
Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function


Abstract
:1. Introduction
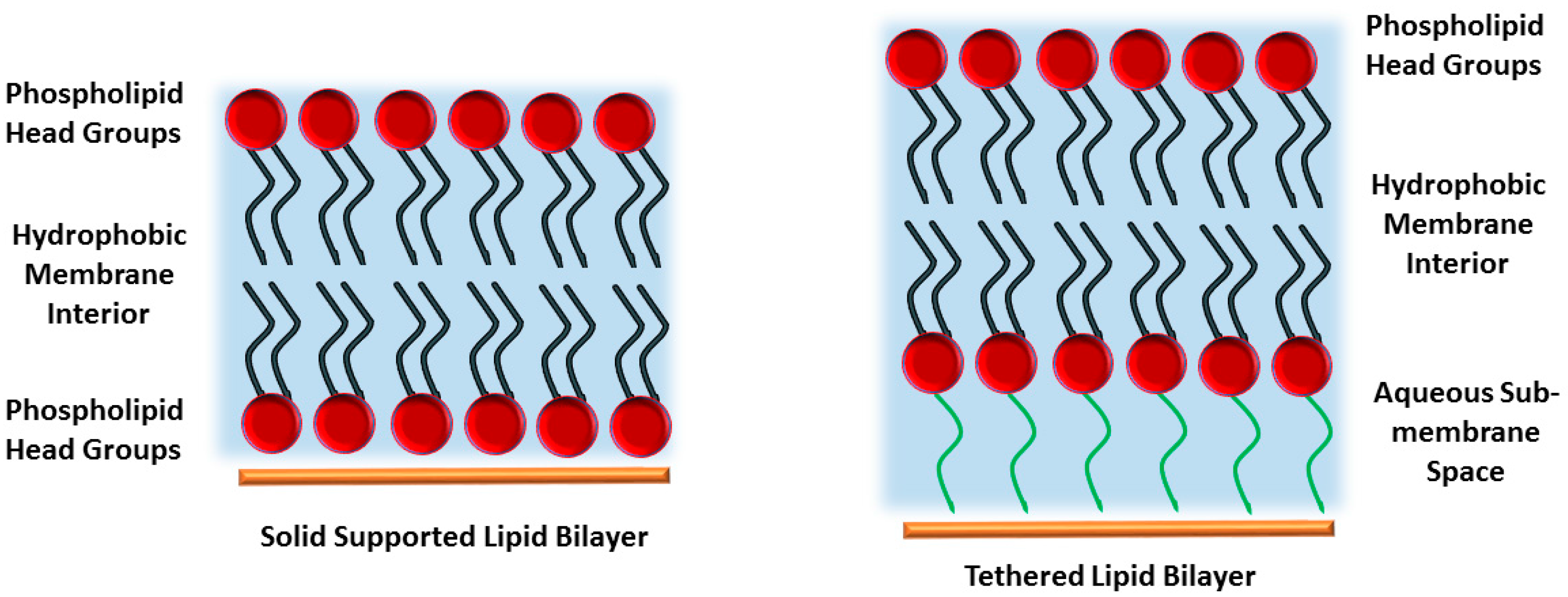
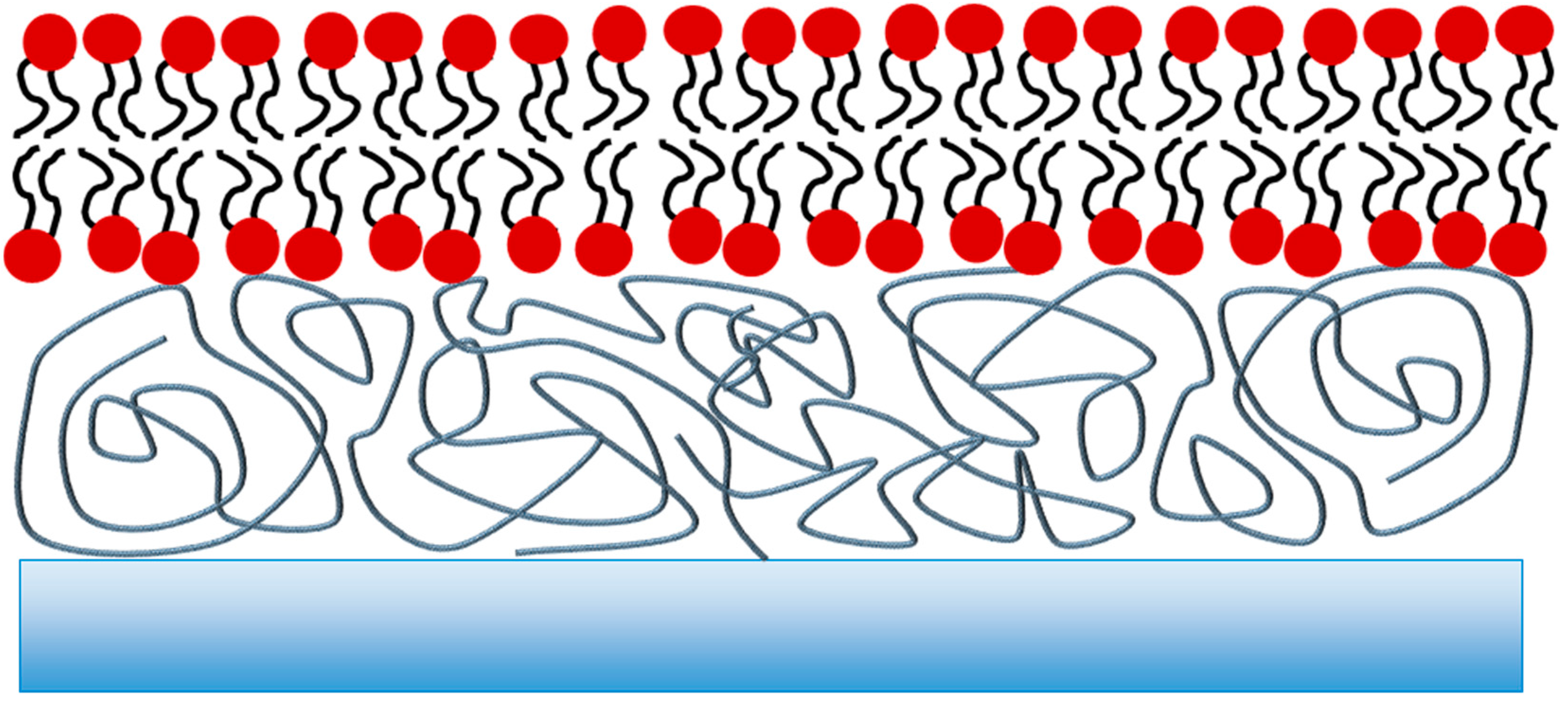
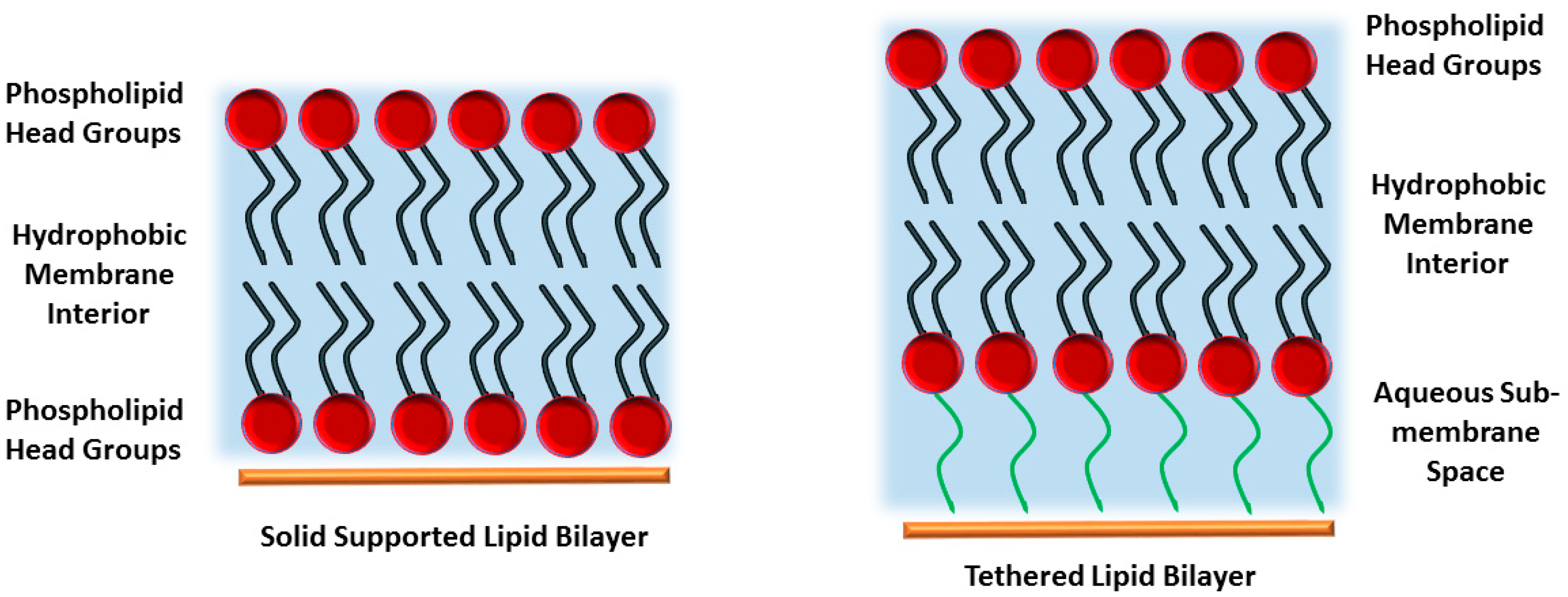
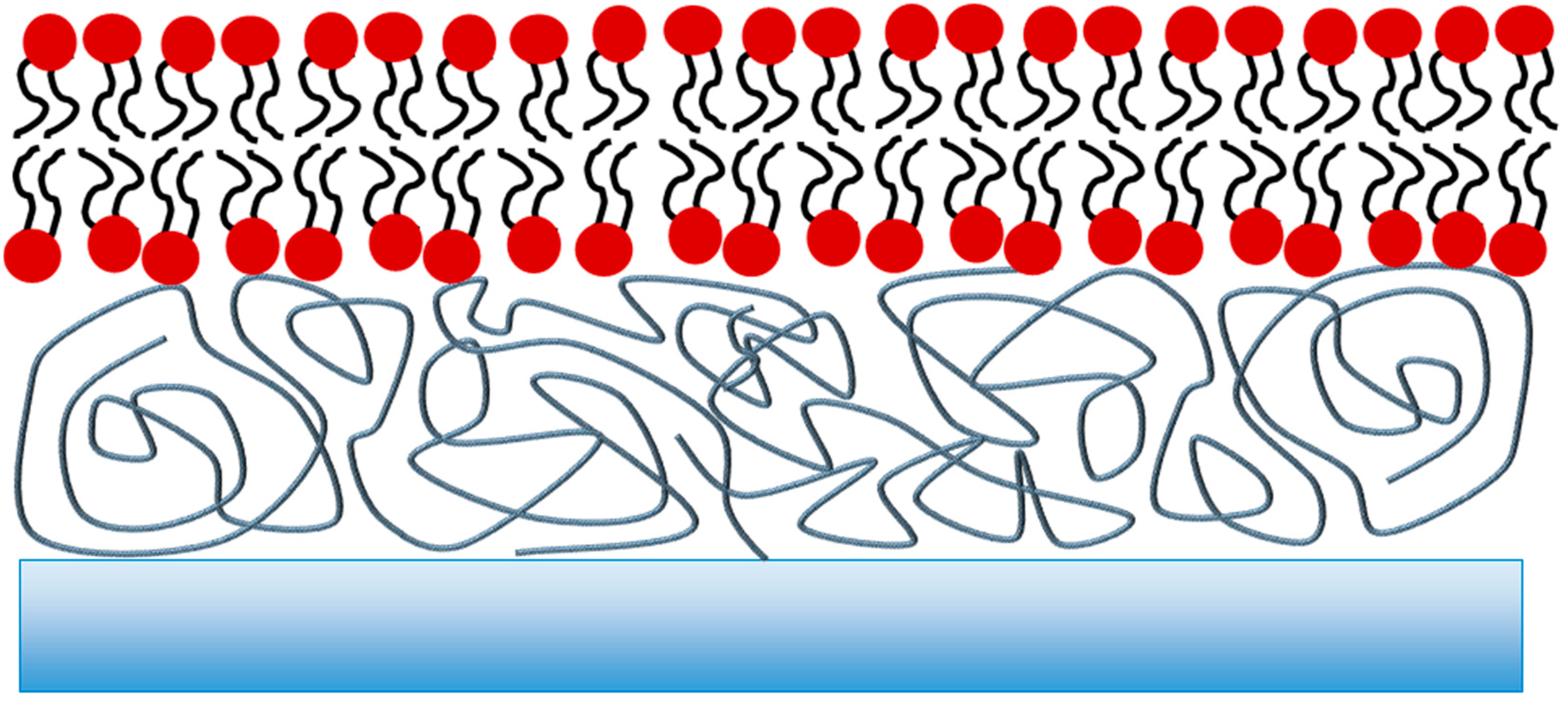
2. Polymer-Supported Lipid Bilayers
Effect of the Supporting Polymer on Bilayer Properties
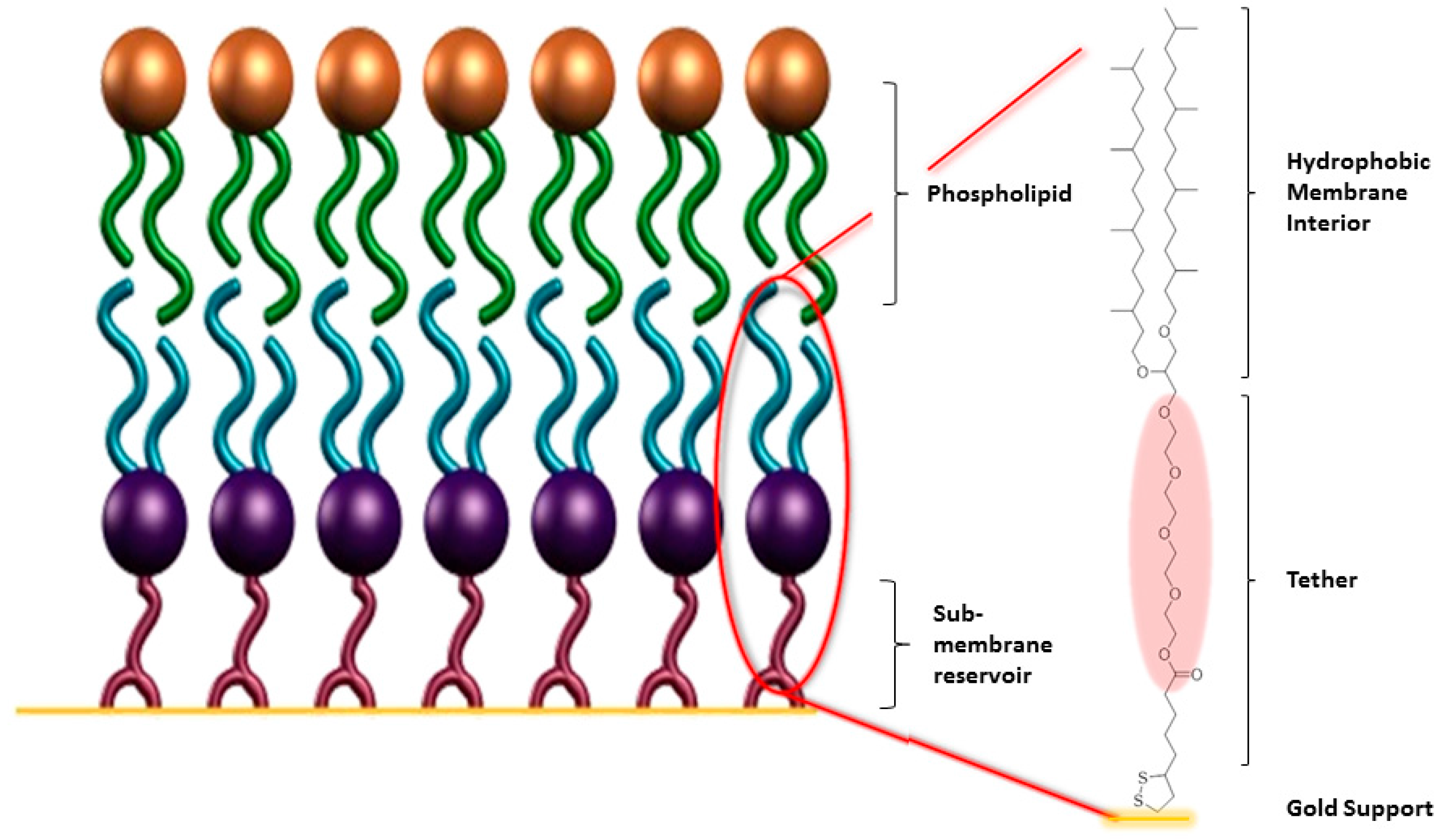
3. Tethered Bilayer Lipid Membranes
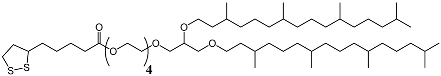
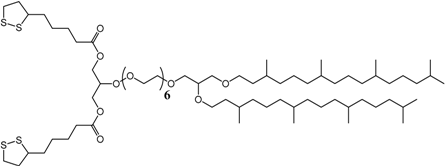
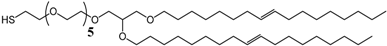
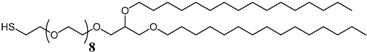
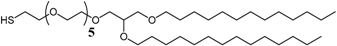
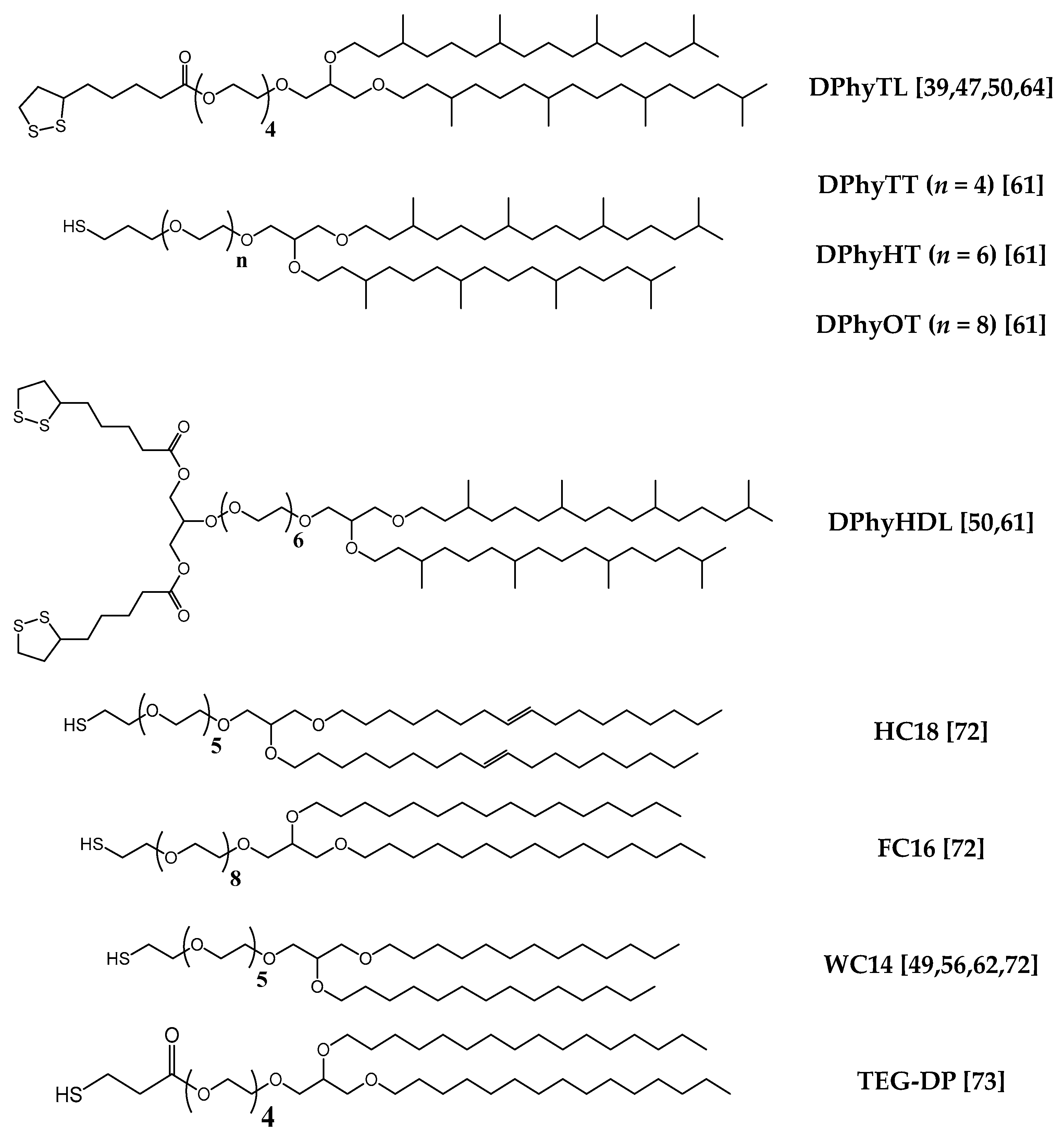
3.1. Effect of the Tether Structure on Bilayer Properties
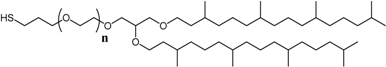
3.2. Sparsely Tethered Bilayer Lipid Membranes
4. Conclusions
Conflicts of Interest
References
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal structure of rhodopsin: A G protein-coupled receptor. Science 2000, 289, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Leutenegger, M.; Lasser, T.; Sinner, E.K.; Robelek, R. Imaging of G protein-coupled receptors in solid-supported planar lipid membranes. Biointerphases 2008, 3, FA136–FA145. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, M.; Chikura, S.; Watari, K.; Mizuno, N.; Mochinaga, K.; Mangmool, S.; Koyanagi, S.; Ohdo, S.; Sato, Y.; Ide, T.; et al. Induction of Cardiac Fibrosis by beta-Blocker in G Proteinin-dependent and G Protein-coupled Receptor Kinase 5/beta-Arrestin2-dependent Signaling Pathways. J. Biol. Chem. 2012, 287, 35669–35677. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, K. Molecular mechanisms of ligand binding, signaling, and regulation within the superfamily of G-protein-coupled receptors: Molecular modeling and mutagenesis approaches to receptor structure and function. Pharmacol. Ther. 2004, 103, 21–80. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.H.; Gregory, R.J.; Marshall, J.; Paul, S.; Souza, D.W.; White, G.A.; Oriordan, C.R.; Smith, A.E. Defective intracellular-transport and processing of CFTR is the molecular basis of most cystic-fibrosis. Cell 1990, 63, 827–834. [Google Scholar] [CrossRef]
- Emery, A.E.H. The muscular dystrophies. Lancet 2002, 359, 687–695. [Google Scholar] [CrossRef]
- Klabunde, T.; Hessler, G. Drug design strategies for targeting G-protein-coupled receptors. Chembiochem. Eur. J. Chem. Biol. 2002, 3, 928–944. [Google Scholar] [CrossRef]
- Guo, D.; Hillger, J.M.; Ijzerman, A.P.; Heitman, L.H. Drug-Target Residence Time—A Case for G Protein-Coupled Receptors. Med. Res. Rev. 2014, 34, 856–892. [Google Scholar] [CrossRef] [PubMed]
- Knobloch, J.; Suhendro, D.K.; Zieleniecki, J.L.; Shapter, J.G.; Köper, I. Membrane—Drug interactions studied using model membrane systems. Saudi J. Biol. Sci. 2015, 22, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Sackmann, E. Supported membranes: Scientific and practical applications. Science 1996, 271, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Sackmann, E. Polymer-supported membranes as models of the cell surface. Nature 2005, 437, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Winterhalter, M. Black lipid membranes. Cur. Opin. Colloid Interface Sci. 2000, 5, 250–255. [Google Scholar] [CrossRef]
- Chan, Y.H.M.; Boxer, S.G. Model membrane systems and their applications. Cur. Opin. Chem. Biol. 2007, 11, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Castellana, E.T.; Cremer, P.S. Solid supported lipid bilayers: From biophysical studies to sensor design. Surface Sci. Rep. 2006, 61, 429–444. [Google Scholar] [CrossRef]
- Johnson, S.J.; Bayerl, T.M.; McDermott, D.C.; Adam, G.W.; Rennie, A.R.; Thomas, R.K.; Sackmann, E. Structure of an adsorbed dimyristoylphosphatidylcholine bilayer measured with specular reflection of neutrons. Biophys. J. 1991, 59, 289–294. [Google Scholar] [CrossRef]
- Koenig, B.W.; Krueger, S.; Orts, W.J.; Majkrzak, C.F.; Berk, N.F.; Silverton, J.V.; Gawrisch, K. Neutron Reflectivity and Atomic Force Microscopy Studies of a Lipid Bilayer in Water Adsorbed to the Surface of a Silicon Single Crystal. Langmuir 1996, 12, 1343–1350. [Google Scholar] [CrossRef]
- Rinia, H.A.; Snel, M.M.E.; van der Eerden, J.; de Kruijff, B. Visualizing detergent resistant domains in model membranes with atomic force microscopy. FEBS Lett. 2001, 501, 92–96. [Google Scholar] [CrossRef]
- Kiessling, V.; Crane, J.M.; Tamm, L.K. Transbilayer effects of raft-like lipid domains in asymmetric planar bilayers measured by single molecule tracking. Biophys. J. 2006, 91, 3313–3326. [Google Scholar] [CrossRef] [PubMed]
- Tokumasu, F.; Jin, A.J.; Feigenson, G.W.; Dvorak, J.A. Nanoscopic lipid domain dynamics revealed by atomic force microscopy. Biophys. J. 2003, 84, 2609–2618. [Google Scholar] [CrossRef]
- Wildman, K.A.H.; Lee, D.K.; Ramamoorthy, A. Mechanism of lipid bilayer disruption by the human antimicrobial peptide, LL-37. Biochemistry 2003, 42, 6545–6558. [Google Scholar] [CrossRef] [PubMed]
- Urbina, J.A.; Pekerar, S.; Le, H.B.; Patterson, J.; Montez, B.; Oldfield, E. Molecular order and dynamics of phosphatidycholine bilayer-membranes in the presence of cholesterol, ergosterol and lanosterol—A comparative study using H-2-NMR, C-13-NMR and P-31-NMR Spectroscopy. Biochim. Et Biophys. Acta-Biomembr. 1995, 1238, 163–176. [Google Scholar] [CrossRef]
- Rinia, H.A.; Kik, R.A.; Demel, R.A.; Snel, M.M.E.; Killian, J.A.; van der Eerden, J.; de Kruijff, B. Visualization of highly ordered striated domains induced by transmembrane peptides in supported phosphatidylcholine bilayers. Biochemistry 2000, 39, 5852–5858. [Google Scholar] [CrossRef] [PubMed]
- Naumann, C.A.; Prucker, O.; Lehmann, T.; Rühe, J.; Knoll, W.; Frank, C.W. The polymer-supported phospholipid bilayer: Tethering as a new approach to substrate-membrane stabilization. Biomacromolecules 2002, 3, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.L.; Tamm, L.K. Tethered polymer-supported planar lipid bilayers for reconstitution of integral membrane proteins: Silane-polyethyleneglycol-lipid as a cushion and covalent linker. Biophys. J. 2000, 79, 1400–1414. [Google Scholar] [CrossRef]
- Su, Z.; Jiang, Y.; Velázquez-Manzanares, M.; Leitch, J.J.; Kycia, A.; Lipkowski, J. Electrochemical and PM-IRRAS studies of floating lipid bilayers assembled at the Au (111) electrode pre-modified with a hydrophilic monolayer. J. Electroanal. Chem. 2013, 688, 76–85. [Google Scholar] [CrossRef]
- Deverall, M.A.; Gindl, E.; Sinner, E.K.; Besir, H.; Ruehe, J.; Saxton, M.J.; Naumann, C.A. Membrane Lateral Mobility Obstructed by Polymer-Tethered Lipids Studied at the Single Molecule Level. Biophys. J. 2005, 88, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, V.; Alves, I.D.; Salgado, G.F.J.; Lau, P.-W.; Wysocki, R.J.; Salamon, Z.; Tollin, G.; Hruby, V.J.; Brown, M.F.; Saavedra, S.S. Rhodopsin Reconstituted into a Planar-Supported Lipid Bilayer Retains Photoactivity after Cross-Linking Polymerization of Lipid Monomers. J. Am. Chem. Soc. 2005, 127, 5320–5321. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.L.; Tamm, L.K. Reconstituted Syntaxin1A/SNAP25 Interacts with Negatively Charged Lipids as Measured by Lateral Diffusion in Planar Supported Bilayers. Biophys. J. 2001, 81, 266–275. [Google Scholar] [CrossRef]
- Kiessling, V.; Tamm, L.K. Measuring Distances in Supported Bilayers by Fluorescence Interference-Contrast Microscopy: Polymer Supports and SNARE Proteins. Biophys. J. 2003, 84, 408–418. [Google Scholar] [CrossRef]
- Rossi, C.; Chopineau, J. Biomimetic tethered lipid membranes designed for membrane-protein interaction studies. Eur. Biophys. J. 2007, 36, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Purrucker, O.; Gonnenwein, S.; Fortig, A.; Jordan, R.; Rusp, M.; Barmann, M.; Moroder, L.; Sackmann, E.; Tanaka, M. Polymer-tethered membranes as quantitative models for the study of integrin-mediated cell adhesion. Soft Matter 2007, 3, 333–336. [Google Scholar] [CrossRef]
- Beyer, D.; Elender, G.; Knoll, W.; Kühner, M.; Maus, S.; Ringsdorf, H.; Sackmann, E. Influence of anchor lipids on the homogeneity and mobility of lipid bilayers on thin polymer films. Angew. Chem. Int. Ed. Engl. 1996, 35, 1682–1685. [Google Scholar] [CrossRef]
- Purrucker, O.; Förtig, A.; Jordan, R.; Tanaka, M. Supported Membranes with Well-Defined Polymer Tethers—Incorporation of Cell Receptors. ChemPhysChem 2004, 5, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Renner, L.; Pompe, T.; Lemaitre, R.; Drechsel, D.; Werner, C. Controlled enhancement of transmembrane enzyme activity in polymer cushioned supported bilayer membranes. Soft Matter 2010, 6, 5382–5389. [Google Scholar] [CrossRef]
- Blakeston, A.C.; Alswieleh, A.M.; Heath, G.R.; Roth, J.S.; Bao, P.; Cheng, N.; Armes, S.P.; Leggett, G.J.; Bushby, R.J.; Evans, S.D. New Poly(amino acid methacrylate) Brush Supports the Formation of Well-Defined Lipid Membranes. Langmuir 2015, 31, 3668–3677. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Konradi, R.; Textor, M.; Reimhult, E. Liposomes Tethered to Omega-Functional PEG Brushes and Induced Formation of PEG Brush Supported Planar Lipid Bilayers. Langmuir 2009, 25, 13534–13539. [Google Scholar] [CrossRef] [PubMed]
- Hertrich, S.; Stetter, F.; Rühm, A.; Hugel, T.; Nickel, B. Highly Hydrated Deformable Polyethylene Glycol-Tethered Lipid Bilayers. Langmuir 2014, 30, 9442–9447. [Google Scholar] [CrossRef] [PubMed]
- Kibrom, A.; Roskamp, R.F.; Jonas, U.; Menges, B.; Knoll, W.; Paulsen, H.; Naumann, R.L. Hydrogel-supported protein-tethered bilayer lipid membranes: A new approach toward polymer-supported lipid membranes. Soft Matter 2011, 7, 237–246. [Google Scholar] [CrossRef]
- Schiller, S.M.; Naumann, R.; Lovejoy, K.; Kunz, H.; Knoll, W. Archaea analogue thiolipids for tethered bilayer lipid membranes on ultrasmooth gold surfaces. Angew. Chem. Int. Ed. 2003, 42, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Köper, I. Insulating tethered bilayer lipid membranes to study membrane proteins. Mol. Biosyst. 2007, 3, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Cornell, B.A.; BraachMaksvytis, V.L.B.; King, L.G.; Osman, P.D.J.; Raguse, B.; Wieczorek, L.; Pace, R.J. A biosensor that uses ion-channel switches. Nature 1997, 387, 580. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, V.; Knorr, N.; Duran, R.S.; Ingebrandt, S.; Offenhäusser, A.; Knoll, W.; Köper, I. Membrane on a chip: A functional tethered lipid bilayer membrane on silicon oxide surfaces. Biophys. J. 2005, 89, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Roskamp, R.F.; Vockenroth, I.K.; Eisenmenger, N.; Braunagel, J.; Koeper, I. Functional tethered bilayer lipid membranes on aluminum oxide. ChemPhysChem 2008, 9, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
- Becucci, L.; Guidelli, R. Mercury-supported biomimetic membranes for the investigation of antimicrobial peptides. Pharmaceuticals 2014, 7, 136–168. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.; Moldovan, R.; Fitzpatrick, J.; Vanderah, D.J.; Deserno, M.; Losche, M. In-plane homogeneity and lipid dynamics in tethered bilayer lipid membranes (tBLMs). Soft Matter 2010, 6, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Knoll, W.; Köper, I.; Naumann, R.; Sinner, E.-K. Tethered bimolecular lipid membranes—A novel model membrane platform. Electrochim. Acta 2008, 53, 6680–6689. [Google Scholar] [CrossRef]
- Junghans, A.; Champagne, C.; Cayot, P.; Loupiac, C.; Köper, I. Probing Protein—Membrane Interactions Using Solid Supported Membranes. Langmuir 2011, 27, 2709–2716. [Google Scholar] [CrossRef] [PubMed]
- Vockenroth, I.K.; Atanasova, P.P.; Long, J.R.; Jenkins, A.T.A.; Knoll, W.; Köper, I. Functional incorporation of the pore forming segment of AChR M2 into tethered bilayer lipid membranes. Biochim. Biophys. Acta Biomembr. 2007, 1768, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- McGillivray, D.J.; Valincius, G.; Heinrich, F.; Robertson, J.W.; Vanderah, D.J.; Febo-Ayala, W.; Ignatjev, I.; Lösche, M.; Kasianowicz, J.J. Structure of functional Staphylococcus aureus α-hemolysin channels in tethered bilayer lipid membranes. Biophys. J. 2009, 96, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Vockenroth, I.K.; Atanasova, P.P.; Jenkins, A.T.A.; Koper, I. Incorporation of alpha-hemolysin in different tethered bilayer lipid membrane architectures. Langmuir 2008, 24, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Becucci, L.; Moncelli, M.R.; Naumann, R.; Guidelli, R. Potassium ion transport by valinomycin across a Hg-supported lipid bilayer. J. Am. Chem. Soc. 2005, 127, 13316–13323. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, S.R.; Sui, D.; Garavito, R.M.; Worden, R.M. Fabrication of highly insulating tethered bilayer lipid membrane using yeast cell membrane fractions for measuring ion channel activity. J. Colloid Interface Sci. 2008, 322, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.A.; Heinrich, F.; Raghunandan, S.; Krueger, S.; Curtis, J.E.; Rein, A.; Nanda, H. HIV-1 Gag extension: Conformational changes require simultaneous interaction with membrane and nucleic acid. J. Mol. Biol. 2011, 406, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Haass, C.; Selkoe, D.J. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef]
- Lesné, S.; Koh, M.T.; Kotilinek, L.; Kayed, R.; Glabe, C.G.; Yang, A.; Gallagher, M.; Ashe, K.H. A specific amyloid-[beta] protein assembly in the brain impairs memory. Nature 2006, 440, 352–357. [Google Scholar] [CrossRef]
- Valincius, G.; Heinrich, F.; Budvytyte, R.; Vanderah, D.J.; McGillivray, D.J.; Sokolov, Y.; Hall, J.E.; Losche, M. Soluble Amyloid beta-Oligomers Affect Dielectric Membrane Properties by Bilayer Insertion and Domain Formation: Implications for Cell Toxicity. Biophys. J. 2008, 95, 4845–4861. [Google Scholar] [CrossRef]
- Jeuken, L.J.C.; Connell, S.D.; Henderson, P.J.F.; Gennis, R.B.; Evans, S.D.; Bushby, R.J. Redox Enzymes in Tethered Membranes. J. Am. Chem. Soc. 2006, 128, 1711–1716. [Google Scholar] [CrossRef]
- Goreham, R.V.; Thompson, V.C.; Samura, Y.; Gibson, C.T.; Shapter, J.G.; Köper, I. Interaction of silver nanoparticles with tethered bilayer lipid membranes. Langmuir 2015, 31, 5868–5874. [Google Scholar] [CrossRef]
- Braunagel, J.; Junghans, A.; Köper, I. Membrane-based sensing approaches. Austr. J. Chem. 2011, 64, 54–61. [Google Scholar] [CrossRef]
- Kozuch, J.; Steinem, C.; Hildebrandt, P.; Millo, D. Combined Electrochemistry and Surface-Enhanced Infrared Absorption Spectroscopy of Gramicidin A Incorporated into Tethered Bilayer Lipid Membranes. Angew. Chem. Int. Ed. 2012, 51, 8114–8117. [Google Scholar] [CrossRef] [PubMed]
- Junghans, A.; Köper, I. Structural Analysis of Tethered Bilayer Lipid Membranes. Langmuir 2010, 26, 11035–11040. [Google Scholar] [CrossRef] [PubMed]
- McGillivray, D.J.; Valincius, G.; Vanderah, D.J.; Febo-Ayala, W.; Woodward, J.T.; Heinrich, F.; Kasianowicz, J.J.; Losche, M. Molecular-scale structural and functional characterization of sparsely tethered bilayer lipid membranes. Biointerphases 2007, 2, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Faller, R. Conformational, Dynamical. and Tensional Study of Tethered Bilayer Lipid Membranes in Coarse-Grained Molecular Simulations. Langmuir 2012, 28, 15907–15915. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, V.; Atanasova, P.P.; Vockenroth, I.K.; Knorr, N.; Koper, I. A molecular toolkit for highly insulating tethered bilayer lipid membranes on various substrates. Bioconj. Chem. 2006, 17, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Moncelli, M.; Becucci, L.; Schiller, S.M. Tethered bilayer lipid membranes self-assembled on mercury electrodes. Bioelectrochemistry 2004, 63, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Becucci, L.; Faragher, R.J.; Schwan, A. The effect of the hydrophilic spacer length on the functionality of a mercury-supported tethered bilayer lipid membrane. Bioelectrochemistry 2015, 101, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.D.; Linman, M.J.; Wilkop, T.; Cheng, Q. Regenerable Tethered Bilayer Lipid Membrane Arrays for Multiplexed Label-Free Analysis of Lipid—Protein Interactions on Poly (dimethylsiloxane) Microchips Using SPR Imaging. Anal. Chem. 2009, 81, 1146–1153. [Google Scholar] [CrossRef] [PubMed]
- Schrems, A.; Kibrom, A.; Kupcu, S.; Kiene, E.; Sleytr, U.; Schuster, B. Bilayer lipid membrane formation on a chemically modified S-layer lattice. Langmuir 2011, 27, 3731–3738. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Lowe, R.D.; Chan, Y.-H.M.; Ganesan, P.V.; Boxer, S.G. DNA-tethered membranes formed by giant vesicle rupture. J. Struct. Biol. 2009, 168, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Boxer, S.G. Stability of DNA-Tethered Lipid Membranes with Mobile Tethers. Langmuir 2011, 27, 5492–5497. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.D.; Boxer, S.G. DNA-Based Patterning of Tethered Membrane Patches. Langmuir 2013, 29, 12220–12227. [Google Scholar] [CrossRef] [PubMed]
- Budvytyte, R.; Valincius, G.; Niaura, G.; Voiciuk, V.; Mickevicius, M.; Chapman, H.; Goh, H.Z.; Shekhar, P.; Heinrich, F.; Shenoy, S.; et al. Structure and Properties of Tethered Bilayer Lipid Membranes with Unsaturated Anchor Molecules. Langmuir 2013, 29, 8645–8656. [Google Scholar] [CrossRef] [PubMed]
- Basit, H.; Van der Heyden, A.; Gondran, C.; Nysten, B.; Dumy, P.; Labbe, P. Tethered Bilayer Lipid Membranes on Mixed Self-Assembled Monolayers of a Novel Anchoring Thiol: Impact of the Anchoring Thiol Density on Bilayer Formation. Langmuir 2011, 27, 14317–14328. [Google Scholar] [CrossRef] [PubMed]
- Vockenroth, I.K.; Rossi, C.; Shah, M.R.; Köper, I. Formation of tethered bilayer lipid membranes probed by various surface sensitive techniques. Biointerphases 2009, 4, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Leitch, J.; Kunze, J.; Goddard, J.D.; Schwan, A.L.; Faragher, R.J.; Naumann, R.; Knoll, W.; Dutcher, J.R.; Lipkowski, J. In Situ PM-IRRAS Studies of an Archaea Analogue Thiolipid Assembled on a Au(111) Electrode Surface. Langmuir 2009, 25, 10354–10363. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, F.; Ng, T.; Vanderah, D.J.; Shekhar, P.; Mihailescu, M.; Nanda, H.; Lösche, M. A New Lipid Anchor for Sparsely Tethered Bilayer Lipid Membranes. Langmuir 2009, 25, 4219–4229. [Google Scholar] [CrossRef] [PubMed]
- Kendall, J.K.; Johnson, B.R.; Symonds, P.H.; Imperato, G.; Bushby, R.J.; Gwyer, J.D.; van Berkel, C.; Evans, S.D.; Jeuken, L.J. Effect of the Structure of Cholesterol-Based Tethered Bilayer Lipid Membranes on Ionophore Activity. ChemPhysChem 2010, 11, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Vockenroth, I.K.; Fine, D.; Dodobalapur, A.; Jenkins, A.T.A.; Köper, I. Tethered bilayer lipid membranes with giga-ohm resistances. Electrochem. Commun. 2008, 10, 323–328. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximal Leaflet Composition | Distal Leaflet Composition | Sub-Membrane Hydration (Volume-%) | Membrane Resistance (MΩ·cm2) | Average Lipid Diffusion Constant (μm2·s−1) |
|---|---|---|---|---|
| DPhyTL [61] | DPhyPC | 5 | 3–55 | N/A |
| DPhyTT [61] | DPhyPC | 13 | 3–15 | N/A |
| DPhyHT [61] | DPhyPC | 21 | 2–35 | N/A |
| DPhyOT [61] | DPhyPC | 40 | 0.01–0.09 | N/A |
| DPhyHDL [61] | DPhyPC | 65 | 0.2–6.5 | N/A |
| 30% WC14 [45] | DOPC/DPhyPC | N/A | N/A | 4.1/3.1 |
| 40% WC14 [62] | DPPC | 64 [39] | 0.09 | |
| 70% WC14 [72] | DOPC | N/A | N/A | 1.9 |
| FC 16 [76] | DPhyPC | 53 | 0.1 | N/A |
| 30% FC16 | DOPC/DPhyPC | 53 [76] | N/A | 3.6/2.5 [45] |
| 70% FC16 [72] | DOPC | N/A | N/A | 1.6 |
| 30% HC18 [72] | DOPC | 21 | N/A | 4.1 |
| 20% TEG-DP [73] | POPC | N/A | 2 | N/A |
| 80% Cholesterol-tether [77] | E. coli lipid extract | N/A | 11 | N/A |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andersson, J.; Köper, I. Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function. Membranes 2016, 6, 30. https://doi.org/10.3390/membranes6020030
Andersson J, Köper I. Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function. Membranes. 2016; 6(2):30. https://doi.org/10.3390/membranes6020030
Chicago/Turabian StyleAndersson, Jakob, and Ingo Köper. 2016. "Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function" Membranes 6, no. 2: 30. https://doi.org/10.3390/membranes6020030
APA StyleAndersson, J., & Köper, I. (2016). Tethered and Polymer Supported Bilayer Lipid Membranes: Structure and Function. Membranes, 6(2), 30. https://doi.org/10.3390/membranes6020030