
Interactions of Galloylated Polyphenols with a Simple Gram-Negative Bacterial Membrane Lipid Model





Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lipid Vesicle Preparation
2.3. Differential Scanning Calorimetry
3. Results and Discussion
3.1. DSC Thermograms of Individual Lipid Components
3.2. Interactions of (-)-EGCg with Ideally and Non-Ideally Mixed Lipid Vesicles
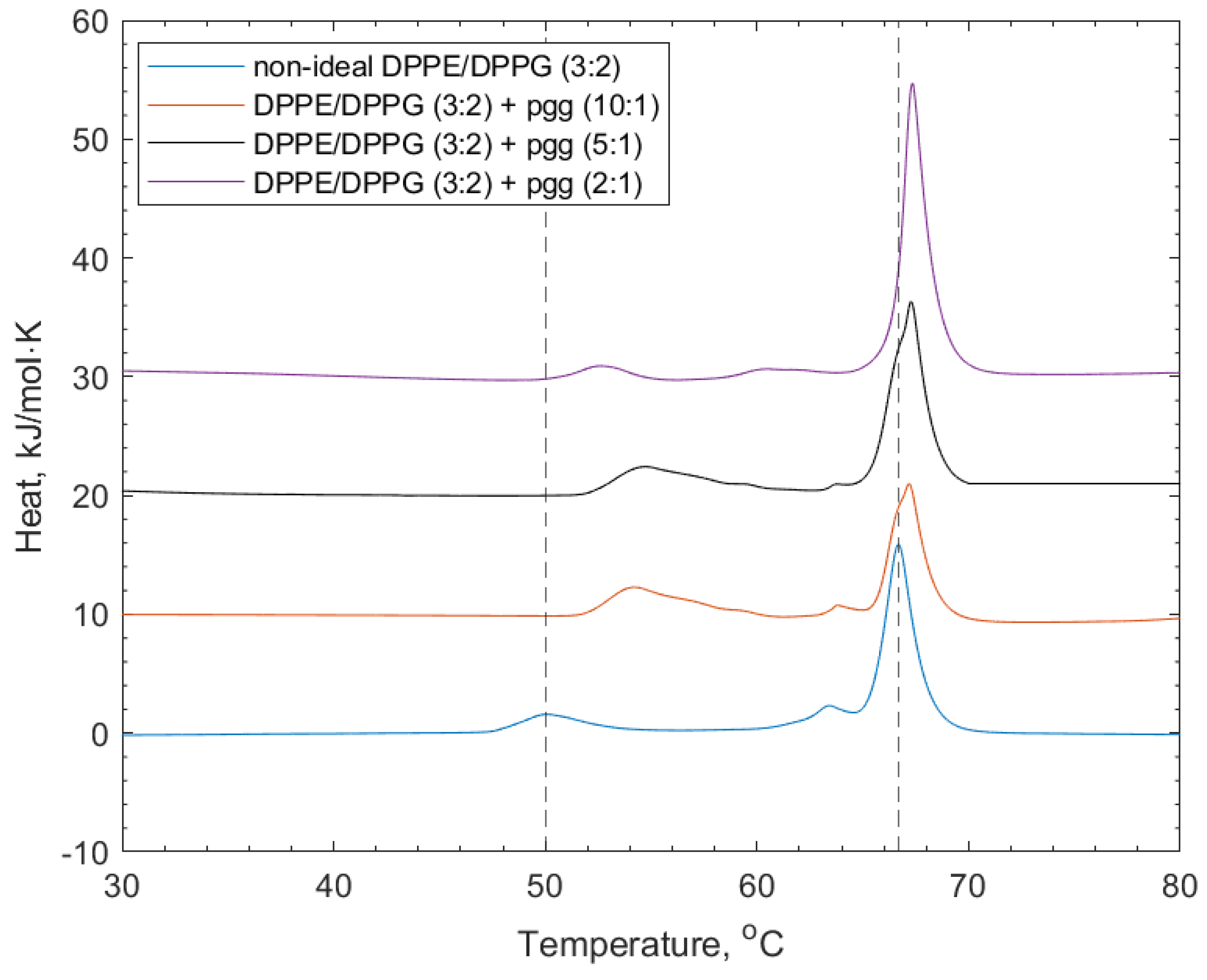
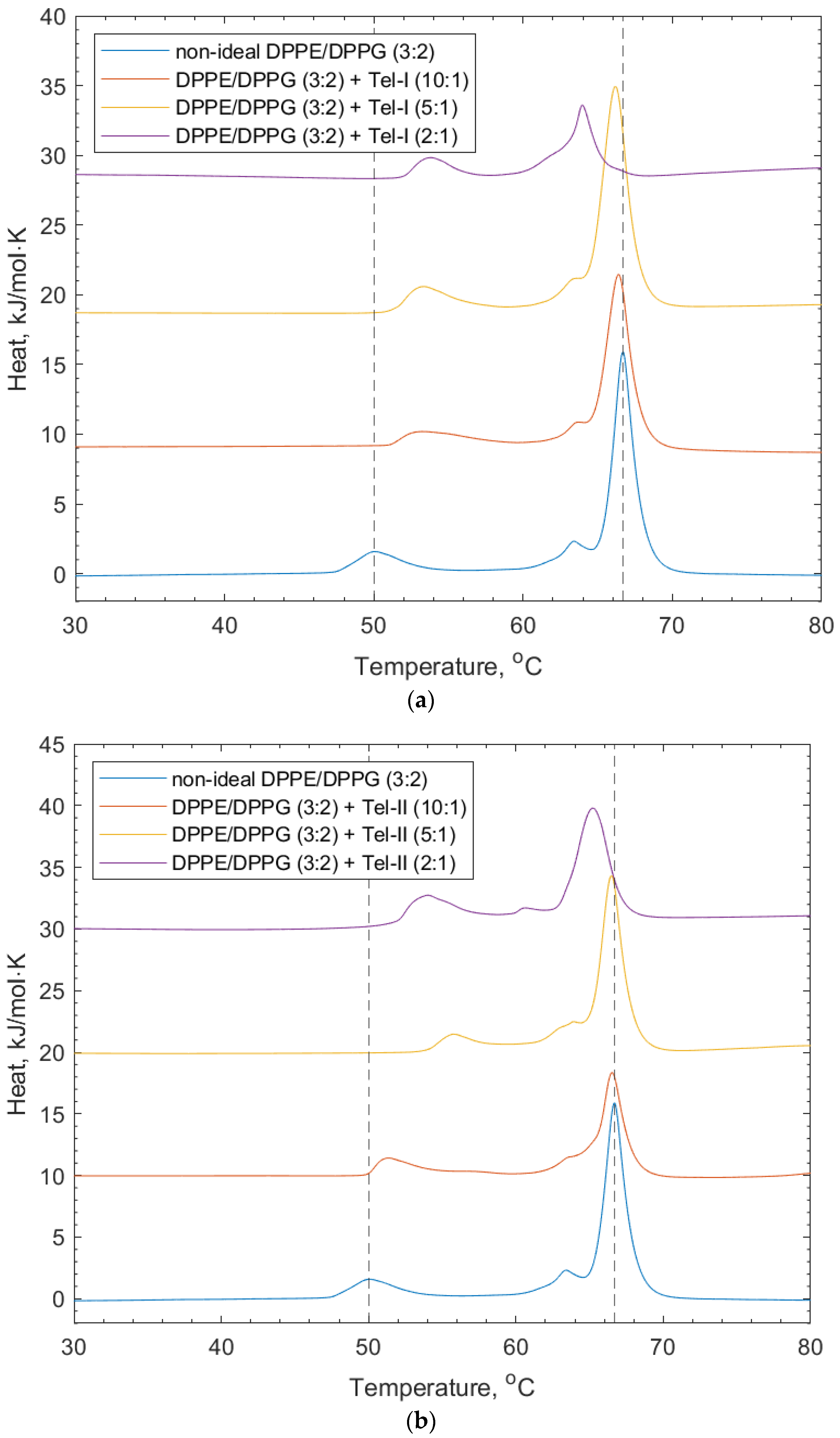
3.3. Interactions of PGG, Tel-I, and Tel-II with Non-Ideally Mixed Lipid Vesicles
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Li, A.N.; Li, S.; Zhang, Y.J.; Xu, X.R.; Chen, Y.M.; Bin Li, H. Resources and biological activities of natural polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef] [PubMed]
- Hoste, H.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Mueller-Harvey, I.; Sotiraki, S.; Louvandini, H.; Thamsborg, S.M.; Terrill, T.H. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 2015, 212, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Encinar, J.A.; Rodríguez-Díaz, J.C.; Micol, V. Antimicrobial capacity of plant polyphenols against Gram-positive Bacteria: A comprehensive review. Curr. Med. Chem. 2020, 27, 2576–2606. [Google Scholar] [CrossRef] [PubMed]
- Puljula, E.; Walton, G.; Woodward, M.J.; Karonen, M. Antimicrobial activities of Ellagitannins against Clostridiales perfringens, Escherichia coli, Lactobacillus plantarum and Staphylococcus aureus. Molecules 2020, 25, 3714. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Hashimoto, T.; Kajiya, K.; Kumazawa, S. Affinity of polyphenols for lipid bilayers. Biofactors 2000, 13, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Hung, W.C.; Chen, F.Y.; Lee, C.C.; Huang, H.W. Interaction of tea catechin (-)-epigallocatechin gallate with lipid bilayers. Biophys. J. 2009, 96, 1026–1035. [Google Scholar] [CrossRef]
- Phan, H.T.T.; Yoda, T.; Chahal, B.; Morita, M.; Takagi, M.; Vestergaard, M.C. Structure-dependent interactions of polyphenols with a biomimetic membrane system. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2670–2677. [Google Scholar] [CrossRef]
- Virtanen, V.; Räikkönen, S.; Puljula, E.; Karonen, M. Ellagitannin–Lipid interaction by HR-MAS NMR Spectroscopy. Molecules 2021, 26, 373. [Google Scholar] [CrossRef]
- Virtanen, V.; Green, R.J.; Karonen, M. Interactions between Hydrolysable Tannins and Lipid Vesicles from Escherichia coli with isothermal titration calorimetry. Molecules 2022, 27, 3204. [Google Scholar] [CrossRef]
- Altunayar-Unsalan, C.; Unsalan, O.; Mavromoustakos, T. Insights into molecular mechanism of action of citrus flavonoids hesperidin and naringin on lipid bilayers using spectroscopic, calorimetric, microscopic and theoretical studies. J. Mol. Liq. 2022, 347, 118411. [Google Scholar] [CrossRef]
- Sirk, T.W.; Brown, E.F.; Sum, A.K.; Friedman, M. Molecular dynamics study on the biophysical interactions of seven green tea catechins with lipid bilayers of cell membranes. J. Agric. Food Chem. 2008, 56, 7750–7758. [Google Scholar] [CrossRef]
- Karonen, M. Insights into polyphenol–lipid interactions: Chemical methods, molecular aspects and their effects on membrane structures. Plants 2022, 11, 1809. [Google Scholar] [CrossRef] [PubMed]
- Nassarawa, S.S.; Nayik, G.A.; Gupta, S.D.; Areche, F.O.; Jagdale, Y.D.; Ansari, M.J. Chemical aspects of polyphenol-protein interactions and the antibacterial activity. Crit. Rev. Food Sci. Nutr. 2023, 63, 9482–9505. [Google Scholar] [CrossRef] [PubMed]
- Caturla, N.; Vera-Samper, E.; Villalaín, J.; Mateo, C.R.; Micol, V. The relationship between the antioxidant and the antibacterial properties of galloylated catechins and the structure of phospholipid model membranes. Free Radic. Biol. Med. 2003, 34, 648–662. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; de Freitas, V. When polyphenols meet lipids: Challenges in membrane biophysics and opportunities in epithelial lipidomics. Food Chem. 2020, 333, 127509. [Google Scholar] [CrossRef] [PubMed]
- Kajiya, K.; Kumazawa, S.; Naito, A.; Nakayama, T. Solid-state NMR analysis of the orientation and dynamics of epigallocatechin gallate, a green tea polyphenol, incorporated into lipid bilayers. Magn. Reson. Chem. 2008, 46, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Clifton, L.A.; Hall, S.C.L.; Mahmoudi, N.; Knowles, T.J.; Heinrich, F.; Lakey, J.H. Structural investigations of protein-lipid complexes using neutron scattering. Methods Mol. Biol. 2019, 2003, 201–251. [Google Scholar] [CrossRef] [PubMed]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed]
- Coones, R.T.; Green, R.J.; Frazier, R.A. Investigating lipid headgroup composition within epithelial membranes: A systematic review. Soft Matter 2021, 17, 6773–6786. [Google Scholar] [CrossRef]
- Simons, K.; Vaz, W.L.C. Model Ssystems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef] [PubMed]
- Karonen, M.; Virtanen, V. Partition coefficients of hydrolysable tannins. Molecules 2020, 25, 3691. [Google Scholar] [CrossRef]
- Salminen, J.P.; Lempa, K. Effects of hydrolysable tannins on a herbivorous insect: Fate of individual tannins in insect digestive tract. Chemoecology 2002, 12, 203–211. [Google Scholar] [CrossRef]
- Raudino, A.; Sarpietro, M.G.; Pannuzzo, M. Differential scanning calorimetry (DSC): Theoretical fundamentals. In Drug-Biomembrane Interaction Studies: The Application of Calorimetric Techniques; Pignatello, R., Ed.; Woodhead Publishing: Cambridge, UK, 2013; pp. 127–168. [Google Scholar] [CrossRef]
- Lohner, K.; Prenner, E.J. Differential scanning calorimetry and X-ray diffraction studies of the specificity of the interaction of antimicrobial peptides with membrane-mimetic systems. Biochim. Biophys. Acta Biomembr. 1999, 1462, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Pabst, G.; Kučerka, N.; Nieh, M.P.; Rheinstädter, M.C.; Katsaras, J. Applications of neutron and X-ray scattering to the study of biologically relevant model membranes. Chem. Phys. Lipids. 2010, 163, 460–479. [Google Scholar] [CrossRef]
- Arouri, A.; Dathe, M.; Blume, A. Peptide induced demixing in PG/PE lipid mixtures: A mechanism for the specificity of antimicrobial peptides towards bacterial membranes? Biochim. Biophys. Acta Biomembr. 2009, 1788, 650–659. [Google Scholar] [CrossRef]
- Schwieger, C.; Blume, A. Interaction of poly(L-lysines) with negatively charged membranes: An FT-IR and DSC study. Eur. Biophys. J. 2007, 36, 437–450. [Google Scholar] [CrossRef]
- Colina, J.R.; Suwalsky, M.; Manrique-Moreno, M.; Petit, K.; Aguilar, L.F.; Jemiola-Rzeminska, M.; Strzalka, K. Protective effect of epigallocatechin gallate on human erythrocytes. Colloids Surf. B Biointerfaces 2019, 173, 742–750. [Google Scholar] [CrossRef]
- Nie, R.; Dang, M.; Ge, Z.; Huo, Y.; Yu, B.; Tang, S. Influence of the gallate moiety on the interactions between green tea polyphenols and lipid membranes elucidated by molecular dynamics simulations. Biophys. Chem. 2021, 274, 106592. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Ito, H.; Chevyreva, V.; Makky, A.; Kaufmann, S.; Okano, K.; Kobayashi, N.; Suganuma, M.; Nakabayashi, S.; Yoshikawa, H.Y.; et al. Adsorption of galloyl catechin aggregates significantly modulates membrane mechanics in the absence of biochemical cues. Phys. Chem. Chem. Phys. 2017, 19, 19937–19947. [Google Scholar] [CrossRef]
- Tanaka, T.; Zhang, H.; Jiang, Z.-H.; Kouno, I. Relationship between hydrophobicity and structure of Hydrolyzable Tannins, and association of Tannins with crude drug constituents in aqueous solution. Chem. Pharm. Bull. 1997, 12, 1891–1897. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Meng, N.; Chen, S.W.; Li, Y. Study of acetylated EGCG synthesis by enzymatic transesterification in organic media. Arab. J. Chem. 2020, 13, 8824–8834. [Google Scholar] [CrossRef]
- Zhu, S.; Li, Y.; Li, Z.; Ma, C.; Lou, Z.; Yokoyama, W.; Wang, H. Lipase-catalyzed synthesis of acetylated EGCG and antioxidant properties of the acetylated derivatives. Food Res. Int. 2014, 56, 279–286. [Google Scholar] [CrossRef]
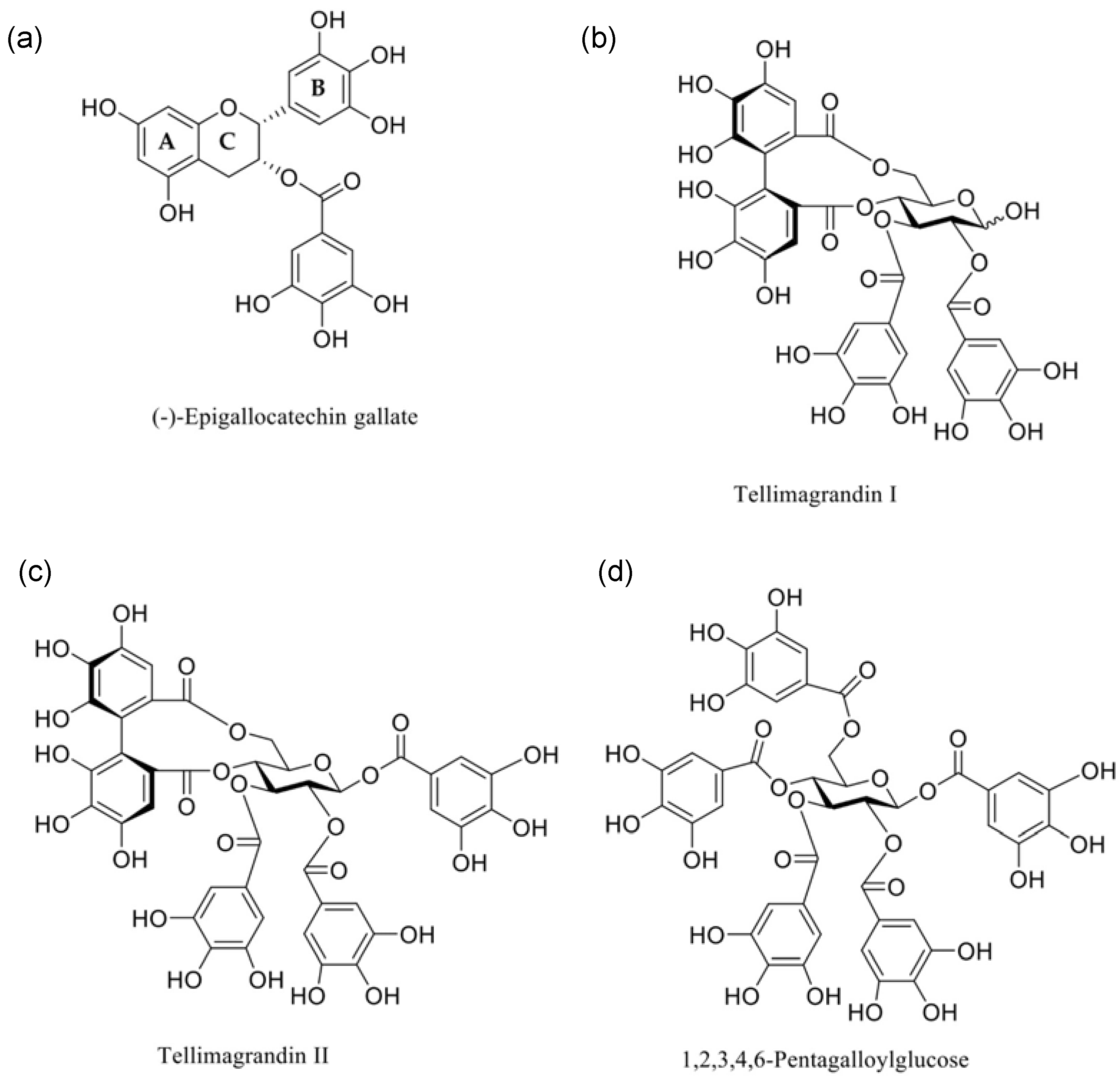



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Vesicle System | Peak 1/°C | Std. Dev. (n = 3) | Peak 2/°C | Std. Dev. (n = 3) |
|---|---|---|---|---|
| Ideal DPPE/DPPG (3:2) | 60.51 | 0.17 | - | - |
| + (-)-EGCg (10:1) * | 62.10 | 0.07 | - | - |
| + (-)-EGCg (5:1) | 63.99 | 0.15 | - | - |
| + (-)-EGCg (2:1) | 50.46 | 0.23 | 66.12 | 0.06 |
| Non-ideal DPPE/DPPG (3:2) | 51.60 | 0.60 | 67.00 | 0.10 |
| + (-)-EGCg (10:1) | 52.00 | 0.50 | 66.20 | 0.10 |
| + (-)-EGCg (5:1) | - | - | 66.40 | 0.10 |
| Lipid Vesicle System | Peak 1/°C | Std. Dev. (n = 3) | Peak 2/°C | Std. Dev. (n = 3) |
|---|---|---|---|---|
| Non-ideal DPPE/DPPG (3:2) | 51.60 | 0.60 | 67.00 | 0.10 |
| + PGG (10:1) | 57.00 | 1.20 | 65.70 | 0.10 |
| + PGG (5:1) | 55.90 | 0.80 | 66.20 | 0.90 |
| + PGG (2:1) | 52.90 | 0.20 | 67.40 | 1.10 |
| + Tel-I (10:1) | 50.60 | 0.10 | 66.50 | 0.10 |
| + Tel-I (5:1) | 52.00 | 0.80 | 66.10 | 0.10 |
| + Tel-I (2:1) | 53.25 | 0.73 | 65.26 | 1.36 |
| + Tel-II (10:1) | 53.00 | 0.80 | 66.40 | 0.10 |
| + Tel-II (5:1) | 54.60 | 1.30 | 66.50 | 0.10 |
| + Tel-II (2:1) | 53.30 | 0.40 | 65.40 | 0.20 |
| Polyphenol | DPPG Peak Shift | Mean DPPG Peak Temp Shift a/°C | DPPE Peak Shift | Mean DPPE Peak Temp Shift a/°C | Number of Free Galloyl Groups | Nominal Mass | LogP |
|---|---|---|---|---|---|---|---|
| (-)-EGCg | - | - | Decrease | −0.6 | 1 | 442 Da | 1.89 b |
| Tel-I | Increase | +1.7 | Decrease | −1.7 | 2 | 786 Da | −0.44 c |
| Tel-II | Increase | +1.7 | Decrease | −1.6 | 3 | 938 Da | 0.86 c |
| PGG | Increase | +1.3 | Increase | +0.4 | 5 | 940 Da | 1.49 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coones, R.T.; Karonen, M.; Green, R.J.; Frazier, R. Interactions of Galloylated Polyphenols with a Simple Gram-Negative Bacterial Membrane Lipid Model. Membranes 2024, 14, 47. https://doi.org/10.3390/membranes14020047
Coones RT, Karonen M, Green RJ, Frazier R. Interactions of Galloylated Polyphenols with a Simple Gram-Negative Bacterial Membrane Lipid Model. Membranes. 2024; 14(2):47. https://doi.org/10.3390/membranes14020047
Chicago/Turabian StyleCoones, Ryan T., Maarit Karonen, Rebecca J. Green, and Richard Frazier. 2024. "Interactions of Galloylated Polyphenols with a Simple Gram-Negative Bacterial Membrane Lipid Model" Membranes 14, no. 2: 47. https://doi.org/10.3390/membranes14020047
APA StyleCoones, R. T., Karonen, M., Green, R. J., & Frazier, R. (2024). Interactions of Galloylated Polyphenols with a Simple Gram-Negative Bacterial Membrane Lipid Model. Membranes, 14(2), 47. https://doi.org/10.3390/membranes14020047