. Exp. Physiol. 2018, 103, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Bhogal, N.K.; Hasan, A.; Gorelik, J. The Development of Compartmentation of cAMP Signaling in Cardiomyocytes: The Role of T-Tubules and Caveolae Microdomains. J. Cardiovasc. Dev. Dis. 2018, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Asghari, P.; Scriven, D.R.; Hoskins, J.; Fameli, N.; van Breemen, C.; Moore, E.D. The structure and functioning of the couplon in the mammalian cardiomyocyte. Protoplasma 2012, 249 (Suppl. 1), S31–S38. [Google Scholar] [CrossRef] [PubMed]
- Fill, M.; Copello, J.A. Ryanodine receptor calcium release channels. Physiol. Rev. 2002, 82, 893–922. [Google Scholar] [CrossRef]
- Wong, J.; Baddeley, D.; Bushong, E.A.; Yu, Z.; Ellisman, M.H.; Hoshijima, M.; Soeller, C. Nanoscale distribution of ryanodine receptors and caveolin-3 in mouse ventricular myocytes: Dilation of t-tubules near junctions. Biophys. J. 2013, 104, L22-24. [Google Scholar] [CrossRef] [PubMed]
- Song, D.W.; Lee, K.E.; Ryu, J.Y.; Jeon, H.; Kim, D.H. The molecular interaction of heart LIM protein (HLP) with RyR2 and caveolin-3 is essential for Ca(2+)-induced Ca(2+) release in the heart. Biochem. Biophys. Res. Commun. 2015, 463, 975–981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fang, Q.; Du, T.; Chen, G.; Wang, Y.; Wang, D.W. Cardiac-Specific Caveolin-3 Overexpression Prevents Post-Myocardial Infarction Ventricular Arrhythmias by Inhibiting Ryanodine Receptor-2 Hyperphosphorylation. Cardiology 2020, 145, 136–147. [Google Scholar] [CrossRef]
- Castets, F.; Rakitina, T.; Gaillard, S.; Moqrich, A.; Mattei, M.G.; Monneron, A. Zinedin, SG2NA, and striatin are calmodulin-binding, WD repeat proteins principally expressed in the brain. J. Biol. Chem. 2000, 275, 19970–19977. [Google Scholar] [CrossRef] [PubMed]
- Nader, M.; Alotaibi, S.; Alsolme, E.; Khalil, B.; Abu-Zaid, A.; Alsomali, R.; Bakheet, D.; Dzimiri, N. Cardiac striatin interacts with caveolin-3 and calmodulin in a calcium sensitive manner and regulates cardiomyocyte spontaneous contraction rate. Can. J. Physiol. Pharmacol. 2017, 95, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Barbuti, A.; Terragni, B.; Brioschi, C.; DiFrancesco, D. Localization of f-channels to caveolae mediates specific beta2-adrenergic receptor modulation of rate in sinoatrial myocytes. J. Mol. Cell. Cardiol. 2007, 42, 71–78. [Google Scholar] [CrossRef]
- Motloch, L.J.; Larbig, R.; Darabi, T.; Reda, S.; Motloch, K.A.; Wernly, B.; Lichtenauer, M.; Gebing, T.; Schwaiger, A.; Zagidullin, N.; et al. Long-QT syndrome-associated Caveolin-3 mutations differentially regulate the hyperpolarization-activated cyclic nucleotide gated channel 4. Physiol. Int. 2017, 104, 130–138. [Google Scholar] [CrossRef]
- Kuwabara, Y.; Kuwahara, K.; Takano, M.; Kinoshita, H.; Arai, Y.; Yasuno, S.; Nakagawa, Y.; Igata, S.; Usami, S.; Minami, T.; et al. Increased expression of HCN channels in the ventricular myocardium contributes to enhanced arrhythmicity in mouse failing hearts. J. Am. Heart Assoc. 2013, 2, e000150. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Y.; Qu, X.; Chen, L.Z.; Zheng, G.S.; Wu, X.L.; Chen, X.X.; Huang, W.J.; Zhou, H. Potential Roles of Serum Caveolin-3 Levels in Patients with Atrial Fibrillation. Front. Aging Neurosci. 2017, 9, 90. [Google Scholar] [CrossRef]
- Reilly, S.N.; Liu, X.; Carnicer, R.; Recalde, A.; Muszkiewicz, A.; Jayaram, R.; Carena, M.C.; Wijesurendra, R.; Stefanini, M.; Surdo, N.C.; et al. Up-regulation of miR-31 in human atrial fibrillation begets the arrhythmia by depleting dystrophin and neuronal nitric oxide synthase. Sci. Transl. Med. 2016, 8, 340ra374. [Google Scholar] [CrossRef] [PubMed]
- Egorov, Y.V.; Lang, D.; Tyan, L.; Turner, D.; Lim, E.; Piro, Z.D.; Hernandez, J.J.; Lodin, R.; Wang, R.; Schmuck, E.G.; et al. Caveolae-Mediated Activation of Mechanosensitive Chloride Channels in Pulmonary Veins Triggers Atrial Arrhythmogenesis. J. Am. Heart Assoc. 2019, 8, e012748. [Google Scholar] [CrossRef] [PubMed]
- Andelova, K.; Egan Benova, T.; Szeiffova Bacova, B.; Sykora, M.; Prado, N.J.; Diez, E.R.; Hlivak, P.; Tribulova, N. Cardiac Connexin-43 Hemichannels and Pannexin1 Channels: Provocative Antiarrhythmic Targets. Int. J. Mol. Sci. 2020, 22, 260. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Yan, X.; Yang, Y.; Chen, M.; Wu, L.; Gou, Z.; Sun, Z.; Talabieke, S.; Zheng, Y.; Luo, D. Connexin 43 dephosphorylation contributes to arrhythmias and cardiomyocyte apoptosis in ischemia/reperfusion hearts. Basic Res. Cardiol. 2019, 114, 40. [Google Scholar] [CrossRef]
- Saffitz, J.E. Arrhythmogenic cardiomyopathy and abnormalities of cell-to-cell coupling. Heart Rhythm 2009, 6, S62–S65. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Shimura, D.; Baum, R.; Hernandez, D.M.; Agvanian, S.; Nagaoka, Y.; Katsumata, M.; Lampe, P.D.; Kleber, A.G.; Hong, T.; et al. Auxiliary trafficking subunit GJA1-20k protects connexin-43 from degradation and limits ventricular arrhythmias. J. Clin. Investig. 2020, 130, 4858–4870. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xu, J.; Hu, W.; Yu, D.; Bai, X. Effects of Pinocembrin Pretreatment on Connexin 43 (Cx43) Protein Expression After Rat Myocardial Ischemia-Reperfusion and Cardiac Arrhythmia. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 5008–5014. [Google Scholar] [CrossRef]
- Rugowska, A.; Wiernicki, B.; Maczewski, M.; Mackiewicz, U.; Chojnacka, K.; Bednarek-Rajewska, K.; Kluk, A.; Majewski, P.; Kolanowski, T.; Malcher, A.; et al. Human skeletal muscle-derived stem/progenitor cells modified with connexin-43 prevent arrhythmia in rat post-infarction hearts and influence gene expression in the myocardium. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2019, 70. [Google Scholar] [CrossRef]
- Yang, K.C.; Rutledge, C.A.; Mao, M.; Bakhshi, F.R.; Xie, A.; Liu, H.; Bonini, M.G.; Patel, H.H.; Minshall, R.D.; Dudley, S.C., Jr. Caveolin-1 modulates cardiac gap junction homeostasis and arrhythmogenecity by regulating cSrc tyrosine kinase. Circ. Arrhythmia Electrophysiol. 2014, 7, 701–710. [Google Scholar] [CrossRef]
- Liu, L.; Li, Y.; Lin, J.; Liang, Q.; Sheng, X.; Wu, J.; Huang, R.; Liu, S.; Li, Y. Connexin43 interacts with Caveolin-3 in the heart. Mol. Biol. Rep. 2010, 37, 1685–1691. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.S.; Mendonça, M.C.; Rocha, T.; Kalapothakis, E.; da Cruz-Höfling, M.A. Are Synchronized Changes in Connexin-43 and Caveolin-3 a Bystander Effect in a Phoneutria nigriventer Venom Model of Blood-Brain Barrier Breakdown? J. Mol. Neurosci. MN 2016, 59, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.K.; Wang, S.M.; Chen, Y.L.; Wang, H.S.; Wu, J.C. Lipopolysaccharide-induced inhibition of connexin43 gap junction communication in astrocytes is mediated by downregulation of caveolin-3. Int. J. Biochem. Cell Biol. 2010, 42, 762–770. [Google Scholar] [CrossRef]
- Valera, I.C.; Wacker, A.L.; Hwang, H.S.; Holmes, C.; Laitano, O.; Landstrom, A.P.; Parvatiyar, M.S. Essential roles of the dystrophin-glycoprotein complex in different cardiac pathologies. Adv. Med. Sci. 2021, 66, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Sinagra, G.; Dal Ferro, M.; Gigli, M. The heart of dystrophinopathies. Eur. J. Heart Fail. 2021, 23, 1287–1289. [Google Scholar] [CrossRef]
- Kamdar, F.; Garry, D.J. Dystrophin-Deficient Cardiomyopathy. J. Am. Coll. Cardiol. 2016, 67, 2533–2546. [Google Scholar] [CrossRef] [PubMed]
- Doyle, D.D.; Goings, G.; Upshaw-Earley, J.; Ambler, S.K.; Mondul, A.; Palfrey, H.C.; Page, E. Dystrophin associates with caveolae of rat cardiac myocytes: Relationship to dystroglycan. Circ. Res. 2000, 87, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Aravamudan, B.; Volonte, D.; Ramani, R.; Gursoy, E.; Lisanti, M.P.; London, B.; Galbiati, F. Transgenic overexpression of caveolin-3 in the heart induces a cardiomyopathic phenotype. Hum. Mol. Genet. 2003, 12, 2777–2788. [Google Scholar] [CrossRef][Green Version]
- Sotgia, F.; Lee, J.K.; Das, K.; Bedford, M.; Petrucci, T.C.; Macioce, P.; Sargiacomo, M.; Bricarelli, F.D.; Minetti, C.; Sudol, M.; et al. Caveolin-3 directly interacts with the C-terminal tail of beta -dystroglycan. Identification of a central WW-like domain within caveolin family members. J. Biol. Chem. 2000, 275, 38048–38058. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. Compr. Physiol. 2018, 8, 1031–1063. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, X.; Jasmin, J.F.; Lau, W.B.; Li, R.; Yuan, Y.; Yi, W.; Chuprun, K.; Lisanti, M.P.; Koch, W.J.; et al. Essential role of caveolin-3 in adiponectin signalsome formation and adiponectin cardioprotection. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Yi, W.; Sun, Y.; Gao, E.; Wei, X.; Lau, W.B.; Zheng, Q.; Wang, Y.; Yuan, Y.; Wang, X.; Tao, L.; et al. Reduced cardioprotective action of adiponectin in high-fat diet-induced type II diabetic mice and its underlying mechanisms. Antioxid. Redox Signal. 2011, 15, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Li, H.; Xu, J.; Liu, Y.; Gao, X.; Wang, J.; Ng, K.F.; Lau, W.B.; Ma, X.L.; Rodrigues, B.; et al. Hyperglycemia-induced protein kinase C β2 activation induces diastolic cardiac dysfunction in diabetic rats by impairing caveolin-3 expression and Akt/eNOS signaling. Diabetes 2013, 62, 2318–2328. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Zhou, L.J.; Mu, P.W.; Liu, S.P.; Chen, S.J.; Fu, X.D.; Wang, T.H. Caveolin-3 is involved in the protection of resveratrol against high-fat-diet-induced insulin resistance by promoting GLUT4 translocation to the plasma membrane in skeletal muscle of ovariectomized rats. J. Nutr. Biochem. 2012, 23, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yao, W.; Liu, Z.; Xu, A.; Huang, Y.; Ma, X.L.; Irwin, M.G.; Xia, Z. Hyperglycemia Abrogates Ischemic Postconditioning Cardioprotection by Impairing AdipoR1/Caveolin-3/STAT3 Signaling in Diabetic Rats. Diabetes 2016, 65, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Hegyi, B.; Bers, D.M.; Bossuyt, J. CaMKII signaling in heart diseases: Emerging role in diabetic cardiomyopathy. J. Mol. Cell. Cardiol. 2019, 127, 246–259. [Google Scholar] [CrossRef]
- Lau, D.H.; Nattel, S.; Kalman, J.M.; Sanders, P. Modifiable Risk Factors and Atrial Fibrillation. Circulation 2017, 136, 583–596. [Google Scholar] [CrossRef]
- Huang, Y.; Deng, Y.; Shang, L.; Yang, L.; Huang, J.; Ma, J.; Liao, X.; Zhou, H.; Xian, J.; Liang, G.; et al. Effect of type 2 diabetes mellitus caveolin-3 K15N mutation on glycometabolism. Exp. Ther. Med. 2019, 18, 2531–2539. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Chen, T.; Xian, J.; Deng, Y.; Huang, Y.; Zhao, Q.; Liang, G.; Liang, Z.; Lian, F.; Wei, H.; et al. The caveolin-3 P104L mutation in LGMD-1C patients inhibits non-insulin-stimulated glucose metabolism and growth but promotes myocyte proliferation. Cell Biol. Int. 2019, 43, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, M.; Wu, T.; Xu, M.; Cai, H.; Zhang, Z. Effects of D-Pinitol on Insulin Resistance through the PI3K/Akt Signaling Pathway in Type 2 Diabetes Mellitus Rats. J. Agric. Food Chem. 2015, 63, 6019–6026. [Google Scholar] [CrossRef]
- Yang, M.; Ren, Y.; Lin, Z.; Tang, C.; Jia, Y.; Lai, Y.; Zhou, T.; Wu, S.; Liu, H.; Yang, G.; et al. Krüppel-like factor 14 increases insulin sensitivity through activation of PI3K/Akt signal pathway. Cell. Signal. 2015, 27, 2201–2208. [Google Scholar] [CrossRef]
- Mu, P.; Tan, Z.; Cui, Y.; Liu, H.; Xu, X.; Huang, Q.; Zeng, L.; Wang, T. 17β-Estradiol attenuates diet-induced insulin resistance and glucose intolerance through up-regulation of caveolin-3. Ir. J. Med. Sci. 2011, 180, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Su, W.; Xia, Z.Y.; Wang, Y.; Zhou, L.; Qiao, S.; Zhao, B.; Xia, Z.; Irwin, M.G. Hyperglycemia-Induced Oxidative Stress Abrogates Remifentanil Preconditioning-Mediated Cardioprotection in Diabetic Rats by Impairing Caveolin-3-Modulated PI3K/Akt and JAK2/STAT3 Signaling. Oxidative Med. Cell. Longev. 2019, 2019, 9836302. [Google Scholar] [CrossRef] [PubMed]
- Margolis, K.L.; Bonds, D.E.; Rodabough, R.J.; Tinker, L.; Phillips, L.S.; Allen, C.; Bassford, T.; Burke, G.; Torrens, J.; Howard, B.V. Effect of oestrogen plus progestin on the incidence of diabetes in postmenopausal women: Results from the Women’s Health Initiative Hormone Trial. Diabetologia 2004, 47, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Capozza, F.; Combs, T.P.; Cohen, A.W.; Cho, Y.R.; Park, S.Y.; Schubert, W.; Williams, T.M.; Brasaemle, D.L.; Jelicks, L.A.; Scherer, P.E.; et al. Caveolin-3 knockout mice show increased adiposity and whole body insulin resistance, with ligand-induced insulin receptor instability in skeletal muscle. Am. J. Physiology. Cell Physiol. 2005, 288, C1317–C1331. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.K.; Chen, Y.C.; Chen, J.H.; Chen, S.A.; Chen, Y.J. Adipocytes modulate the electrophysiology of atrial myocytes: Implications in obesity-induced atrial fibrillation. Basic Res. Cardiol. 2012, 107, 293. [Google Scholar] [CrossRef] [PubMed]
- Omran, J.; Bostick, B.P.; Chan, A.K.; Alpert, M.A. Obesity and Ventricular Repolarization: A Comprehensive Review. Prog. Cardiovasc. Dis. 2018, 61, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ruiz, A.; Milagro, F.I.; Campión, J.; Martínez, J.A.; de Miguel, C. Caveolin expression and activation in retroperitoneal and subcutaneous adipocytes: Influence of a high-fat diet. J. Cell. Physiol. 2010, 225, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ruiz, A.; Milagro, F.I.; Campión, J.; Martínez, J.A.; de Miguel, C. High-fat diet feeding alters metabolic response to fasting/non fasting conditions. Effect on caveolin expression and insulin signalling. Lipids Health Dis. 2011, 10, 55. [Google Scholar] [CrossRef]
- Feillet-Coudray, C.; Sutra, T.; Fouret, G.; Ramos, J.; Wrutniak-Cabello, C.; Cabello, G.; Cristol, J.P.; Coudray, C. Oxidative stress in rats fed a high-fat high-sucrose diet and preventive effect of polyphenols: Involvement of mitochondrial and NAD(P)H oxidase systems. Free Radic. Biol. Med. 2009, 46, 624–632. [Google Scholar] [CrossRef] [PubMed]

| Phenotype | CAV3 Mutation | Serum CK Concentrations | Ref. |
|---|---|---|---|
| LQTS | p.A85T | NA | Vatta, M. et al. [16] |
| Sudden infant death syndrome | p.V14 L | NA | Cronk, L.B. et al. [17] |
| Dilated cardiomyopathy | p.A46V | High | Catteruccia, M. et al. [18] |
| Dilated cardiomyopathy | p.T78M | High | Traverso, M. et al. [19] |
| Hypertrophic cardiomyopathy | p.T63S | Normal | Hayashi, T. et al. [20] |
| Hypertrophic cardiomyopathy | P104L | NA | Ohsawa, Y. et al. [21] |
| Hypercholesterolemia | p. Val44Met | High | Bruno, G. et al. [22] |
| Atrial standstill | p. Leu84Pro | NA | Gal, D. B. et al. [23] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Qiu, J.; Wang, Y.; Bai, Y.; Chen, G. Caveolin-3 and Arrhythmias: Insights into the Molecular Mechanisms. J. Clin. Med. 2022, 11, 1595. https://doi.org/10.3390/jcm11061595
He M, Qiu J, Wang Y, Bai Y, Chen G. Caveolin-3 and Arrhythmias: Insights into the Molecular Mechanisms. Journal of Clinical Medicine. 2022; 11(6):1595. https://doi.org/10.3390/jcm11061595
Chicago/Turabian StyleHe, Miaomiao, Jie Qiu, Yan Wang, Yang Bai, and Guangzhi Chen. 2022. "Caveolin-3 and Arrhythmias: Insights into the Molecular Mechanisms" Journal of Clinical Medicine 11, no. 6: 1595. https://doi.org/10.3390/jcm11061595
APA StyleHe, M., Qiu, J., Wang, Y., Bai, Y., & Chen, G. (2022). Caveolin-3 and Arrhythmias: Insights into the Molecular Mechanisms. Journal of Clinical Medicine, 11(6), 1595. https://doi.org/10.3390/jcm11061595