
Interaction of Hyaluronan Acid with Some Proteins in Aqueous Solution as Studied by NMR



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
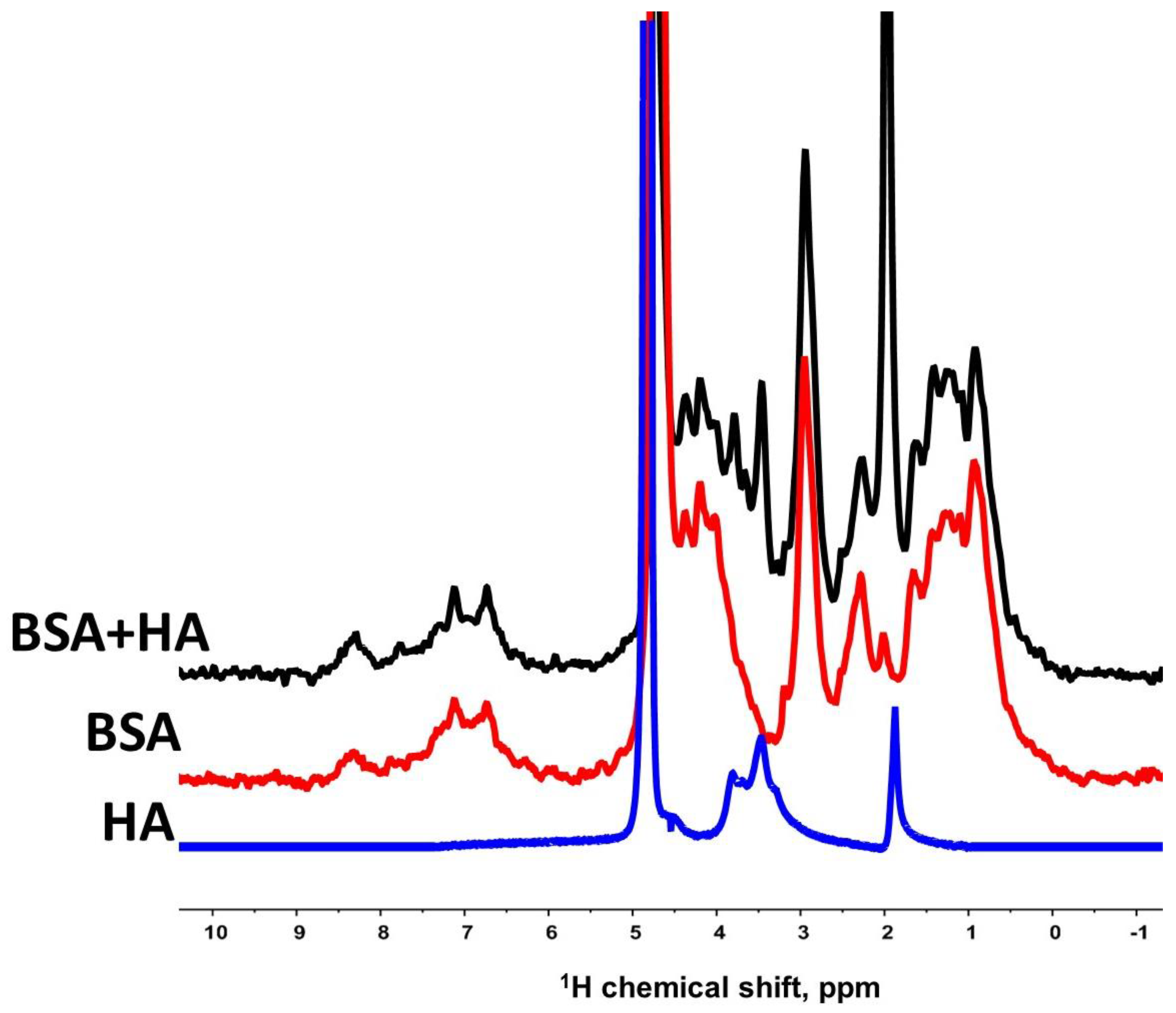
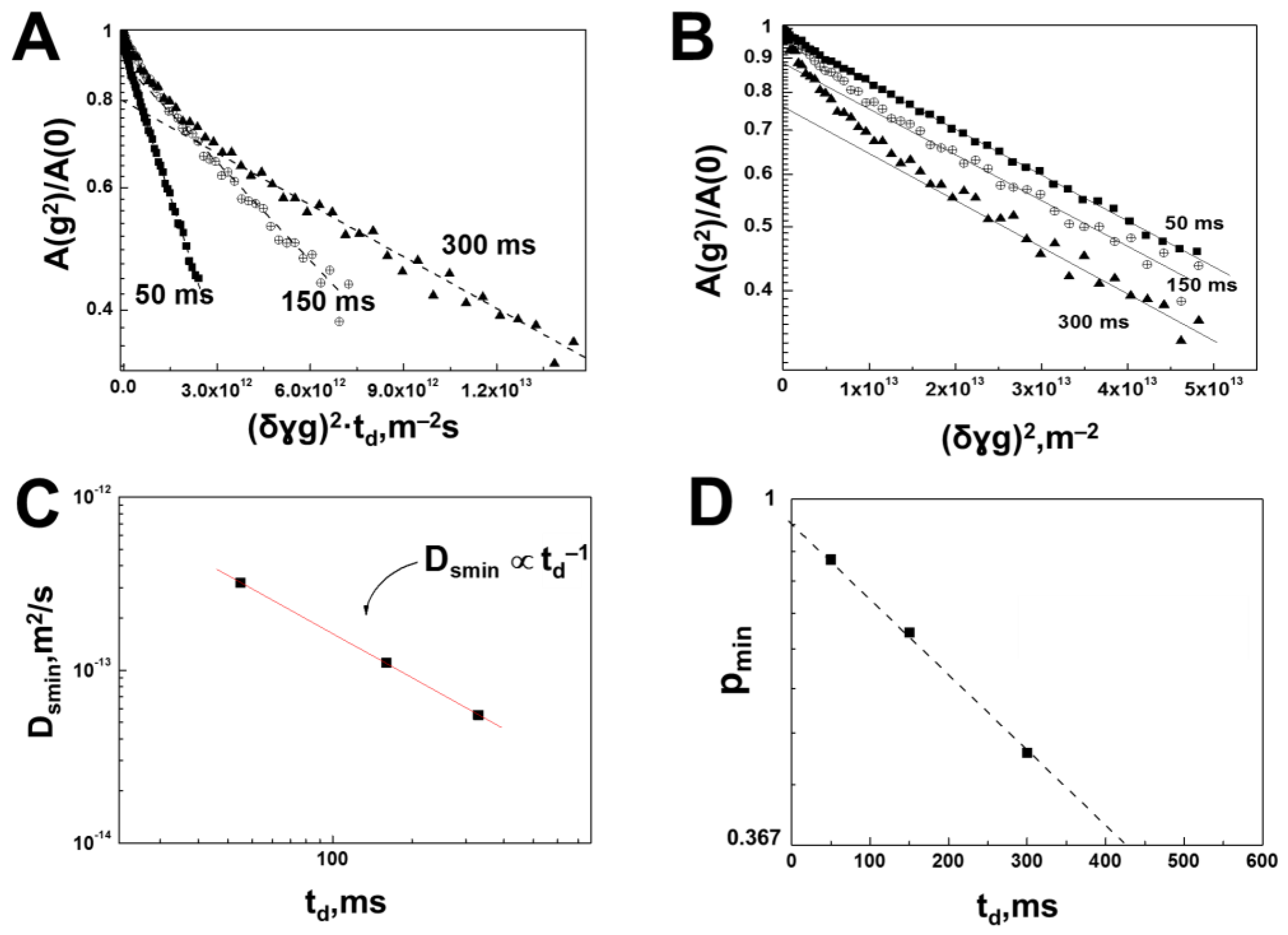
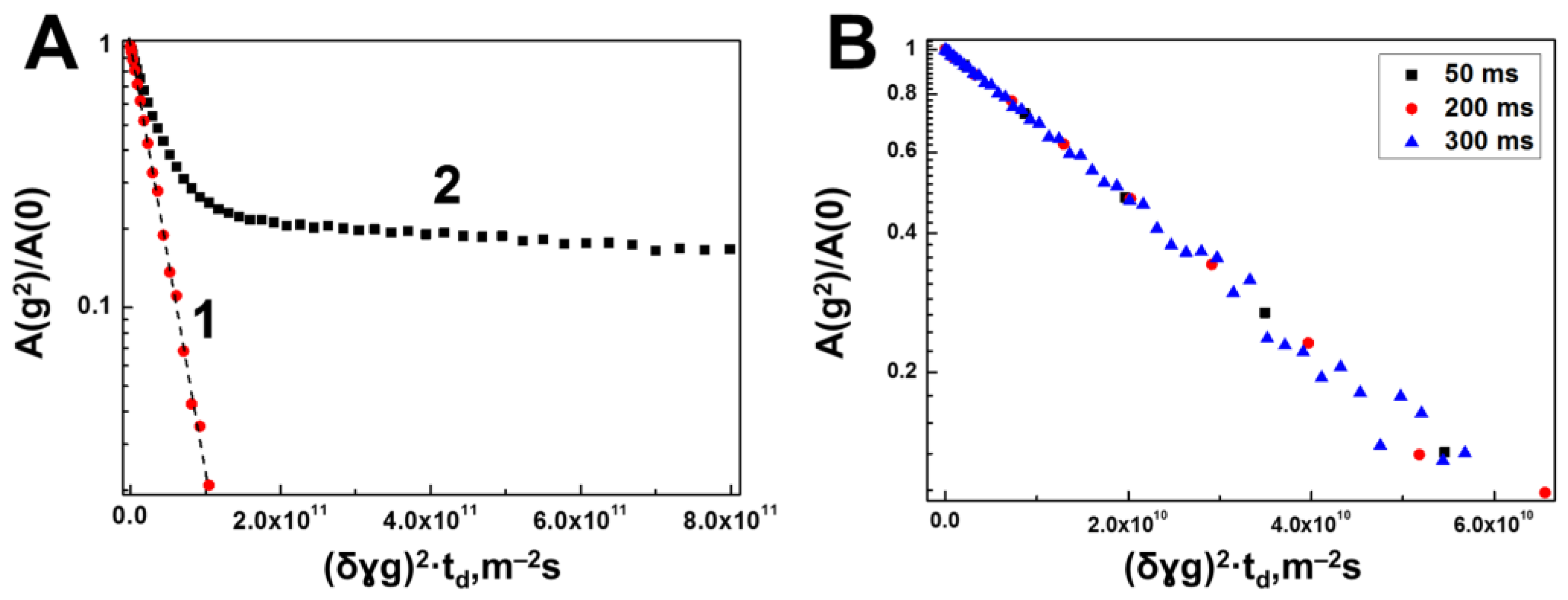
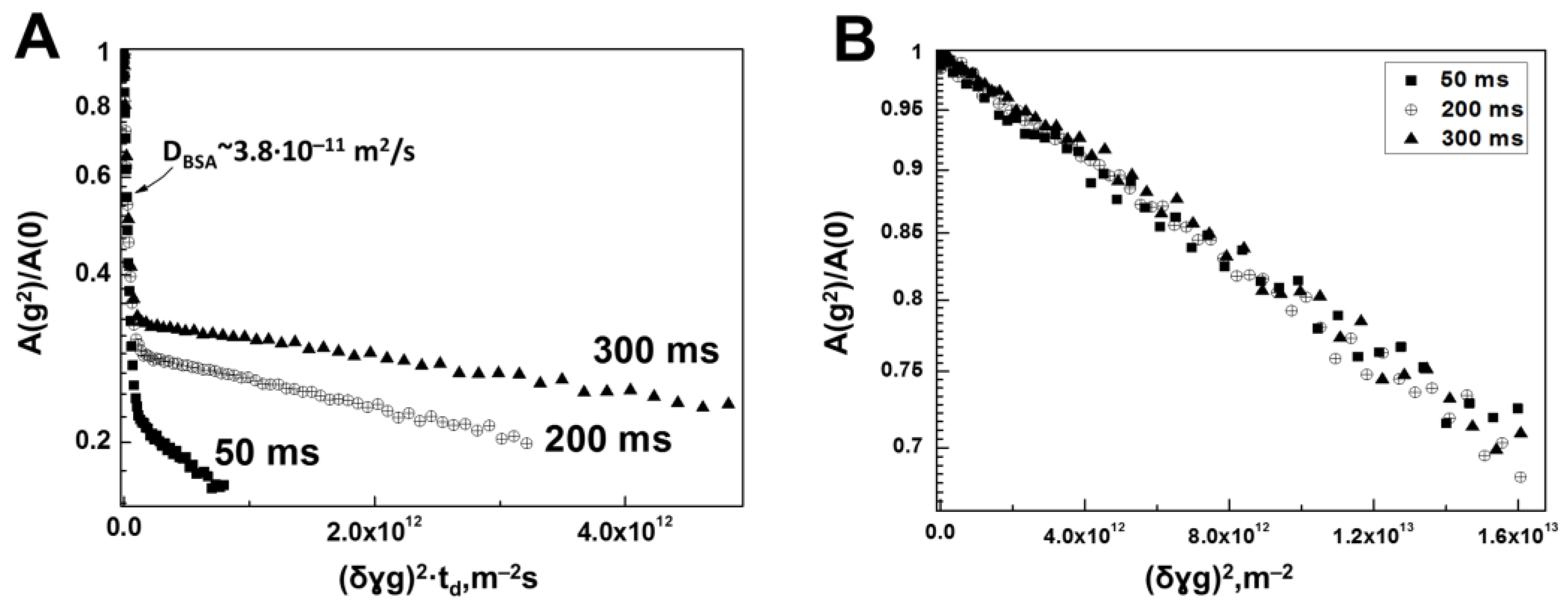
3.1. Interaction of Bovine Serum Albumin with Hyaluronic Acid Molecules
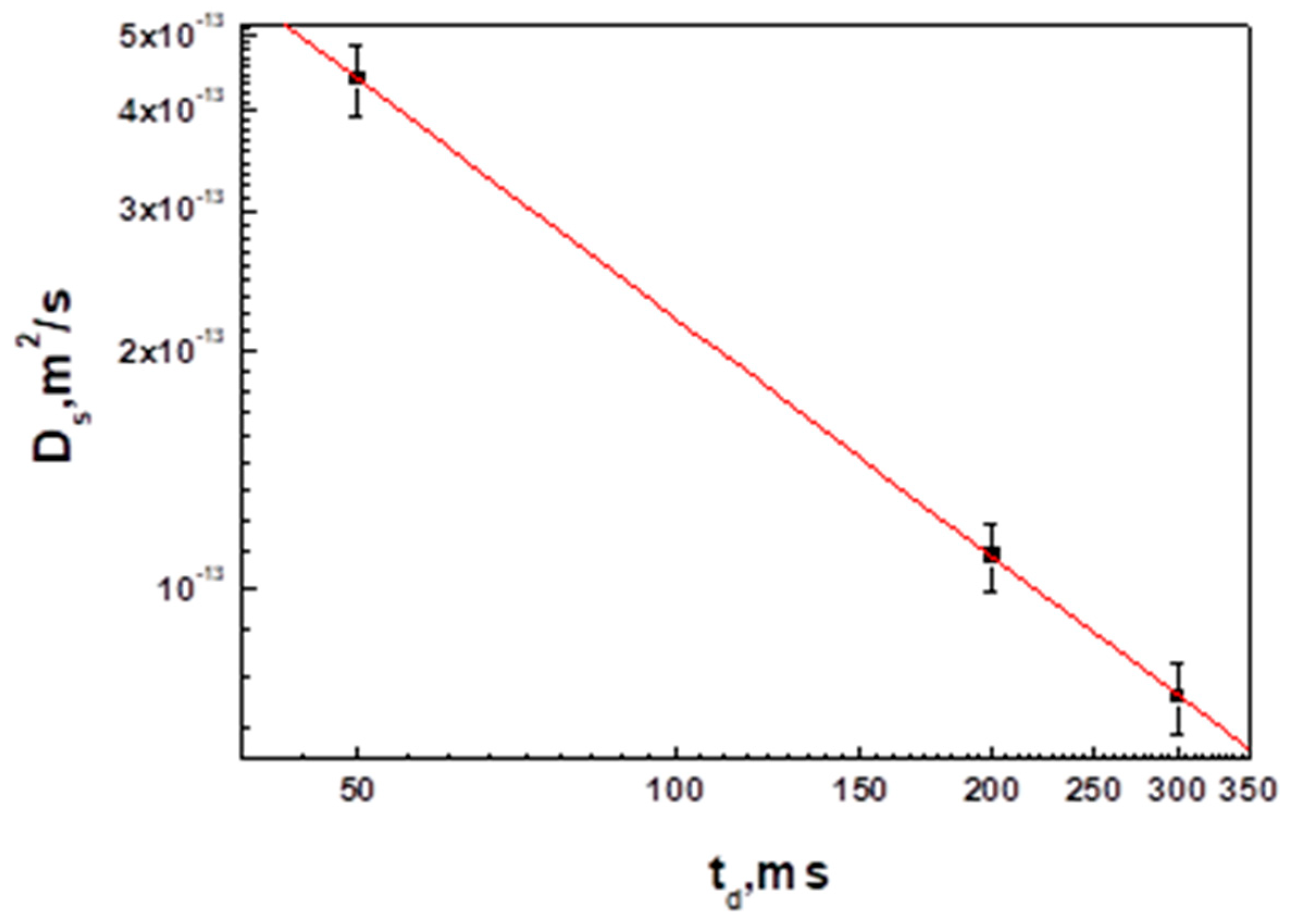
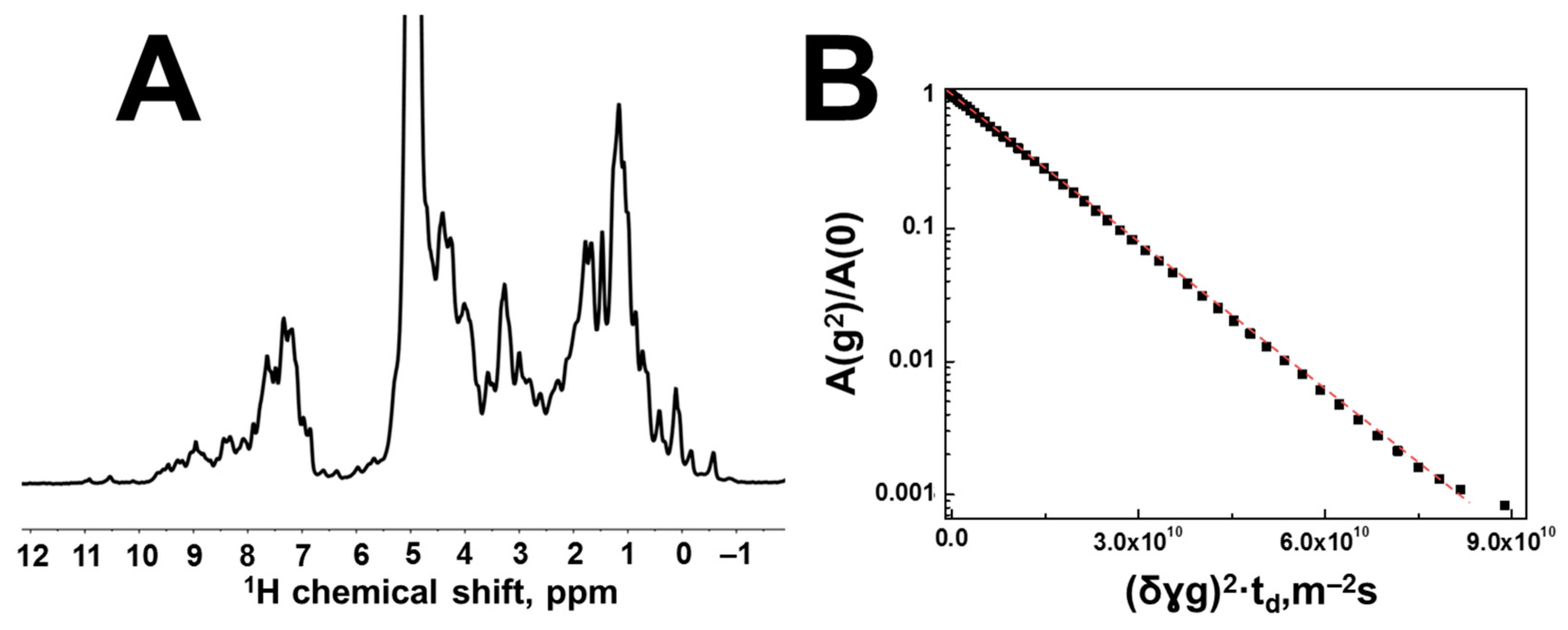
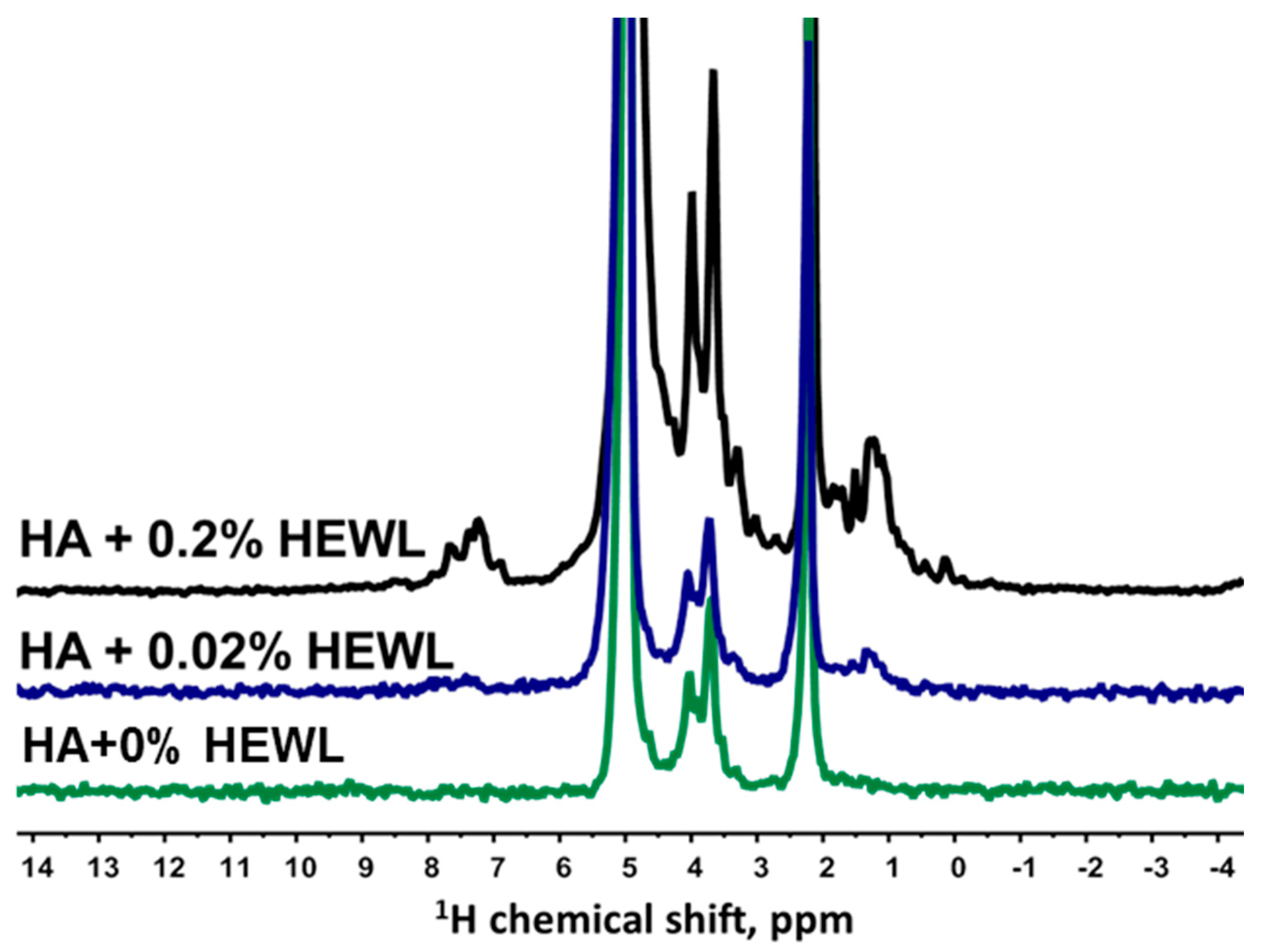
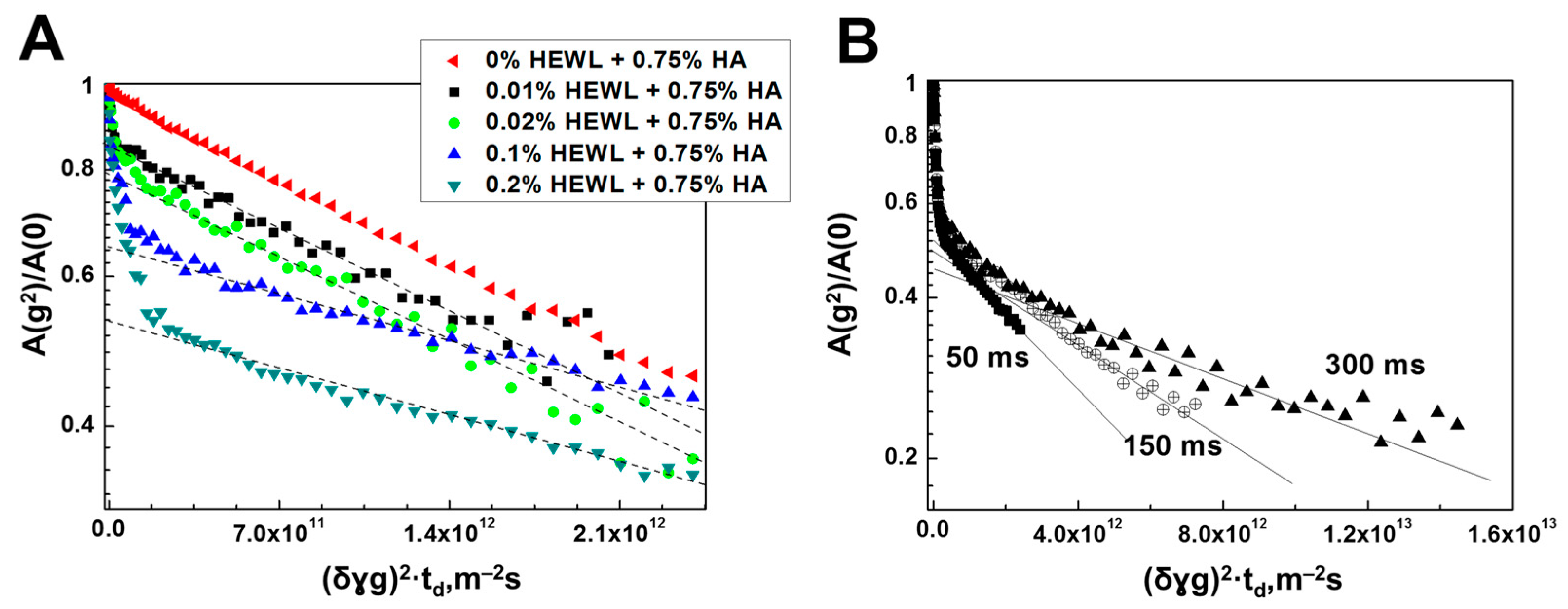
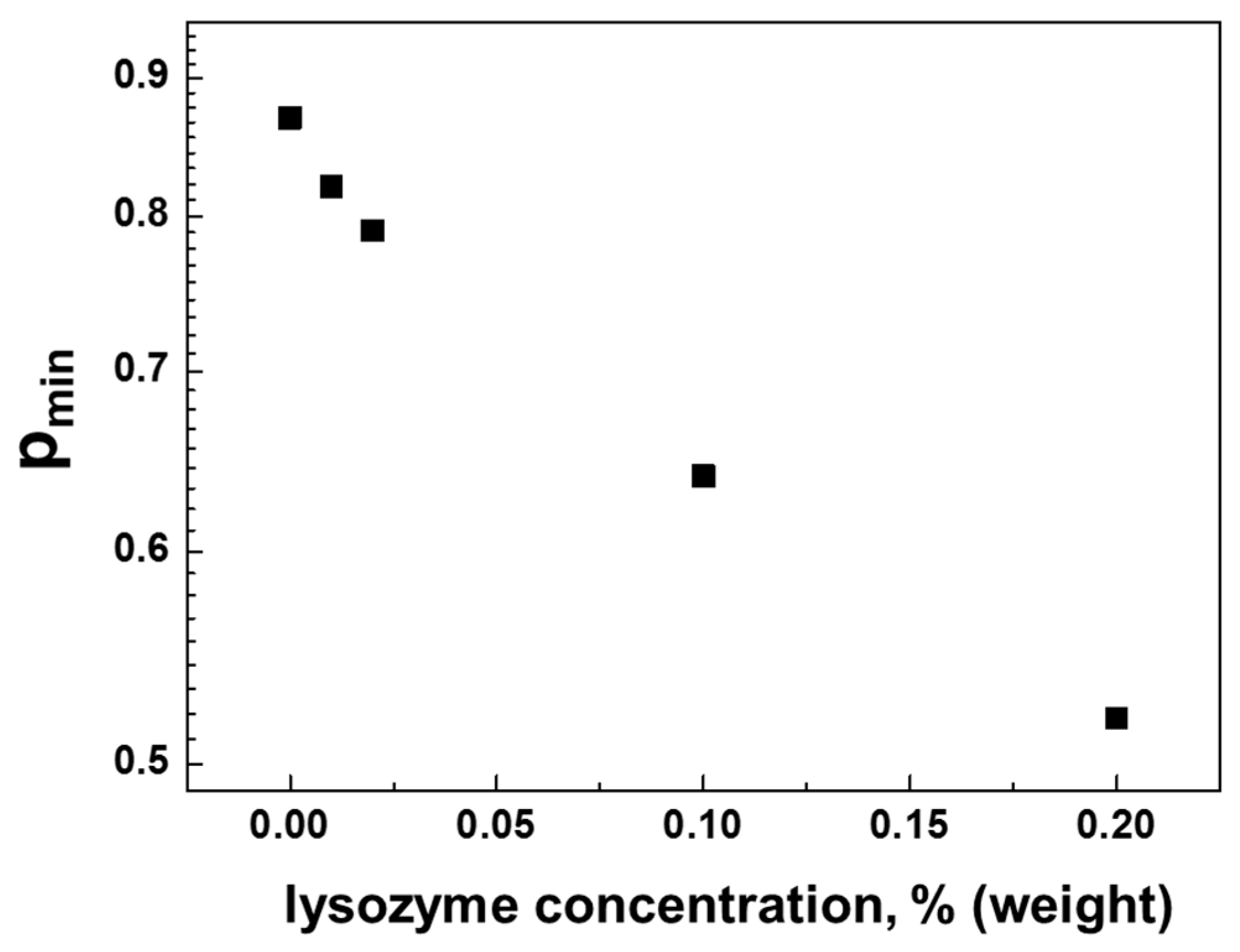
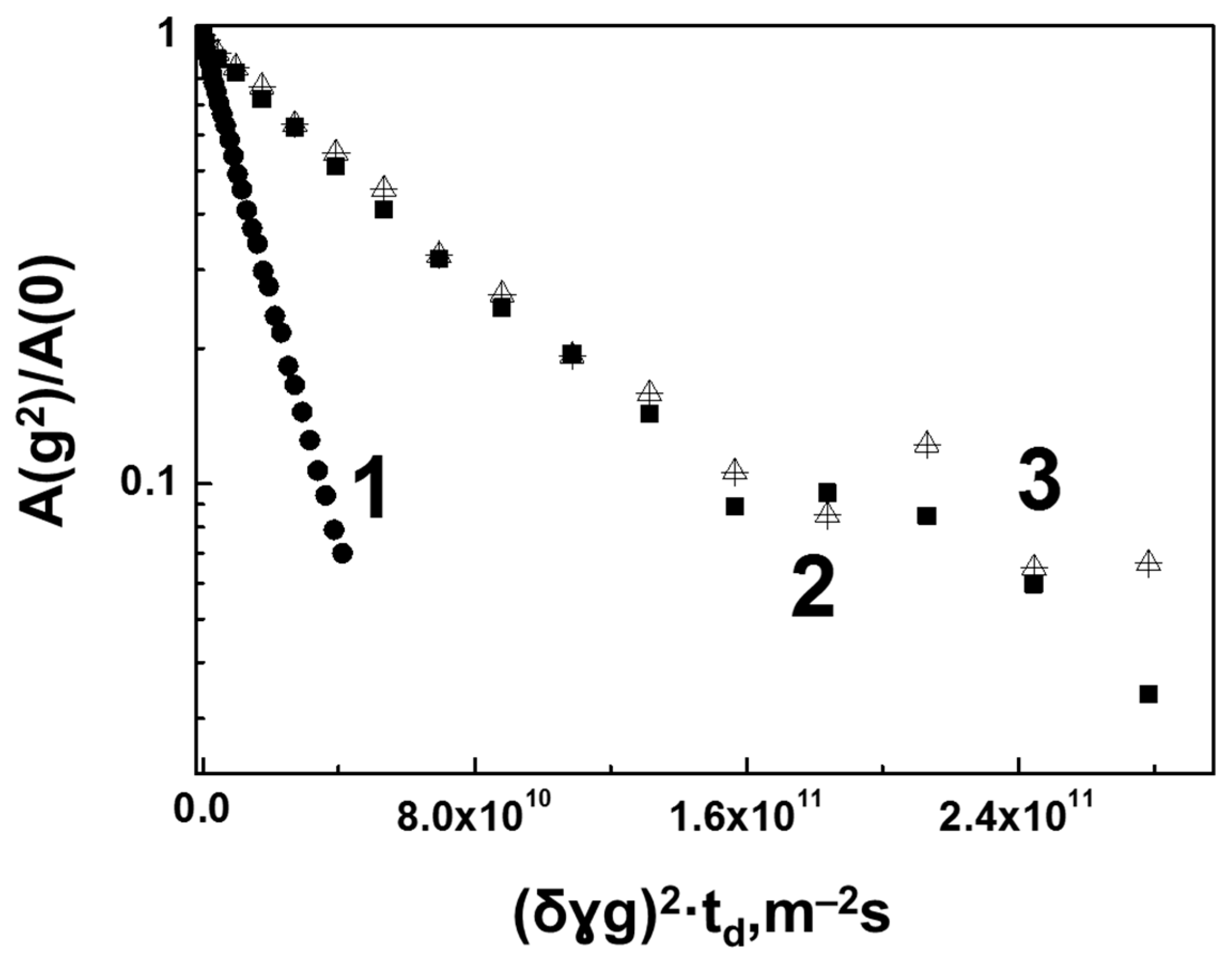
3.2. Interaction of Hen Egg-White Lysozyme (HEWL) with Hyaluronic Acid Molecules
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheetz, M.P. Cellular plasma membrane domains. Mol. Membr. Biol. 1995, 12, 89–91. [Google Scholar] [CrossRef]
- Schmidtke, D.W.; Diamond, S.L. Direct observation of membrane tethers formed during neutrophil attachment to platelets or P-selectin under physiological flow. J. Cell Biol. 2000, 149, 719–730. [Google Scholar] [CrossRef]
- Roux, A.; Cappello, G.; Cartaud, J.; Prost, J.; Goud, B.; Bassereau, P. A minimal system allowing tubulation with molecular motors pulling on giant liposomes. Proc. Natl. Acad. Sci. USA 2002, 99, 5394–5399. [Google Scholar] [CrossRef]
- Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H.H. Nanotubular highways for intercellular organelle transport. Science 2004, 303, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Koster, G.; VanDuijn, M.; Hofs, B.; Dogterom, M. Membrane tube formation from giant vesicles by dynamic association of motor proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 15583–15588. [Google Scholar] [CrossRef]
- Afratis, N.; Gialeli, C.; Nikitovic, D.; Tsegenidis, T.; Karousou, E.; Theocharis, A.D.; Pavao, M.S.; Tzanakakis, G.N.; Karamanos, N.K. Glycosaminoglycans: Key players in cancer cell biology and treatment. FEBS J. 2012, 279, 1177–1197. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan as an immune regulator in human diseases. Physiol. Rev. 2011, 91, 221–264. [Google Scholar] [CrossRef]
- Vigetti, D.; Rizzi, M.; Moretto, P.; Deleonibus, S.; Dreyfuss, J.M.; Karousou, E.; Viola, M.; Clerici, M.; Hascall, V.C.; Ramoni, M.F.; et al. Glycosaminoglycans and glucose prevent apoptosis in 4-methylumbelliferone-treated human aortic smooth muscle cells. J. Biol. Chem. 2011, 286, 34497–34503. [Google Scholar] [CrossRef] [PubMed]
- Dicker, K.T.; Gurski, L.A.; Pradhan-Bhatt, S.; Witt, R.L.; Farach-Carson, M.C.; Jia, X. Hyaluronan: A simple polysaccharide with diverse biological functions. Acta Biomater. 2014, 10, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Melnikova, D.L.; Skirda, V.D.; Nesmelova, I.V. Effect of Intrinsic Disorder and Self-Association on the Translational Diffusion of Proteins: The Case of α-Casein. J. Phys. Chem. B 2017, 121, 2980–2988. [Google Scholar] [CrossRef]
- Nesmelova, I.V.; Skirda, V.D.; Fedotov, V.D. Generalized concentration dependence of globular protein self-diffusion coefficients in aqueous solutions. Biopolymers 2002, 63, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Sathyadevi, P.; Krishnamoorthy, P.; Butorac, R.R.; Cowley, A.H.; Bhuvanesh, N.S.; Dharmaraj, N. Effect of substitution and planarity of the ligand on DNA/BSA interaction, free radical scavenging and cytotoxicity of diamagnetic Ni(II) complexes: A systematic investigation. Dalton Trans. 2011, 40, 9690–9702. [Google Scholar] [CrossRef]
- Fasano, M.; Curry, S.; Terreno, E.; Galliano, M.; Fanali, G.; Narciso, P.; Notari, S.; Ascenzi, P. The extraordinary ligand binding properties of human serum albumin. IUBMB Life 2005, 57, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Galiniak, S.; Bartosz, G. Kinetics of glycoxidation of bovine serum albumin by methylglyoxal and glyoxal and its prevention by various compounds. Molecules 2014, 19, 4880–4896. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Voronina, P.A.; Goncharov, N.V. Integrative Role of Albumin: Evolutionary, Biochemical and Pathophysiological Aspects. J. Evol. Biochem. Physiol. 2021, 57, 1419–1448. [Google Scholar] [CrossRef]
- Xu, X.; Jha, A.K.; Harrington, D.A.; Farach-Carson, M.C.; Jia, X. Hyaluronic Acid-Based Hydrogels: From a Natural Polysaccharide to Complex Networks. Soft Matter 2012, 8, 3280–3294. [Google Scholar] [CrossRef]
- Ribitsch, G.; Schurz, J.; Ribitsch, V. Investigation of the solution structure of hyaluronic acid by light scattering, SAXS, and viscosity measurements. Colloid Polym. Sci. 1980, 258, 1322–1334. [Google Scholar] [CrossRef]
- Heatley, F.; Scott, J.E. A water molecule participates in the secondary structure of hyaluronan. Biochem. J. 1988, 254, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.E.; Cummings, C.; Brass, A.; Chen, Y. Secondary and tertiary structures of hyaluronan in aqueous solution, investigated by rotary shadowing-electron microscopy and computer simulation. Hyaluronan is a very efficient network-forming polymer. Biochem. J. 1991, 274, 699–705. [Google Scholar] [CrossRef]
- Scott, J.E.; Heatley, F. Hyaluronan forms specific stable tertiary structures in aqueous solution: A 13C NMR study. Proc. Natl. Acad. Sci. USA 1999, 96, 4850–4855. [Google Scholar] [CrossRef]
- Fallacara, A.; Baldini, E.; Manfredini, S.; Vertuani, S. Hyaluronic Acid in the Third Millennium. Polymers 2018, 10, 701. [Google Scholar] [CrossRef]
- Venkataramani, S.; Truntzer, J.; Coleman, D.R. Thermal stability of high concentration lysozyme across varying pH: A Fourier Transform Infrared study. J. Pharm. Bioallied Sci. 2013, 5, 148. [Google Scholar] [CrossRef]
- Johnson, L.N.; Cheetham, J.; McLaughlin, P.J.; Acharya, K.R.; Barford, D.; Phillips, D.C. Protein-oligosaccharide interactions: Lysozyme, phosphorylase, amylases. In Carbohydrate-Protein Interaction; Part of the Current Topics in Microbiology and Immunology Book Series (CT MICROBIOLOGY); Springer: Berlin/Heidelberg, Germany, 1988; Volume 139, pp. 81–134. [Google Scholar] [CrossRef]
- Stone, B.A.; Svensson, B.; Collins, M.E.; Rastall, R.A. Polysaccharide degradation. In Glycoscience; Springer: Berlin/Heidelberg, Germany, 2008; pp. 2325–2375. ISBN 978-3-540-36154-1. [Google Scholar] [CrossRef]
- Ardizzoni, A.; Neglia, R.G.; Baschieri, M.C.; Cermelli, C.; Caratozzolo, M.; Righi, E.; Palmieri, B.; Blasi, E. Influence of hyaluronic acid on bacterial and fungal species, including clinically relevant opportunistic pathogens. J. Mater. Sci. Mater. Med. 2011, 22, 2329–2338. [Google Scholar] [CrossRef]
- Day, A.J.; Prestwich, G.D. Hyaluronan-binding proteins: Tying up the giant. J. Biol. Chem. 2002, 277, 4585–4588. [Google Scholar] [CrossRef]
- Toole, B.P. Hyaluronan: From extracellular glue to pericellular cue. Nat. Rev. Cancer 2004, 4, 528–539. [Google Scholar] [CrossRef]
- Murai, T. Lipid raft-mediated regulation of hyaluronan–CD44 interactions in inflammation and cancer. Front. Immunol. 2015, 6, 420. [Google Scholar] [CrossRef]
- Naor, D.; Sionov, R.V.; Ish-Shalom, D. CD44: Structure, function and association with the malignant process. Adv. Cancer Res. 1997, 71, 241–319. [Google Scholar] [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Tanner, J.E. Use of the stimulated echo in NMR diffusion studies. J. Chem. Phys. 1970, 52, 2523–2526. [Google Scholar] [CrossRef]
- Maklakov, A.I.; Skirda, V.D.; Fatkullin, N.F. Self-Diffusion in Polymer Solutions and Melts; Kazan University Press: Kazan, Russia, 1987; p. 224. [Google Scholar]
- Sadler, P.J.; Tucker, A. Proton NMR studies of bovine serum albumin. Assignment of spin systems. Eur. J. Biochem. 1992, 205, 631–643. [Google Scholar] [CrossRef]
- Belinskaia, D.A.; Voronina, P.A.; Shmurak, V.I.; Vovk, M.A.; Batalova, A.A.; Jenkins, R.O.; Goncharov, N.V. The Universal Soldier: Enzymatic and Non-Enzymatic Antioxidant Functions of Serum Albumin. Antioxidants 2020, 9, 966. [Google Scholar] [CrossRef] [PubMed]
- Youm, I.; Agrahari, V.; Murowchick, J.B.; Youan, B.B. Uptake and cytotoxicity of docetaxel-loaded hyaluronic acid-grafted oily core nanocapsules in MDA-MB 231 cancer cells. Pharm. Res. 2014, 31, 2439–2452. [Google Scholar] [CrossRef] [PubMed]
- Gafurov, I.R.; Skirda, V.D.; Maklakov, A.I.; Perevezentseva, S.P.; Zimkin, Y.A. NMR Study of the Structure of Aqueous Gelatine Gels and the Process of Their Formation. Polym. Sci. 1989, 31, 292–300. [Google Scholar] [CrossRef]
- Melnikova, D.L.; Skirda, V.D.; Nesmelova, I.V. Effect of Reducing Agent TCEP on Translational Diffusion and Supramolecular Assembly in Aqueous Solutions of α-Casein. J. Phys. Chem. B 2019, 123, 2305–2315. [Google Scholar] [CrossRef]
- Filippov, A.; Artamonova, M.; Rudakova, M.; Gimatdinov, R.; Skirda, V. Self-diffusion in a hyaluronic acid-albumin-water system as studied by NMR. Magn. Reason. Chem. 2012, 50, 114–119. [Google Scholar] [CrossRef]
- Masschalck, B.; Michiels, C.W. Antimicrobial properties of lysozyme in relation to foodborne vegetative bacteria. Crit. Rev. Microbiol. 2003, 29, 191–214. [Google Scholar] [CrossRef]
- Minones Conde, M.; Conde, O.; Trillo, J.M.; Minones, J., Jr. Interactions in monolayers: A study of the behavior of poly(methyl methacrylate)-lysozyme mixed films from surface pressure-area and ellipsometric measurements. J. Phys. Chem. B 2011, 115, 8667–8678. [Google Scholar] [CrossRef]
- Falcone, P.; Speranza, B.; Del Nobile, M.A.; Corbo, M.R.; Sinigaglia, M. A study on the antimicrobial activity of thymol intended as a natural preservative. J. Food Prot. 2005, 68, 1664–1670. [Google Scholar] [CrossRef]
- Du, X.; Dubin, P.L.; Hoagland, D.A.; Sun, L. Protein-selective coacervation with hyaluronic acid. Biomacromolecules 2014, 15, 726–734. [Google Scholar] [CrossRef]
- Kapelner, R.A.; Yeong, V.; Obermeyer, A.C. Molecular determinants of protein-based coacervates. Curr. Opin. Colloid Interface Sci. 2021, 52, 101407. [Google Scholar] [CrossRef]
- Chang, L.W.; Lytle, T.K.; Radhakrishna, M.; Madinya, J.J.; Vélez, J.; Sing, C.E.; Perry, S.L. Sequence and entropy-based control of complex coacervates. Nat. Commun. 2017, 8, 1273. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Eckmann, C.R.; Courson, D.S.; Rybarska, A.; Hoege, C.; Gharakhani, J.; Jülicher, F.; Hyman, A.A. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science 2009, 324, 1729–1732. [Google Scholar] [CrossRef] [PubMed]
- Lunde, B.M.; Moore, C.; Varani, G. RNA-binding proteins: Modular design for efficient function. Nat. Rev. Mol. Cell Biol. 2007, 8, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Kedersha, N. RNA granules. J. Cell Biol. 2006, 172, 803–808. [Google Scholar] [CrossRef]
- Malay, O.; Bayraktar, O.; Batigün, A. Complex coacervation of silk fibroin and hyaluronic acid. Int. J. Biol. Macromol. 2007, 40, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Izutsu, K.I.; Usui, A.; Yamamoto, E.; Abe, Y.; Yoshida, H.; Goda, Y. Effect of Complex Coacervation with Hyaluronic Acid on Protein Transition in a Subcutaneous Injection Site Model System. Chem. Pharm. Bull. 2020, 68, 1109–1112. [Google Scholar] [CrossRef]
- Dickerson, R.E.; Geis, I. Structure and Action of Proteins; Benjamin-Cummings Publishing Company: San Francisco, CA, USA, 1969. [Google Scholar]
- Proctor, V.A.; Cunningham, F.E.; Fung, D.Y.C. The chemistry of lysozyme and its use as a food preservative and a pharmaceutical. Crit. Rev. Food Sci. Nutr. 1988, 26, 359–395. [Google Scholar] [CrossRef]
- McCarter, J.D.; Withers, G.S. Mechanisms of enzymatic glycoside hydrolysis. Curr. Opin. Struct. Biol. 1994, 4, 885–892. [Google Scholar] [CrossRef]
- Scaman, C.; Nakai, S.; Aminlari, M. Effect of pH, temperature and sodium bisulfite or cysteine on the level of Maillard-based conjugation of lysozyme with dextran, galactomannan and mannan. Food Chem. 2006, 99, 368–380. [Google Scholar] [CrossRef]
- Masschalck, B.; Van Houdt, R.; Van Haver, E.G.; Michiels, C.W. Inactivation of gram-negative bacteria by lysozyme, denatured lysozyme, and lysozyme-derived peptides under high hydrostatic pressure. Appl. Environ. Microbiol. 2001, 67, 339–344. [Google Scholar] [CrossRef]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of lysozyme, an innate immune defense factor, as an alternative antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnikova, D.; Khisravashirova, C.; Smotrina, T.; Skirda, V. Interaction of Hyaluronan Acid with Some Proteins in Aqueous Solution as Studied by NMR. Membranes 2023, 13, 436. https://doi.org/10.3390/membranes13040436
Melnikova D, Khisravashirova C, Smotrina T, Skirda V. Interaction of Hyaluronan Acid with Some Proteins in Aqueous Solution as Studied by NMR. Membranes. 2023; 13(4):436. https://doi.org/10.3390/membranes13040436
Chicago/Turabian StyleMelnikova, Daria, Catherine Khisravashirova, Tatiana Smotrina, and Vladimir Skirda. 2023. "Interaction of Hyaluronan Acid with Some Proteins in Aqueous Solution as Studied by NMR" Membranes 13, no. 4: 436. https://doi.org/10.3390/membranes13040436
APA StyleMelnikova, D., Khisravashirova, C., Smotrina, T., & Skirda, V. (2023). Interaction of Hyaluronan Acid with Some Proteins in Aqueous Solution as Studied by NMR. Membranes, 13(4), 436. https://doi.org/10.3390/membranes13040436