
Lipid-Based Catalysis Demonstrated by Bilayer-Enabled Ester Hydrolysis

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
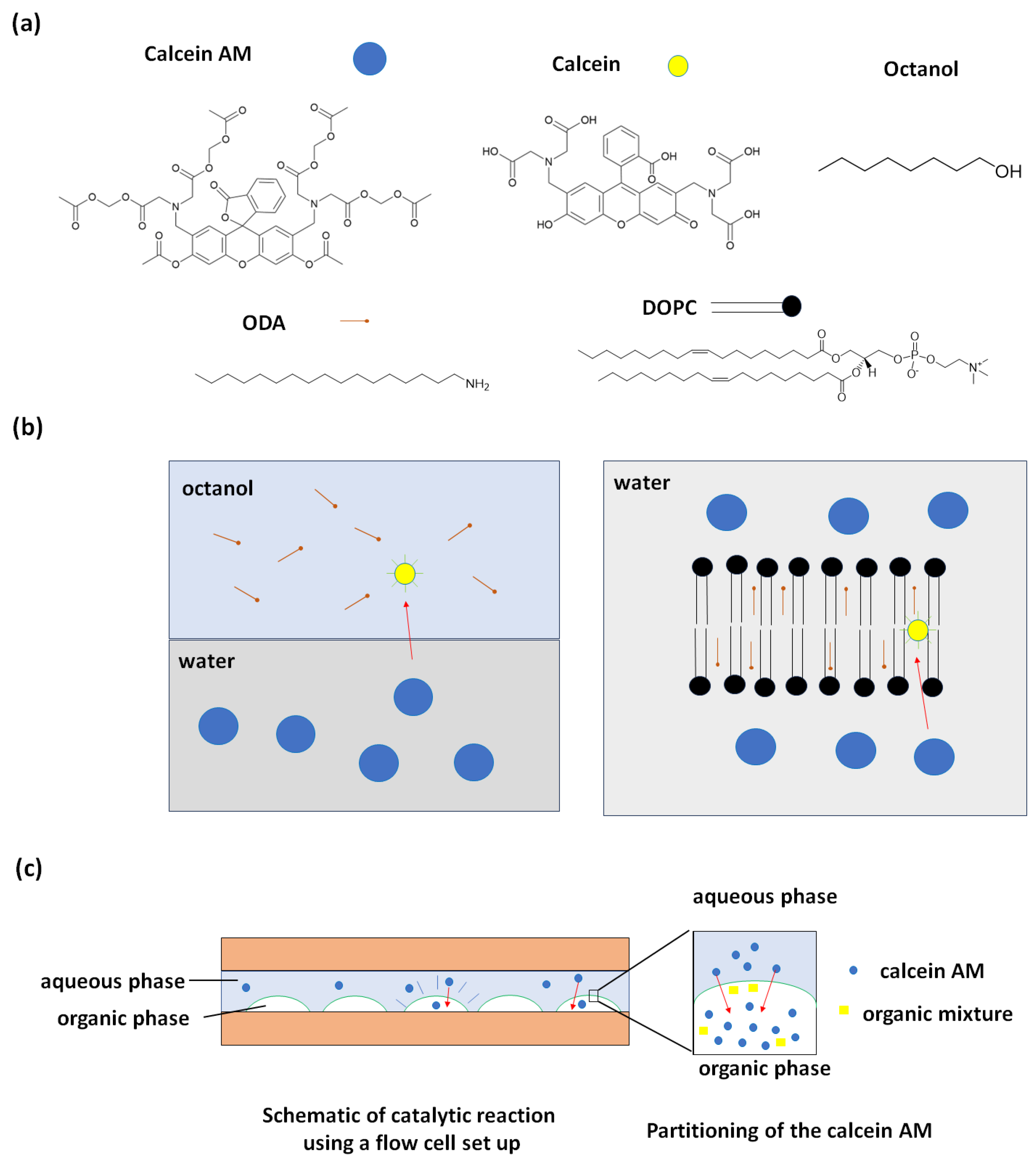
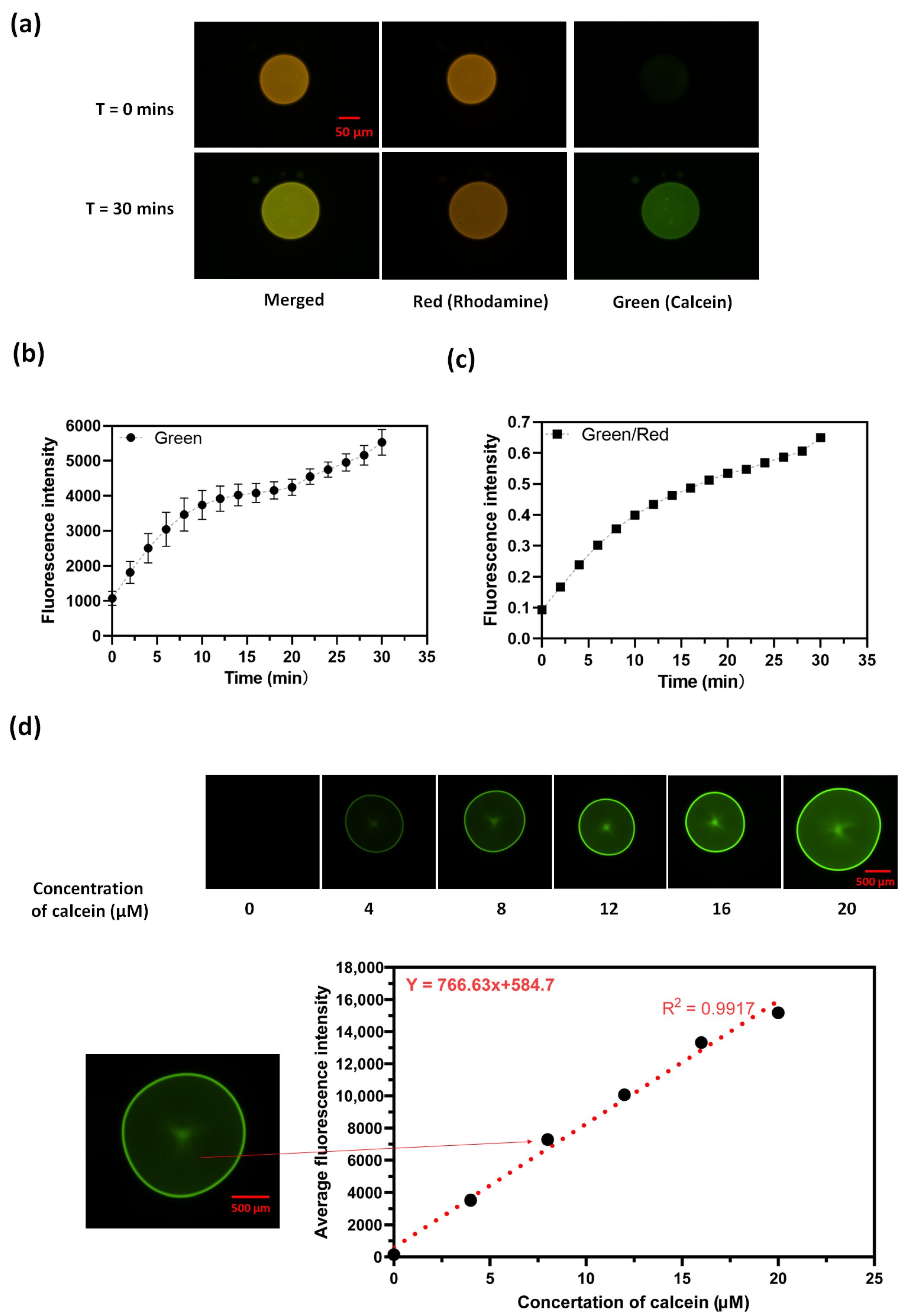
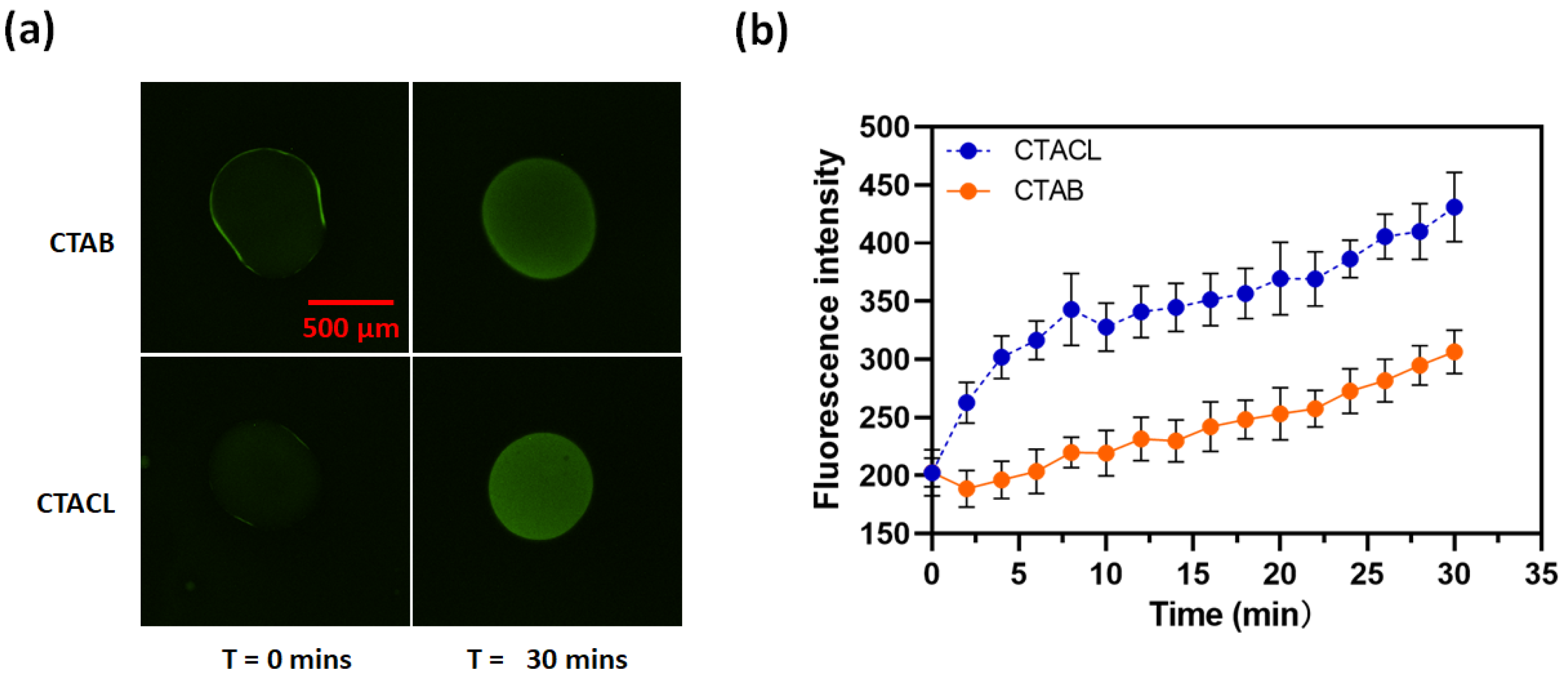
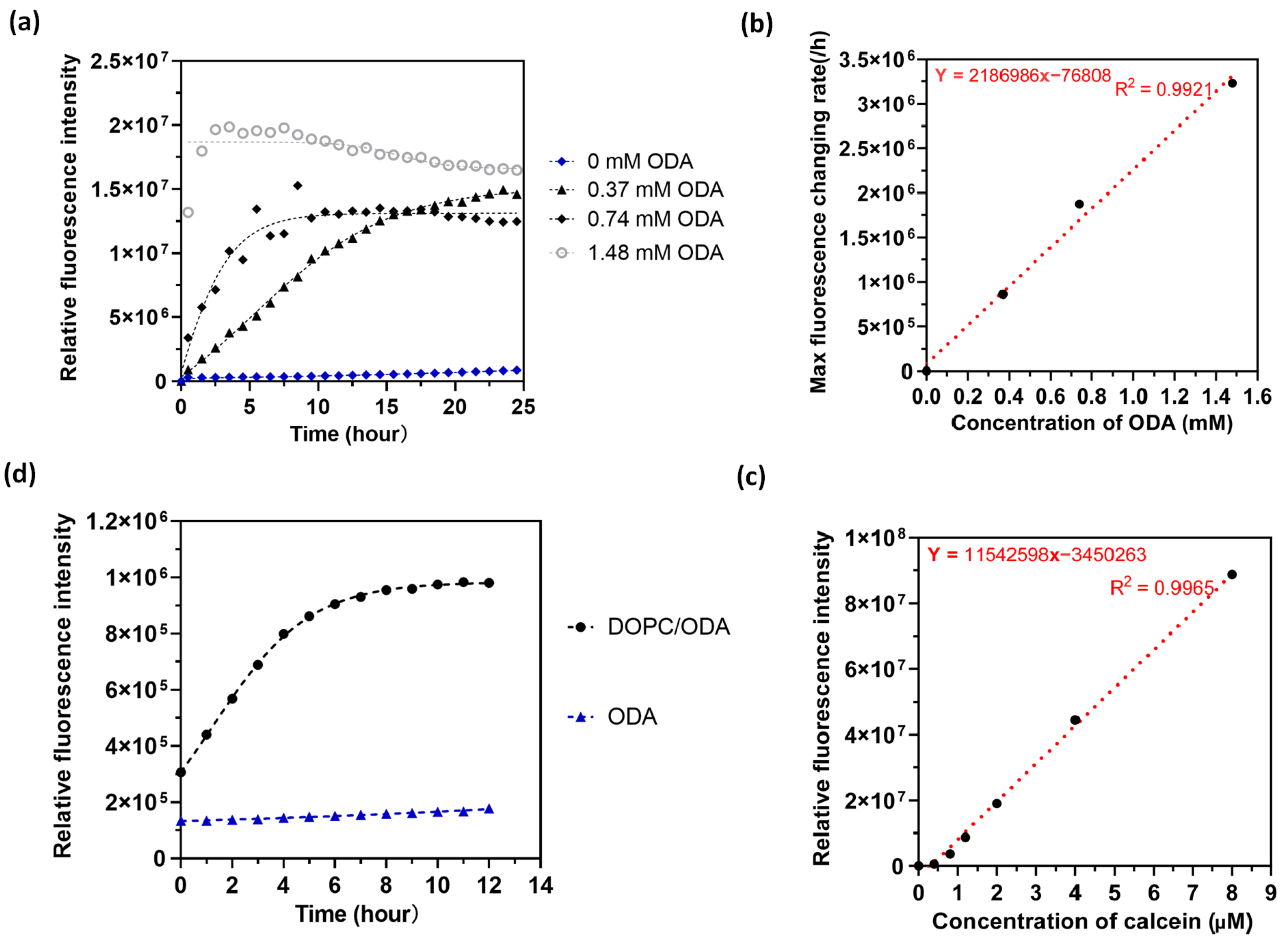
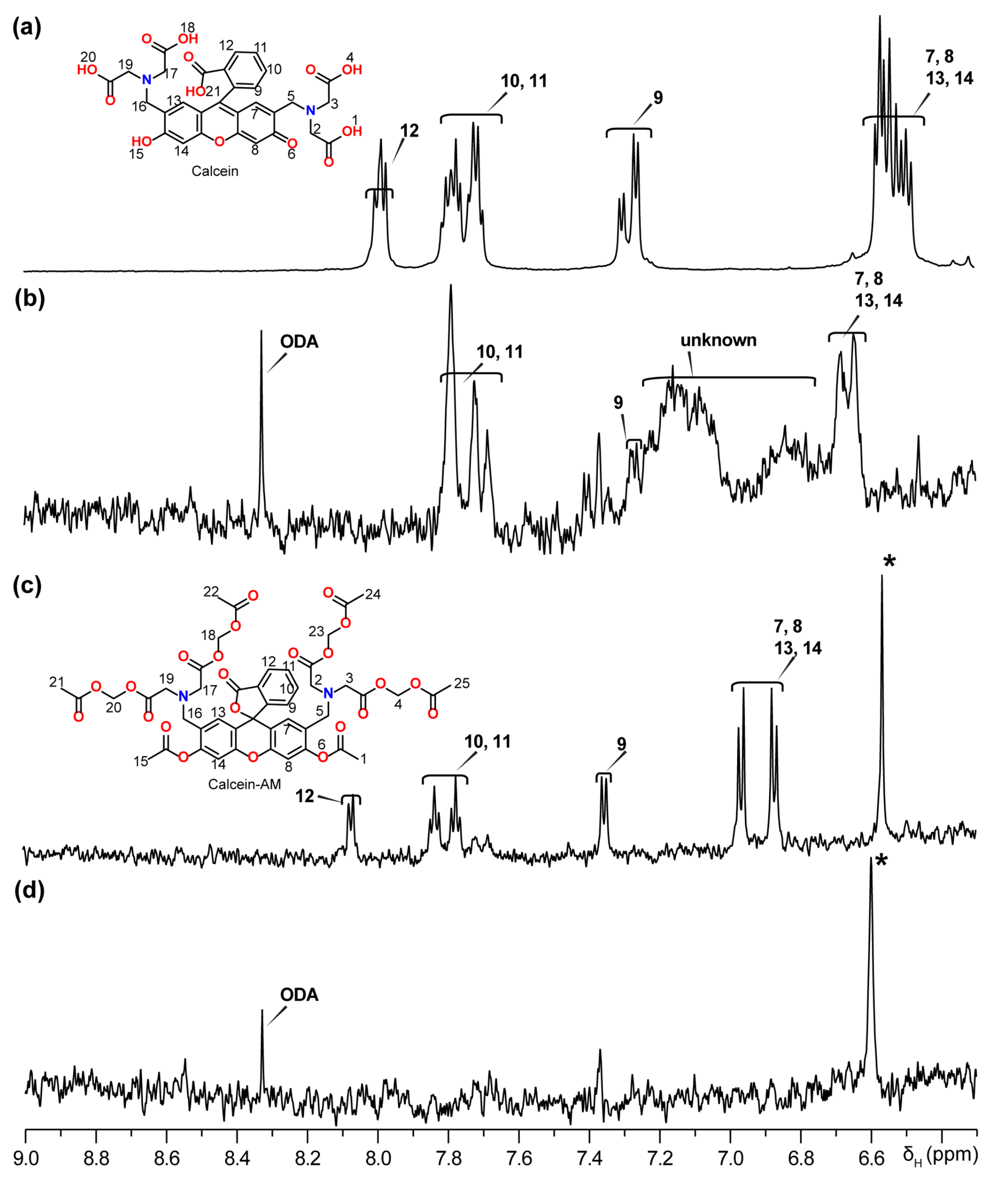
2. Results
3. Discussion
4. Conclusions
5. Methods
5.1. Materials
5.2. Vesicle Preparation
5.3. Fluorescence Microscopy
5.4. Fluorescence Characterization
5.5. Flow Cell Setup
5.6. NMR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| DOPC | 1,2-dioleoyl-sn-glycero-3-phosphocholine |
| Calcein AM | Calcein acetomethyl ester |
| ODA | Octadecylamine |
| CTAB | Cetyltrimethylammonium bromide |
| CTAC | Cetrimonium chloride |
| DMSO | Dimethyl sulfoxide |
References
- Supuran, C.T.; Winum, J.Y. Introduction to Zinc Enzymes as Drug Targets; Wiley: Hoboken, NJ, USA, 2009. [Google Scholar]
- Robertson, J.G. Mechanistic basis of enzyme-targeted drugs. Biochemistry 2005, 44, 5561–5571. [Google Scholar] [CrossRef]
- Copeland, R.A.; Harpel, M.R.; Tummino, P.J. Targeting enzyme inhibitors in drug discovery. Expert Opin. Ther. Targets 2007, 11, 967–978. [Google Scholar] [CrossRef]
- Dodson, G.; Wlodawer, A. Catalytic triads and their relatives. Trends Biochem. Sci. 1998, 23, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Schramm, V.L. Transition states and transition state analogue interactions with enzymes. Accounts Chem. Res. 2015, 48, 1032–1039. [Google Scholar] [CrossRef]
- Koshland, D.E., Jr. Application of a theory of enzyme specificity to protein synthesis. Proc. Natl. Acad. Sci. USA 1958, 44, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Koshland, D.E., Jr. The key–lock theory and the induced fit theory. Angew. Chem. Int. Ed. Engl. 1995, 33, 2375–2378. [Google Scholar] [CrossRef]
- Rebek, J., Jr. Molecular recognition with model systems. Angew. Chem. Int. Ed. Engl. 1990, 29, 245–255. [Google Scholar] [CrossRef]
- Bosshard, H.R. Molecular recognition by induced fit: How fit is the concept? Physiology 2001, 16, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Boehr, D.D.; Nussinov, R.; Wright, P.E. The role of dynamic conformational ensembles in biomolecular recognition. Nat. Chem. Biol. 2009, 5, 789–796. [Google Scholar] [CrossRef]
- Fendler, J.H. Interactions and reactions in reversed micellar systems. Accounts Chem. Res. 1976, 9, 153–161. [Google Scholar] [CrossRef]
- Cuccovia, I.M.; Quina, F.H.; Chaimovich, H. A remarkable enhancement of the rate of ester thiolysis by synthetic amphiphile vesicles. Tetrahedron 1982, 38, 917–920. [Google Scholar] [CrossRef]
- Bunton, C.A.; Fendler, E.J.; Sepulveda, G.L.; Yang, K.-U. Micellar-catalyzed hydrolysis of nitrophenyl phosphates. J. Am. Chem. Soc. 1968, 90, 5512–5518. [Google Scholar] [CrossRef]
- Jagdeep, K. Virdi, Ashish Dusunge, and Sachin Handa. Aqueous micelles as solvent, ligand, and reaction promoter in catalysis. JACS Au 2024, 4, 301–317. [Google Scholar]
- Ballester, P.; Scarso, A. Supramolecular Aspects in Catalysis. Front. Chem. 2019, 7, 174. [Google Scholar] [CrossRef]
- Wei, H.; Wang, E. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes. Chem. Soc. Rev. 2013, 42, 6060–6093. [Google Scholar] [CrossRef] [PubMed]
- Steed, J.W.; Atwood, J.L. Supramolecular Chemistry; John Wiley & Sons: New York, NY, USA, 2022. [Google Scholar]
- Raynal, M.; Ballester, P.; Vidal-Ferran, A.; Van Leeuwen, P.W. Supramolecular catalysis. Part 2: Artificial enzyme mimics. Chem. Soc. Rev. 2014, 43, 1734–1787. [Google Scholar] [CrossRef] [PubMed]
- Kahana, A.; Lancet, D. Self-reproducing catalytic micelles as nanoscopic protocell precursors. Nat. Rev. Chem. 2021, 5, 870–878. [Google Scholar] [CrossRef]
- Doherty, E.A.; Doudna, J.A. Ribozyme structures and mechanisms. Annu. Rev. Biochem. 2000, 69, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Lorsch, J.R. Ribozyme catalysis: Not different, just worse. Nat. Struct. Mol. Biol. 2005, 12, 395–402. [Google Scholar] [CrossRef]
- Chen, X.; Li, N.; Ellington, A.D. Ribozyme catalysis of metabolism in the RNA world. Chem. Biodivers. 2007, 4, 633–655. [Google Scholar] [CrossRef]
- Segré, D.; Ben-Eli, D.; Deamer, D.W.; Lancet, D. The lipid world. Orig. Life Evol. Biosph. 2001, 31, 119–145. [Google Scholar] [CrossRef] [PubMed]
- Lancet, D.; Segrè, D.; Kahana, A. Twenty years of “lipid world”: A fertile partnership with David Deamer. Life 2019, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Kust, P.R.; Rathman, J.F. Synthesis of surfactants by micellar autocatalysis: N, N-dimethyldodecylamine N-oxide. Langmuir 1995, 11, 3007–3012. [Google Scholar] [CrossRef]
- Hardy, M.D.; Yang, J.; Selimkhanov, J.; Cole, C.M.; Tsimring, L.S.; Devaraj, N.K. Self-reproducing catalyst drives repeated phospholipid synthesis and membrane growth. Proc. Natl. Acad. Sci. USA 2015, 112, 8187–8192. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Wick, R.; Fresta, M.; Mangone, A.; Luisi, P.L. Autopoietic self-reproduction of fatty acid vesicles. J. Am. Chem. Soc. 1994, 116, 11649–11654. [Google Scholar] [CrossRef]
- Wei, T.; Pan, T.; Peng, X.; Zhang, M.; Guo, R.; Guo, Y.; Mei, X.; Zhang, Y.; Qi, J.; Dong, F.; et al. Janus liposozyme for the modulation of redox and immune homeostasis in infected diabetic wounds. Nat. Nanotechnol. 2024; in press. [Google Scholar]
- Schuch, R.; Mukherjee, K.D. Interesterification of lipids using an immobilized sn-1, 3-specific triacylglycerol lipase. J. Agric. Food Chem. 1987, 35, 1005–1008. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R. Lipase from Thermomyces lanuginosus: Uses and prospects as an industrial biocatalyst. J. Mol. Catal. B Enzym. 2010, 62, 197–212. [Google Scholar] [CrossRef]
- Bell, T.N.; Feng, K.; Calvin, G.; Van Winkle, D.H.; Lenhert, S. Organic composomes as supramolecular aptamers. ACS Omega 2020, 5, 27393–27400. [Google Scholar] [CrossRef]
- Zhou, H.; Shiel, E.; Bell, T.; Lin, S.; Lenhert, S. Kinetic Mechanism of Surfactant-Based Molecular Recognition: Selective Permeability across an Oil–Water Interface Regulated by Supramolecular Aggregates. J. Phys. Chem. B 2023, 127, 10201–10214. [Google Scholar] [CrossRef]
- Liu, S.; Wei, A.T.; Wang, H.; Van Winkle, D.; Lenhert, S. Combinatorial mixtures of organic solutes for improved liquid/liquid extraction of ions. Soft Matter 2023, 19, 6903–6910. [Google Scholar] [CrossRef]
- Lowry, T.W.; Kusi-Appiah, A.E.; Fadool, D.A.; Lenhert, S. Odor discrimination by lipid membranes. Membranes 2023, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Yotsumoto, M.; Matsuo, M.; Kitahata, H.; Nakanishi, S.; Denda, M.; Nagayama, M.; Nakata, S. Phospholipid Molecular Layer that Enhances Distinction of Odors Based on Artificial Sniffing. ACS Sensors 2023, 8, 4494–4503. [Google Scholar] [CrossRef] [PubMed]
- Rafeeq, H.; Hussain, A.; Shabbir, S.; Ali, S.; Bilal, M.; Sher, F.; Iqbal, H.M. Esterases as emerging biocatalysts: Mechanistic insights, genomic and metagenomic, immobilization, and biotechnological applications. Biotechnol. Appl. Biochem. 2022, 69, 2176–2194. [Google Scholar] [CrossRef] [PubMed]
- Barzkar, N.; Sohail, M.; Tamadoni Jahromi, S.; Gozari, M.; Poormozaffar, S.; Nahavandi, R.; Hafezieh, M. Marine bacterial esterases: Emerging biocatalysts for industrial applications. Appl. Biochem. Biotechnol. 2021, 193, 1187–1214. [Google Scholar] [CrossRef] [PubMed]
- Bratosin, D.; Mitrofan, L.; Palii, C.; Estaquier, J.; Montreuil, J. Novel fluorescence assay using calcein-AM for the determination of human erythrocyte viability and aging. Cytom. Part A J. Int. Soc. Anal. Cytol. 2005, 66, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Bozyczko-Coyne, D.; McKenna, B.W.; Connors, T.J.; Neff, N.T. A rapid fluorometric assay to measure neuronal survival in vitro. J. Neurosci. Methods 1993, 50, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Uggeri, J.; Gatti, R.; Belletti, S.; Scandroglio, R.; Corradini, R.; Rotoli, B.M.; Orlandini, G. Calcein-AM is a detector of intracellular oxidative activity. Histochem. Cell Biol. 2004, 122, 499–505. [Google Scholar] [CrossRef]
- Klonis, N.; Sawyer, W.H. Spectral properties of the prototropic forms of fluorescein in aqueous solution. J. Fluoresc. 1996, 6, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Ristanović, Z.; Hofmann, J.P.; Cremer, G.D.; Kubarev, A.V.; Rohnke, M.; Meirer, F.; Hofkens, J.; Roeffaers, M.B.; Weckhuysen, B.M. Quantitative 3D fluorescence imaging of single catalytic turnovers reveals spatiotemporal gradients in reactivity of zeolite H-ZSM-5 crystals upon steaming. J. Am. Chem. Soc. 2015, 137, 6559–6568. [Google Scholar] [CrossRef]
- Boudart, M. Turnover rates in heterogeneous catalysis. Chem. Rev. 1995, 95, 661–666. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, W.; Liu, G.; Panda, D.; Chen, P. Size-dependent catalytic activity and dynamics of gold nanoparticles at the single-molecule level. J. Am. Chem. Soc. 2010, 132, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Costentin, C.; Drouet, S.; Robert, M.; Saveant, J.M. Turnover numbers, turnover frequencies, and overpotential in molecular catalysis of electrochemical reactions. Cyclic voltammetry and preparative-scale electrolysis. J. Am. Chem. Soc. 2012, 134, 11235–11242. [Google Scholar] [CrossRef]
- Pegis, M.L.; McKeown, B.A.; Kumar, N.; Lang, K.; Wasylenko, D.J.; Zhang, X.P.; Raugei, S.; Mayer, J.M. Homogenous electrocatalytic oxygen reduction rates correlate with reaction overpotential in acidic organic solutions. ACS Cent. Sci. 2016, 2, 850–856. [Google Scholar] [CrossRef]
- Nafday, O.A.; Lenhert, S. High-throughput optical quality control of lipid multilayers fabricated by dip-pen nanolithography. Nanotechnology 2011, 22, 225301. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; MacMillan, D.W. Synergistic catalysis: A powerful synthetic strategy for new reaction development. Chem. Sci. 2012, 3, 633–658. [Google Scholar] [CrossRef] [PubMed]
- Okuom, M.O.; Wilson, M.V.; Jackson, A.; Holmes, A.E. Intermolecular Interactions between Eosin Y and Caffeine Using 1 H-NMR Spectroscopy. Int. J. Spectrosc. 2013, 2013, 245376. [Google Scholar] [CrossRef]
- Herriott, A.W.; Picker, D. Phase transfer catalysis. Evaluation of catalysis. J. Am. Chem. Soc. 1975, 97, 2345–2349. [Google Scholar] [CrossRef]
- Starks, C.M.; Halper, M. Phase-Transfer Catalysis: Fundamentals, Applications, and Industrial Perspectives; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Ooi, T.; Maruoka, K. Recent advances in asymmetric phase-transfer catalysis. Angew. Chem. Int. Ed. 2007, 46, 4222–4266. [Google Scholar] [CrossRef] [PubMed]
- Godha, A.K.; Thiruvengadam, J.; Abhilash, V.; Balgi, P.; Narayanareddy, A.; Vignesh, K.; Gadakh, A.V.; Sathiyanarayanan, A.; Ganesh, S. Environmentally benign nucleophilic substitution reaction of arylalkyl halides in water using CTAB as the inverse phase transfer catalyst. New J. Chem. 2019, 43, 16041–16045. [Google Scholar] [CrossRef]
- Kumar, A.; Joshi, H.; Pasricha, R.; Mandale, A.; Sastry, M. Phase transfer of silver nanoparticles from aqueous to organic solutions using fatty amine molecules. J. Colloid Interface Sci. 2003, 264, 396–401. [Google Scholar] [CrossRef]
- Wikander, K.; Petit, C.; Holmberg, K.; Pileni, M.P. Size control and growth process of alkylamine-stabilized platinum nanocrystals: A comparison between the phase transfer and reverse micelles methods. Langmuir 2006, 22, 4863–4868. [Google Scholar] [CrossRef] [PubMed]
- Starks, C.M. Phase-transfer catalysis. I. Heterogeneous reactions involving anion transfer by quaternary ammonium and phosphonium salts. J. Am. Chem. Soc. 1971, 93, 195–199. [Google Scholar] [CrossRef]
- Kumar, A.; Joshi, H.M.; Mandale, A.B.; Srivastava, R.; Adyanthaya, S.D.; Pasricha, R.; Sastry, M. Phase transfer of platinum nanoparticles from aqueous to organic solutions using fatty amine molecules. J. Chem. Sci. 2004, 116, 293–300. [Google Scholar] [CrossRef]
- Xu, D.Q.; Pan, Z.W. Phase-transfer catalysis of a new cationic gemini surfactant with ester groups for nucleophilic substitution reaction. Chin. Chem. Lett. 2014, 25, 1169–1173. [Google Scholar] [CrossRef]
- Tan, S.; Zhang, X.; Kong, W.; Yang, X.L.; Molnár, G.; Vondráková, Z.; Filepová, R.; Petrášek, J.; Friml, J.; Xue, H.W. The lipid code-dependent phosphoswitch PDK1–D6PK activates PIN-mediated auxin efflux in Arabidopsis. Nat. Plants 2020, 6, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Turunen, M.; Olsson, J.; Dallner, G. Metabolism and function of coenzyme Q. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2004, 1660, 171–199. [Google Scholar] [CrossRef] [PubMed]
- Stafford, D. The vitamin K cycle. J. Thromb. Haemost. 2005, 3, 1873–1878. [Google Scholar] [CrossRef]
- Lokugamage, M.P.; Sago, C.D.; Gan, Z.; Krupczak, B.R.; Dahlman, J.E. Constrained nanoparticles deliver siRNA and sgRNA to T cells in vivo without targeting ligands. Adv. Mater. 2019, 31, 1902251. [Google Scholar] [CrossRef]
- Lian, T.; Ho, R.J. Trends and developments in liposome drug delivery systems. J. Pharm. Sci. 2001, 90, 667–680. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Kumar, K.; Bell, T.; Ramamoorthy, A.; Van Winkle, D.; Lenhert, S. Lipid-Based Catalysis Demonstrated by Bilayer-Enabled Ester Hydrolysis. Membranes 2024, 14, 168. https://doi.org/10.3390/membranes14080168
Liu S, Kumar K, Bell T, Ramamoorthy A, Van Winkle D, Lenhert S. Lipid-Based Catalysis Demonstrated by Bilayer-Enabled Ester Hydrolysis. Membranes. 2024; 14(8):168. https://doi.org/10.3390/membranes14080168
Chicago/Turabian StyleLiu, Shu, Kiran Kumar, Tracey Bell, Ayyalusamy Ramamoorthy, David Van Winkle, and Steven Lenhert. 2024. "Lipid-Based Catalysis Demonstrated by Bilayer-Enabled Ester Hydrolysis" Membranes 14, no. 8: 168. https://doi.org/10.3390/membranes14080168
APA StyleLiu, S., Kumar, K., Bell, T., Ramamoorthy, A., Van Winkle, D., & Lenhert, S. (2024). Lipid-Based Catalysis Demonstrated by Bilayer-Enabled Ester Hydrolysis. Membranes, 14(8), 168. https://doi.org/10.3390/membranes14080168