
Loss of E-Cadherin Staining Continuity in the Trophoblastic Basal Membrane Correlates with Increased Resistance in Uterine Arteries and Proteinuria in Patients with Pregnancy-Induced Hypertension

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. Ultrasound
2.3. Immunohistochemistry
2.4. Statistics
2.5. Ethics
3. Results
3.1. Basic Characteristics of the Study and Control Groups
3.2. Incidence of an Early Diastolic Notch and Proteinuria in the Study and Control Groups
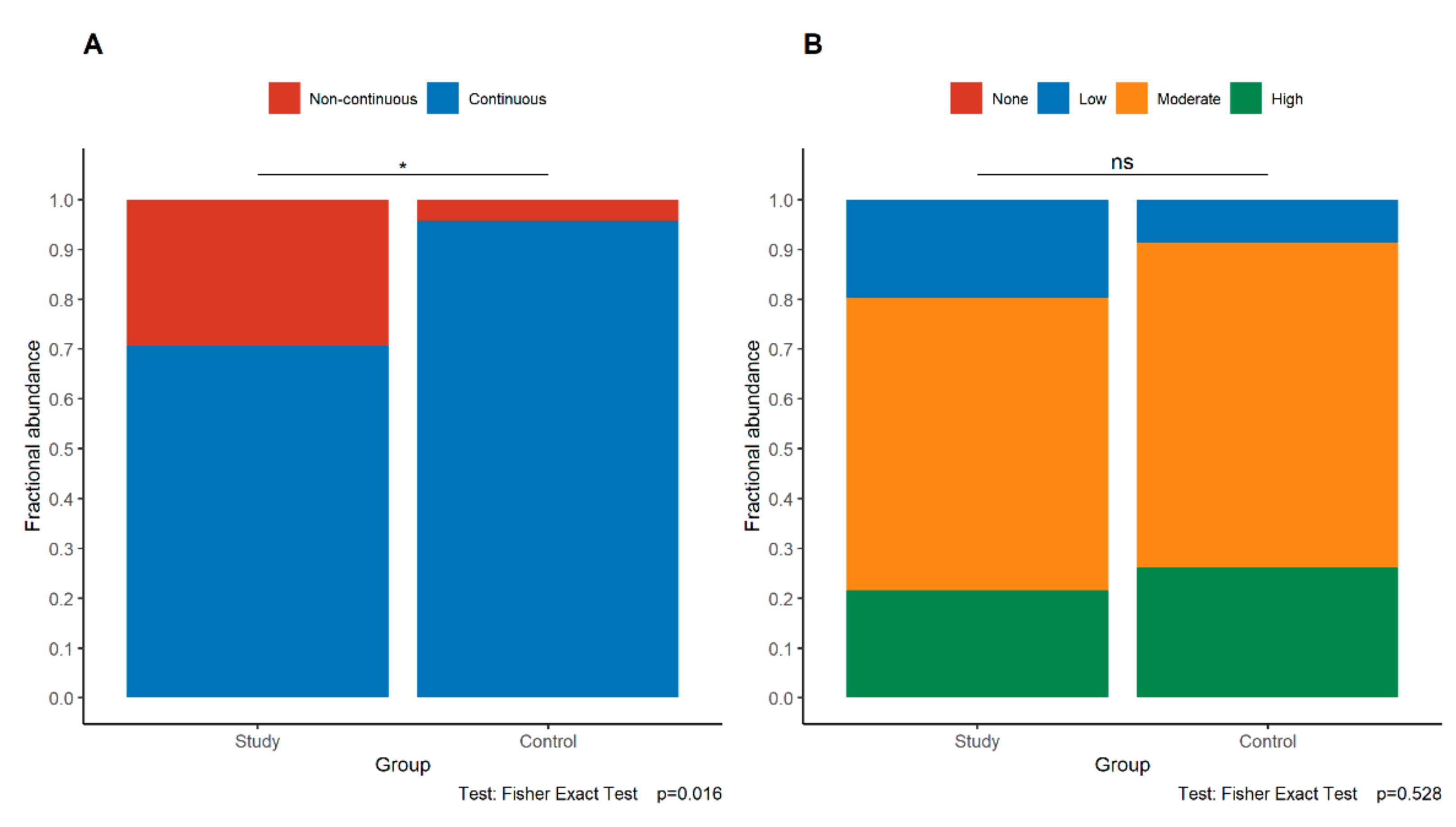
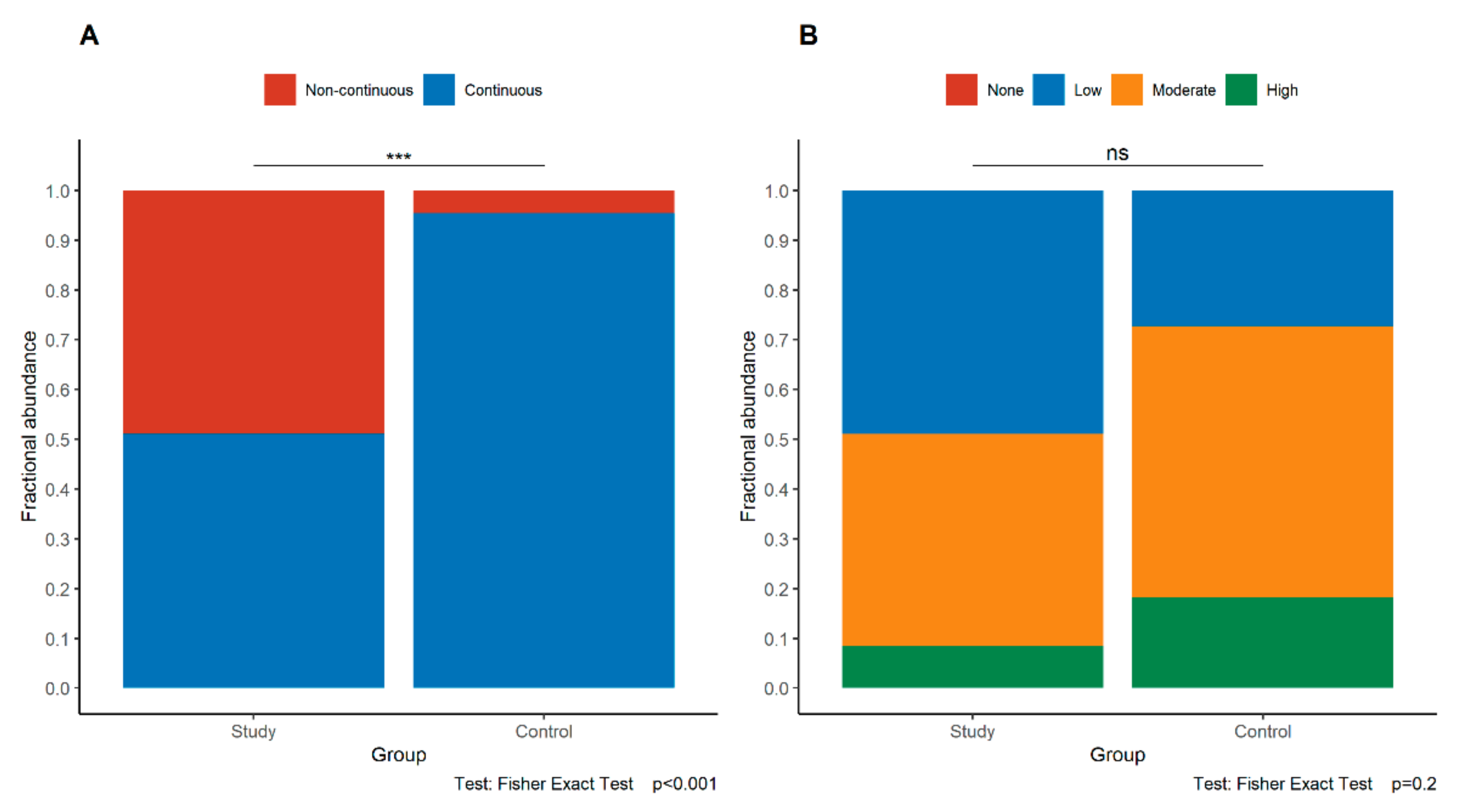
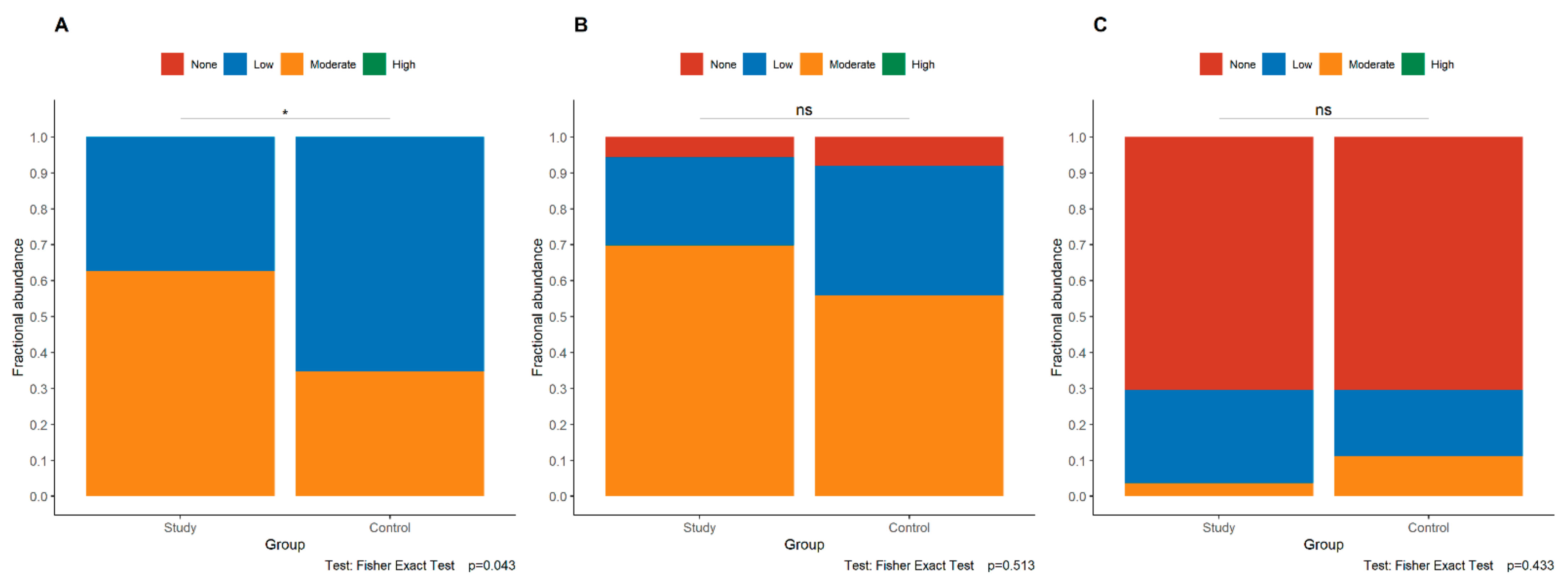
3.3. Expression of E-Cadherin in the Study and Control Groups
3.4. Expression of p120 in the Study and Control Groups
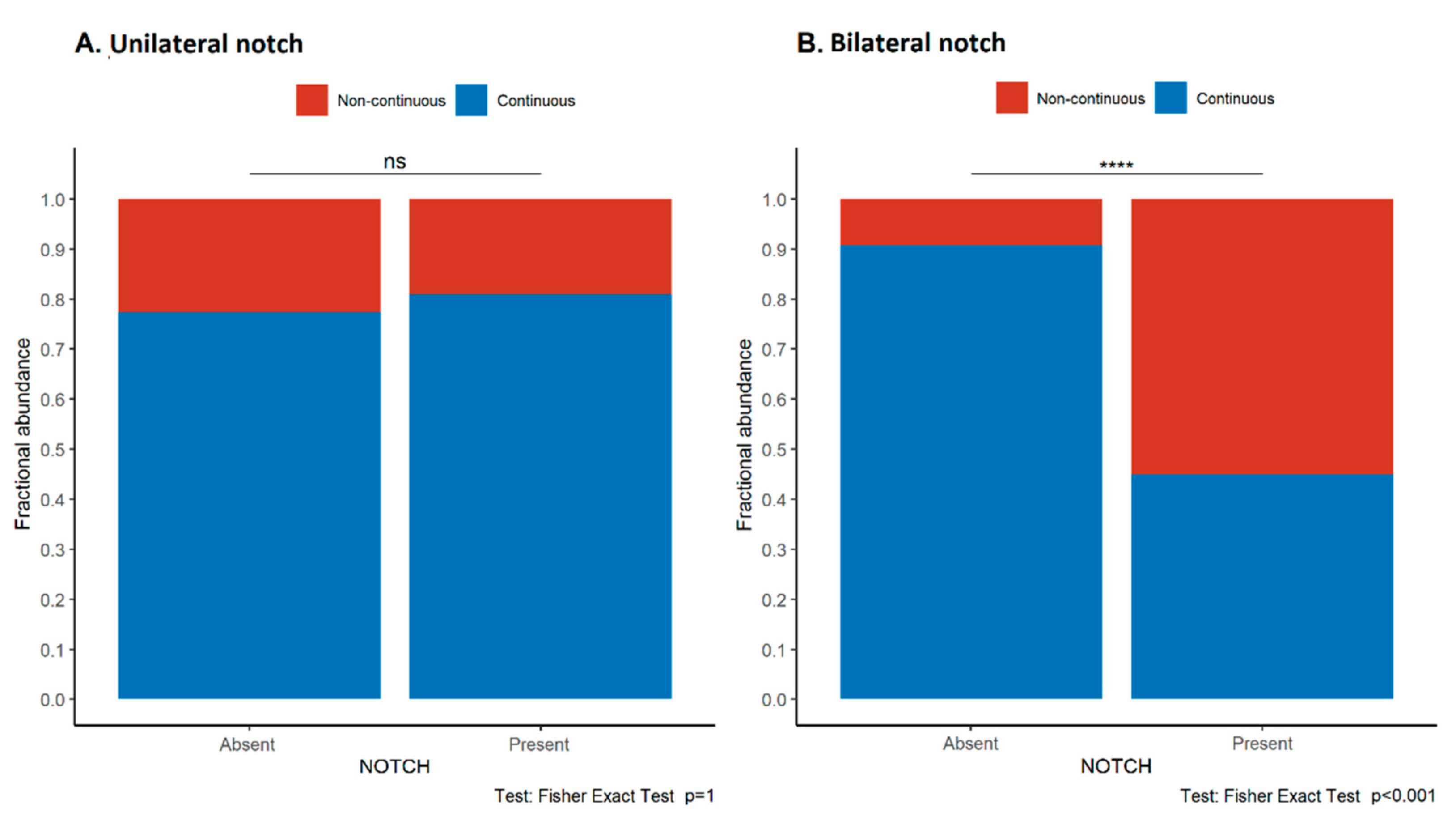
3.5. Loss of E-Cadherin Continuity Is Associated with the Presence of an Early Diastolic Notch in the Uterine Arteries, Maternal Proteinuria, and Lower Apgar Scores
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poon, L.C.; Shennan, A.; Hyett, J.A.; Kapur, A.; Hadar, E.; Divakar, H.; McAuliffe, F.; Silva Costa, F.; Dadelszen, P.; McIntyre, H.D.; et al. The International Federation of Gynecology and Obstetrics (FIGO) initiative on pre-eclampsia: A pragmatic guide for first-trimester screening and prevention. Int. J. Gynecol. Obstet. 2019, 145, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Tsakiridis, I.; Giouleka, S.; Arvanitaki, A.; Giannakoulas, G.; Papazisis, G.; Mamopoulos, A.; Athanasiadis, A.; Dagklis, T. Gestational Hypertension and Preeclampsia: An Overview of National and International Guidelines. Obstet. Gynecol. Surv. 2021, 76, 613–633. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. The hypertensive disorders of pregnancy: ISSHP classification, diagnosis & management recommendations for international practice. Pregnancy Hypertens. 2018, 13, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Task Force on Hypertension in Pregnancy. Hypertension in Pregnancy. Obstet. Gynecol. 2013, 122, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Webster, K.; Fishburn, S.; Maresh, M.; Findlay, S.C.; Chappell, L.C. Diagnosis and management of hypertension in pregnancy: Summary of updated NICE guidance. BMJ 2019, 366, l5119. [Google Scholar] [CrossRef]
- Homer, C.S.; Brown, M.A.; Mangos, G.; Davis, G.K. Non-proteinuric pre-eclampsia: A novel risk indicator in women with gestational hypertension. J. Hypertens. 2008, 26, 295–302. [Google Scholar] [CrossRef]
- Roland, C.S.; Hu, J.; Ren, C.-E.; Chen, H.; Li, J.; Varvoutis, M.S.; Leaphart, L.W.; Byck, D.B.; Zhu, X.; Jiang, S.-W. Morphological changes of placental syncytium and their implications for the pathogenesis of preeclampsia. Cell. Mol. Life Sci. 2016, 73, 365–376. [Google Scholar] [CrossRef]
- Gude, N.M.; Roberts, C.T.; Kalionis, B.; King, R.G. Growth and function of the normal human placenta. Thromb. Res. 2004, 114, 397–407. [Google Scholar] [CrossRef]
- Knöfler, M.; Pollheimer, J. Human placental trophoblast invasion and differentiation: A particular focus on Wnt signaling. Front. Genet. 2013, 4, 190. [Google Scholar] [CrossRef]
- Moser, G.; Windsperger, K.; Pollheimer, J.; de Sousa Lopes, S.C.; Huppertz, B. Human trophoblast invasion: New and unexpected routes and functions. Histochem. Cell Biol. 2018, 150, 361–370. [Google Scholar] [CrossRef]
- Ishiyama, N.; Lee, S.-H.; Liu, S.; Li, G.-Y.; Smith, M.J.; Reichardt, L.F.; Ikura, M. Dynamic and Static Interactions between p120 Catenin and E-Cadherin Regulate the Stability of Cell-Cell Adhesion. Cell 2010, 141, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Venhuizen, J.-H.; Jacobs, F.J.C.; Span, P.N.; Zegers, M.M. P120 and E-cadherin: Double-edged swords in tumor metastasis. Semin. Cancer Biol. 2020, 60, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Floridon, C.; Nielsen, O.; Holund, B.; Sunde, L.; Westergaard, J.G.; Thomsen, S.G.; Teisner, B. Localization of E-cadherin in villous, extravillous and vascular trophoblasts during intrauterine, ectopic and molar pregnancy. Mol. Hum. Reprod. 2000, 6, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Vercruysse, L.; Hanssens, M. The Uterine Spiral Arteries In Human Pregnancy: Facts and Controversies. Placenta 2006, 27, 939–958. [Google Scholar] [CrossRef] [PubMed]
- Bartels, H.C.; Postle, J.D.; Downey, P.; Brennan, D.J. Placenta Accreta Spectrum: A Review of Pathology, Molecular Biology, and Biomarkers. Dis. Markers 2018, 2018, 1507674. [Google Scholar] [CrossRef]
- Ghosh, G.; Breborowicz, A.; Brązert, M.; Maczkiewicz, M.; Kobelski, M.; Dubiel, M.; Gudmundsson, S. Evaluation of third trimester uterine artery flow velocity indices in relationship to perinatal complications. J. Matern. Neonatal Med. 2006, 19, 551–555. [Google Scholar] [CrossRef]
- North, R.A.; Ferrier, C.; Long, D.; Townend, K.; Kincaid-Smith, P. Uterine artery Doppler flow velocity waveforms in the second trimester for the prediction of preeclampsia and fetal growth retardation. Obstet. Gynecol. 1994, 83, 378–386. [Google Scholar]
- Napolitano, R.; Rajakulasingam, R.; Memmo, A.; Bhide, A.; Thilaganathan, B. Uterine artery Doppler screening for pre-eclampsia: Comparison of the lower, mean and higher first-trimester pulsatility indices. Ultrasound Obstet. Gynecol. 2011, 37, 534–537. [Google Scholar] [CrossRef]
- Park, Y.W.; Cho, J.S.; Choi, H.M.; Kim, T.Y.; Lee, S.H.; Yu, J.K.; Kim, J.W. Clinical significance of early diastolic notch depth: Uterine artery Doppler velocimetry in the third trimester. Am. J. Obstet. Gynecol. 2000, 182, 1204–1209. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24275-0. [Google Scholar]
- Madden, J.V.; Flatley, C.J.; Kumar, S. Term small-for-gestational-age infants from low-risk women are at significantly greater risk of adverse neonatal outcomes. Am. J. Obstet. Gynecol. 2018, 218, 525.e1–525.e9. [Google Scholar] [CrossRef]
- Baulon, E.; Fraser, W.D.; Piedboeuf, B.; Buekens, P.; Xiong, X. Pregnancy-induced hypertension and infant growth at 28 and 42 days postpartum. BMC Pregnancy Childbirth 2005, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Bokslag, A.; van Weissenbruch, M.; Mol, B.W.; de Groot, C.J.M. Preeclampsia; short and long-term consequences for mother and neonate. Early Hum. Dev. 2016, 102, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, T.V.; Brunetto, S.; Ramos, J.G.L.; Bernardi, J.R.; Goldani, M.Z. Hypertensive disorders during pregnancy and health outcomes in the offspring: A systematic review. J. Dev. Orig. Health Dis. 2016, 7, 391–407. [Google Scholar] [CrossRef]
- Redman, C.W.G.; Sargent, I.L. Circulating Microparticles in Normal Pregnancy and Pre-Eclampsia. Placenta 2008, 29, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Woolcock, J.; Hennessy, A.; Xu, B.; Thornton, C.; Tooher, J.; Makris, A.; Ogle, R. Soluble Flt-1 as a diagnostic marker of pre-eclampsia. Aust. N. Z. J. Obstet. Gynaecol. 2008, 48, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Li, J.; Wang, F.; Oliver, M.T.; Tipton, T.; Gao, Y.; Jiang, S.-W. Syncytin-1 modulates placental trophoblast cell proliferation by promoting G1/S transition. Cell Signal. 2013, 25, 1027–1035. [Google Scholar] [CrossRef]
- Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.-P.; Skepper, J.N.; Burton, G.J. Onset of Maternal Arterial Blood Flow and Placental Oxidative Stress. Am. J. Pathol. 2000, 157, 2111–2122. [Google Scholar] [CrossRef]
- Vargas, A.; Toufaily, C.; LeBellego, F.; Rassart, É.; Lafond, J.; Barbeau, B. Reduced Expression of Both Syncytin 1 and Syncytin 2 Correlates With Severity of Preeclampsia. Reprod. Sci. 2011, 18, 1085–1091. [Google Scholar] [CrossRef]
- Ruebner, M.; Strissel, P.L.; Ekici, A.B.; Stiegler, E.; Dammer, U.; Goecke, T.W.; Faschingbauer, F.; Fahlbusch, F.B.; Beckmann, M.W.; Strick, R. Reduced Syncytin-1 Expression Levels in Placental Syndromes Correlates with Epigenetic Hypermethylation of the ERVW-1 Promoter Region. PLoS ONE 2013, 8, e56145. [Google Scholar] [CrossRef]
- Zhuang, X.-W.; Li, J.; Brost, B.; Xia, X.-Y.; Chen, H.; Wang, C.-X.; Jiang, S.-W. Decreased Expression and Altered Methylation of Syncytin-1 Gene in Human Placentas Associated with Preeclampsia. Curr. Pharm. Des. 2014, 20, 1796–1802. [Google Scholar] [CrossRef]
- Lee, X.; Keith, J.C.; Stumm, N.; Moutsatsos, I.; McCoy, J.M.; Crum, C.P.; Genest, D.; Chin, D.; Ehrenfels, C.; Pijnenborg, R.; et al. Downregulation of Placental Syncytin Expression and Abnormal Protein Localization in Pre-eclampsia. Placenta 2001, 22, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Tannetta, D.; Masliukaite, I.; Vatish, M.; Redman, C.; Sargent, I. Update of syncytiotrophoblast derived extracellular vesicles in normal pregnancy and preeclampsia. J. Reprod. Immunol. 2017, 119, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Mayhew, T.M. Turnover of human villous trophoblast in normal pregnancy: What do we know and what do we need to know? Placenta 2014, 35, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Guo, F.; Jin, H.Y.; Lau, S.; Stone, P.; Chamley, L. Phagocytosis of apoptotic trophoblastic debris protects endothelial cells against activation. Placenta 2012, 33, 548–553. [Google Scholar] [CrossRef]
- Tannetta, D.S.; Hunt, K.; Jones, C.I.; Davidson, N.; Coxon, C.H.; Ferguson, D.; Redman, C.W.; Gibbins, J.M.; Sargent, I.L.; Tucker, K.L. Syncytiotrophoblast Extracellular Vesicles from Pre-Eclampsia Placentas Differentially Affect Platelet Function. PLoS ONE 2015, 10, e0142538. [Google Scholar] [CrossRef]
- Tannetta, D.; Collett, G.; Vatish, M.; Redman, C.; Sargent, I. Syncytiotrophoblast extracellular vesicles—Circulating biopsies reflecting placental health. Placenta 2017, 52, 134–138. [Google Scholar] [CrossRef]
- Vargas, A.; Zhou, S.; Éthier-Chiasson, M.; Flipo, D.; Lafond, J.; Gilbert, C.; Barbeau, B. Syncytin proteins incorporated in placenta exosomes are important for cell uptake and show variation in abundance in serum exosomes from patients with preeclampsia. FASEB J. 2014, 28, 3703–3719. [Google Scholar] [CrossRef]
- Zhu, S.; Zhao, G.; Zhao, X.; Zhan, X.; Cai, M.; Geng, C.; Pu, Q.; Zhao, Q.; Fu, Q.; Huang, C.; et al. Elevated soluble E-cadherin during the epithelial-mesenchymal transition process and as a diagnostic marker in colorectal cancer. Gene 2020, 754, 144899. [Google Scholar] [CrossRef]
- Hansson, S.R.; Nääv, Å.; Erlandsson, L. Oxidative stress in preeclampsia and the role of free fetal hemoglobin. Front. Physiol. 2015, 5, 516. [Google Scholar] [CrossRef]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef]
- Khong, T.Y.; Mooney, E.E.; Ariel, I.; Balmus, N.C.M.; Boyd, T.K.; Brundler, M.-A.; Derricott, H.; Evans, M.J.; Faye-Petersen, O.M.; Gillan, J.E.; et al. Sampling and Definitions of Placental Lesions: Amsterdam Placental Workshop Group Consensus Statement. Arch. Pathol. Lab. Med. 2016, 140, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Catov, J.M.; Scifres, C.M.; Caritis, S.N.; Bertolet, M.; Larkin, J.; Parks, W.T. Neonatal outcomes following preterm birth classified according to placental features. Am. J. Obstet. Gynecol. 2017, 216, 411.e1–411.e14. [Google Scholar] [CrossRef] [PubMed]
- Sebire, N.J. Placental histology findings in relation to pre-eclampsia: Implications for interpretation of retrospective studies. Ultrasound Obstet. Gynecol. 2017, 50, 291–292. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Group (n = 55) | Control Group (n = 37) | p Value | ||
|---|---|---|---|---|
| Gravida | 1 | n = 37 | n = 20 | |
| 2 | n = 11 | n = 12 | 0.367 | |
| ≥3 | n = 7 | n = 5 | ||
| n/a | n = 0 | n = 0 | ||
| Newborn | D/L/T | n = 16 | n = 20 | |
| D/L/P | n = 18 | n = 0 | ||
| S/L/T | n = 9 | n = 16 | <0.001 | |
| S/L/P | n = 10 | n = 0 | ||
| n/a | n = 2 | n = 1 | ||
| Delivery | cc | n = 44 | n = 7 | |
| vaginal | n = 10 | n = 29 | <0.001 | |
| n/a | n = 1 | n = 1 | ||
| Apgar score | >7 | n = 38 | n = 30 | |
| ≤7 | n = 10 | n = 0 | <0.001 | |
| n/a | n = 7 | n = 8 | ||
| Gestational age | <32 weeks | n = 8 | n = 0 | |
| ≥32 weeks | n = 47 | n = 37 | 0.015 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pęksa, M.; Kamieniecki, A.; Gabrych, A.; Lew-Tusk, A.; Preis, K.; Świątkowska-Freund, M. Loss of E-Cadherin Staining Continuity in the Trophoblastic Basal Membrane Correlates with Increased Resistance in Uterine Arteries and Proteinuria in Patients with Pregnancy-Induced Hypertension. J. Clin. Med. 2022, 11, 668. https://doi.org/10.3390/jcm11030668
Pęksa M, Kamieniecki A, Gabrych A, Lew-Tusk A, Preis K, Świątkowska-Freund M. Loss of E-Cadherin Staining Continuity in the Trophoblastic Basal Membrane Correlates with Increased Resistance in Uterine Arteries and Proteinuria in Patients with Pregnancy-Induced Hypertension. Journal of Clinical Medicine. 2022; 11(3):668. https://doi.org/10.3390/jcm11030668
Chicago/Turabian StylePęksa, Marta, Alexandra Kamieniecki, Anna Gabrych, Anna Lew-Tusk, Krzysztof Preis, and Małgorzata Świątkowska-Freund. 2022. "Loss of E-Cadherin Staining Continuity in the Trophoblastic Basal Membrane Correlates with Increased Resistance in Uterine Arteries and Proteinuria in Patients with Pregnancy-Induced Hypertension" Journal of Clinical Medicine 11, no. 3: 668. https://doi.org/10.3390/jcm11030668
APA StylePęksa, M., Kamieniecki, A., Gabrych, A., Lew-Tusk, A., Preis, K., & Świątkowska-Freund, M. (2022). Loss of E-Cadherin Staining Continuity in the Trophoblastic Basal Membrane Correlates with Increased Resistance in Uterine Arteries and Proteinuria in Patients with Pregnancy-Induced Hypertension. Journal of Clinical Medicine, 11(3), 668. https://doi.org/10.3390/jcm11030668