
Membrane Models and Experiments Suitable for Studies of the Cholesterol Bilayer Domains



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Liposomes with High Chol Content
2.1. Liposome Preparation
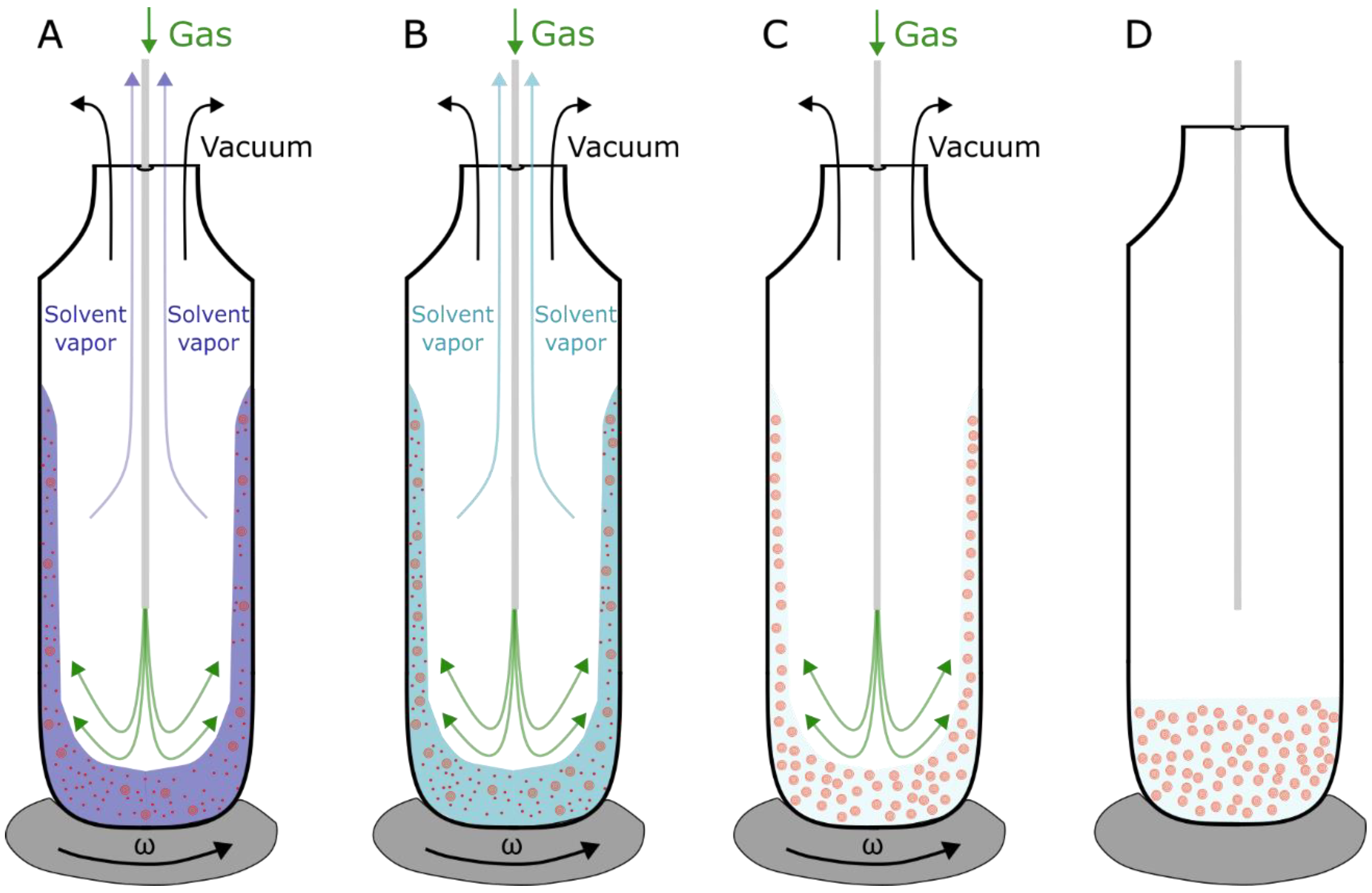
2.1.1. MLVs, SUVs and LUVs
2.1.2. GUVs
2.2. Experimental Techniques Utilizing Liposome Models with Particular Focus on CBDs
2.2.1. X-ray Diffraction
2.2.2. SR EPR Spin Labeling
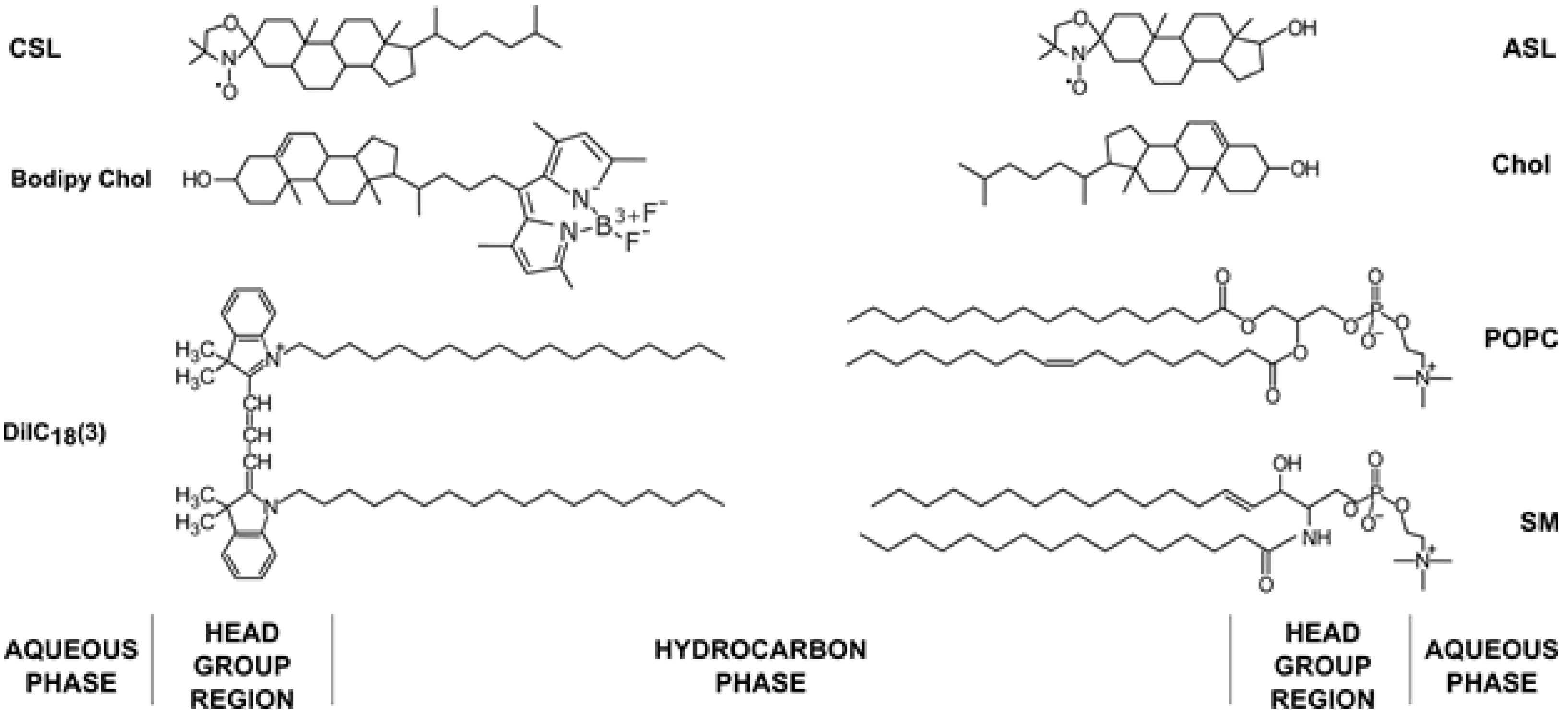
2.2.3. Fluorescence Microscopy
3. SLBs with High Chol Concentrations
3.1. SLBs Preparation
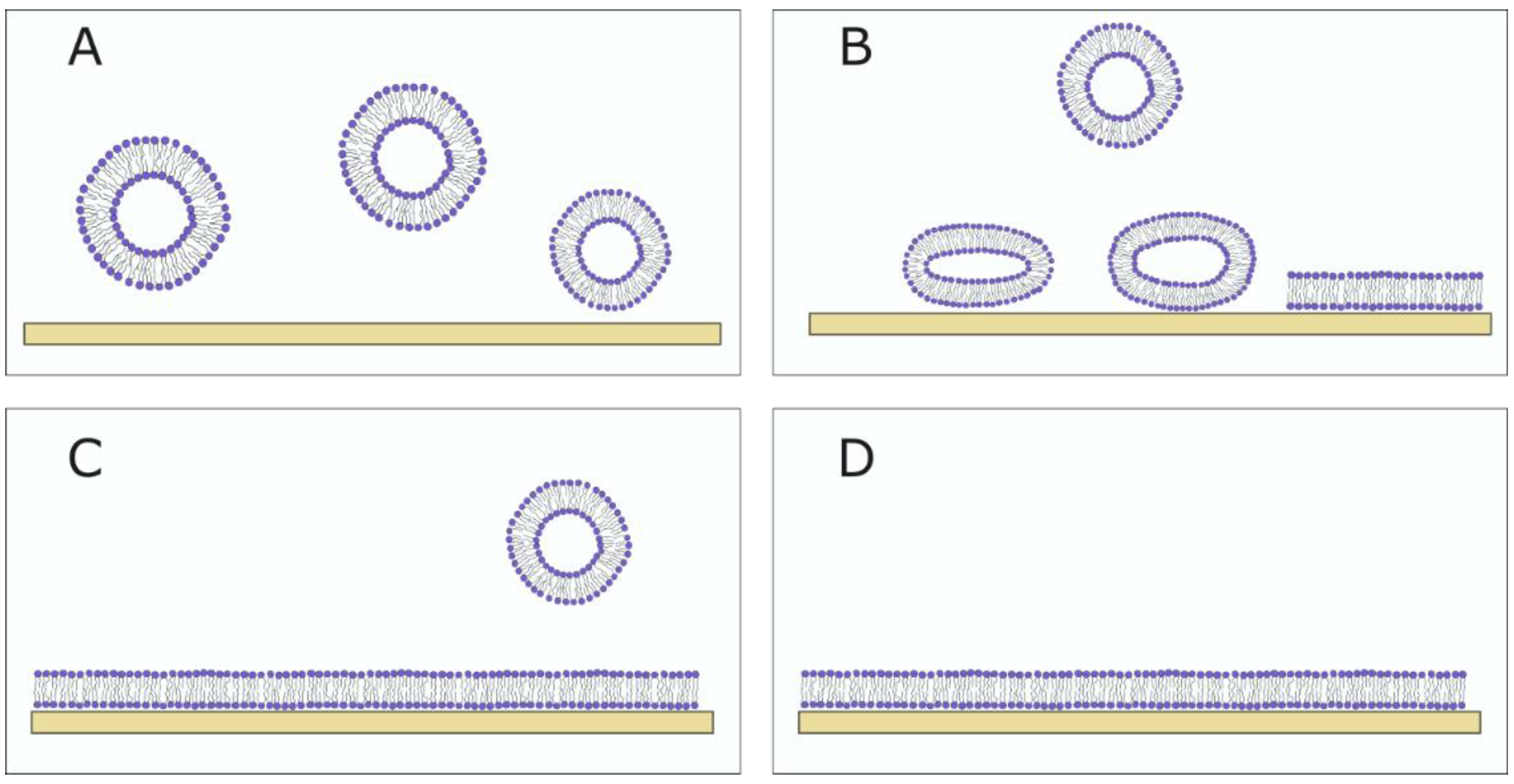
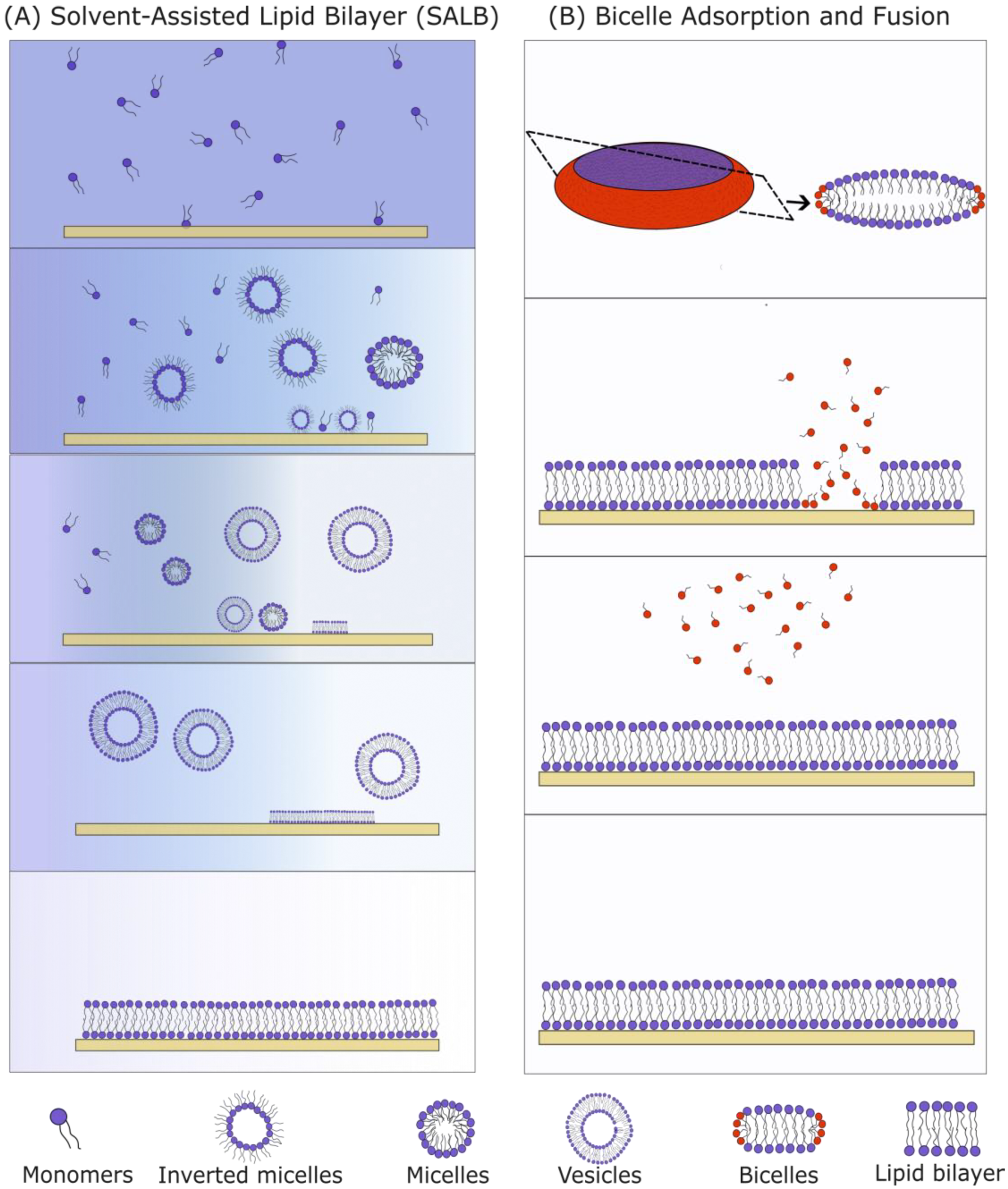
3.1.1. Vesicle Fusion SLBs
3.1.2. Solvent-Assisted and Bicelle-Mediated SLBs
3.2. Experimental Techniques Utilizing SLBs with Particular Focus on CBDs
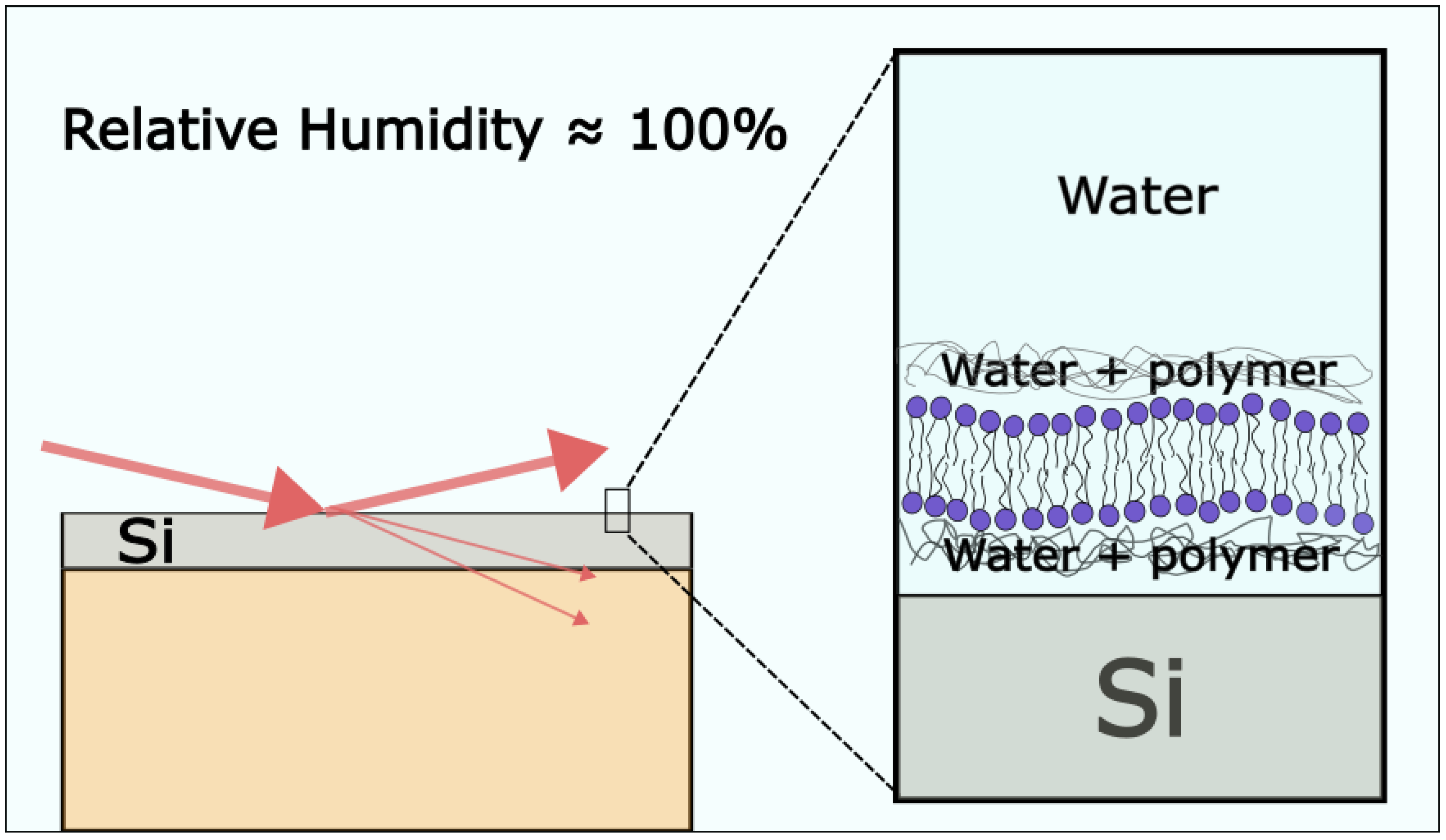
3.2.1. X-ray Diffraction
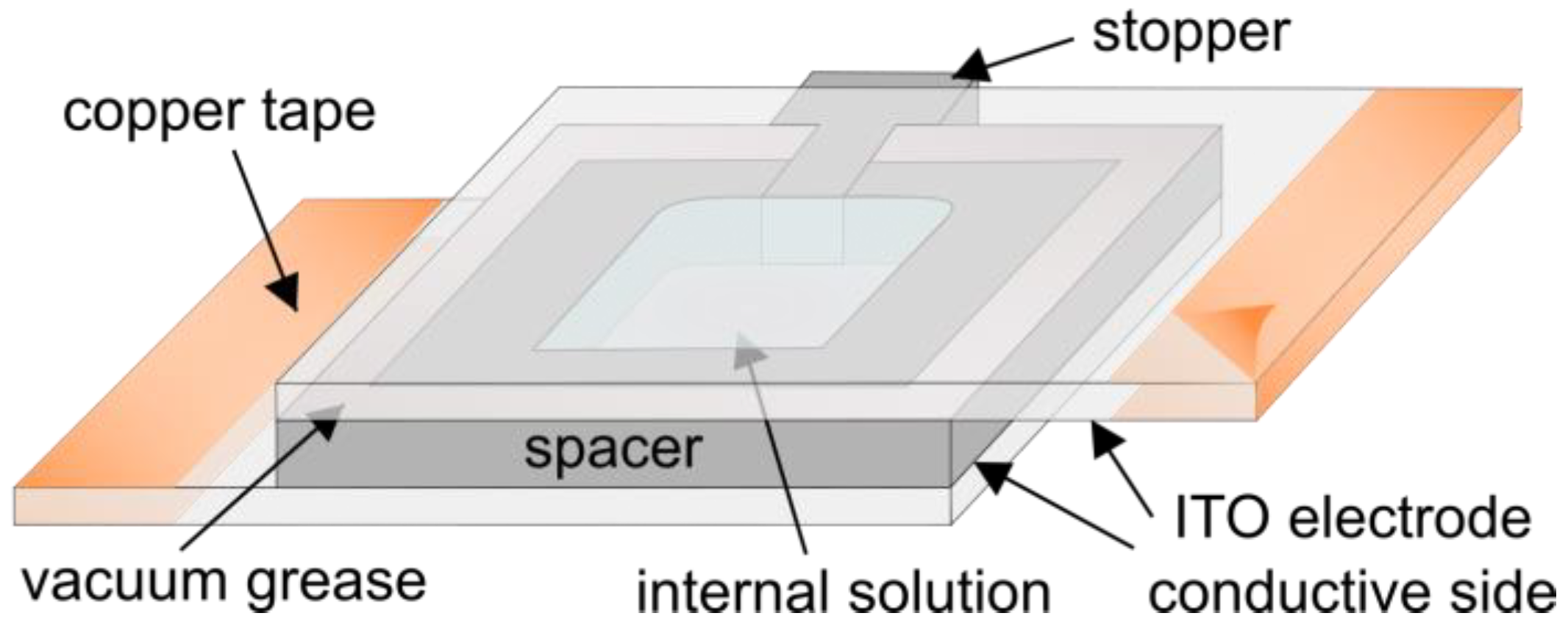
3.2.2. Atomic Force Microscopy
3.2.3. Fluorescence Microscopy
3.2.4. QCM-D
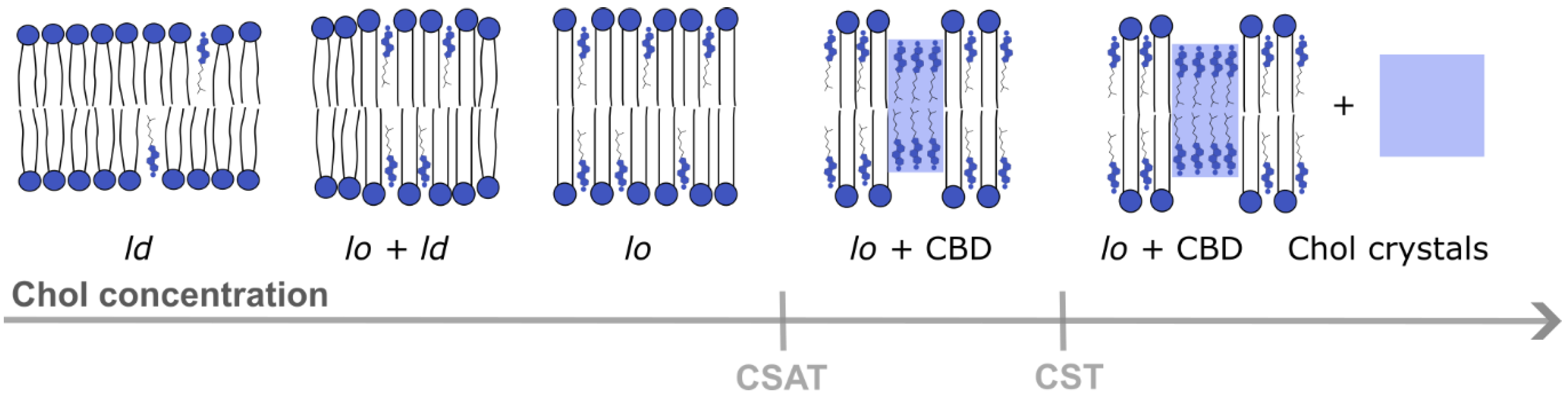
4. Properties of CBDs
5. Understanding of CBD Functions in Model Membranes Helps to Understand Its Functions in Biological Membranes
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AFM | Atomic force microscopy |
| ASL | Androstane spin label |
| CBD | Cholesterol bilayer domain |
| Chol | Cholesterol |
| CSL | Cholestane spin label |
| CST | Cholesterol solubility threshold |
| CSAT | Cholesterol saturation threshold |
| DPPC | 1,2-dipalmitoyl-sn-glycero-3-phosphocholine |
| DiIC18(3) | 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine |
| FRAP | Fluorescence recovery after photobleaching |
| GUV | Giant unilamellar vesicle |
| ld | Liquid-disordered domain |
| lo | Liquid-ordered domain |
| LUV | Large unilamellar vesicle |
| MLV | Multilamellar vesicle |
| MVV | Multivesicular vesicle |
| OLV | Oligolamellar vesicle |
| PL | Phospholipid |
| PM | Plasma membrane |
| POPC | 1-palmitoyl-2-oleoyl-glycero-3-phosphocholine |
| QCM-d | Quartz crystal microbalance with dissipation monitoring |
| RSE | Rapid solvent exchange |
| SALB | Solvent-assisted lipid bilayer |
| SM | Sphingomyelin |
| SLB | Supported lipid bilayer |
| SR EPR | Saturation recovery electron paramagnetic resonance |
| SUV | Small unilamellar vesicle |
References
- Harayama, T.; Riezman, H. Understanding the Diversity of Membrane Lipid Composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, G.L. The Fluid—Mosaic Model of Membrane Structure: Still Relevant to Understanding the Structure, Function and Dynamics of Biological Membranes after More than 40 years. Biochim. Biophys. Acta Biomembr. 2014, 1838, 1451–1466. [Google Scholar] [CrossRef] [PubMed]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane Lipid Composition: Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [PubMed]
- Róg, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. Ordering Effects of Cholesterol and Its Analogues. Biochim. Biophys. Acta Biomembr. 2009, 1788, 97–121. [Google Scholar] [CrossRef] [PubMed]
- Gumí-Audenis, B.; Costa, L.; Carlá, F.; Comin, F.; Sanz, F.; Giannotti, M. Structure and Nanomechanics of Model Membranes by Atomic Force Microscopy and Spectroscopy: Insights into the Role of Cholesterol and Sphingolipids. Membranes 2016, 6, 58. [Google Scholar] [CrossRef]
- Harder, T. Formation of Functional Cell Membrane Domains: The Interplay of Lipid– and Protein–Mediated Interactions. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2003, 358, 863–868. [Google Scholar] [CrossRef]
- Gupta, A.; Phang, I.Y.; Wohland, T. To Hop or Not to Hop: Exceptions in the FCS Diffusion Law. Biophys. J. 2020, 118, 2434–2447. [Google Scholar] [CrossRef]
- Incardona, J.P.; Eaton, S. Cholesterol in Signal Transduction. Curr. Opin. Cell Biol. 2000, 12, 193–203. [Google Scholar] [CrossRef]
- Redondo-Morata, L.; Giannotti, M.I.; Sanz, F. Influence of Cholesterol on the Phase Transition of Lipid Bilayers: A Temperature-Controlled Force Spectroscopy Study. Langmuir 2012, 28, 12851–12860. [Google Scholar] [CrossRef]
- Kotenkov, S.A.; Gnezdilov, O.I.; Khaliullina, A.V.; Antzutkin, O.N.; Gimatdinov, R.S.; Filippov, A.V. Effect of Cholesterol and Curcumin on Ordering of DMPC Bilayers. Appl. Magn. Reson. 2019, 50, 511–520. [Google Scholar] [CrossRef]
- Rabinovich, A.L.; Kornilov, V.V.; Balabaev, N.K.; Leermakers, F.A.M.; Filippov, A.V. Properties of Unsaturated Phospholipid Bilayers: Effect of Cholesterol. Biol. Membr. 2007, 24, 490–505. [Google Scholar] [CrossRef]
- Filippov, A.V.; Rudakova, M.A.; Oradd, G.; Lindblom, G. Lateral Diffusion of Saturated Phosphatidylcholines in Cholesterol-Containing Bilayers. Biophysics 2007, 52, 307–314. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional Rafts in Cell Membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Viola, A. The Amplification of TCR Signaling by Dynamic Membrane Microdomains. Trends Immunol. 2001, 22, 322–327. [Google Scholar] [CrossRef]
- Kusumi, A.; Fujiwara, T.K.; Chadda, R.; Xie, M.; Tsunoyama, T.A.; Kalay, Z.; Kasai, R.S.; Suzuki, K.G.N. Dynamic Organizing Principles of the Plasma Membrane That Regulate Signal Transduction: Commemorating the Fortieth Anniversary of Singer and Nicolson’s Fluid-Mosaic Model. Annu. Rev. Cell Dev. Biol. 2012, 28, 215–250. [Google Scholar] [CrossRef]
- Simons, K.; Toomre, D. Lipid Rafts and Signal Transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, A.; Fujiwara, T.K.; Tsunoyama, T.A.; Kasai, R.S.; Liu, A.; Hirosawa, K.M.; Kinoshita, M.; Matsumori, N.; Komura, N.; Ando, H.; et al. Defining Raft Domains in the Plasma Membrane. Traffic 2020, 21, 106–137. [Google Scholar] [CrossRef]
- Huang, S.; Lim, S.Y.; Gupta, A.; Bag, N.; Wohland, T. Plasma Membrane Organization and Dynamics Is Probe and Cell Line Dependent. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1483–1492. [Google Scholar] [CrossRef]
- Pinkwart, K.; Schneider, F.; Lukoseviciute, M.; Sauka-Spengler, T.; Lyman, E.; Eggeling, C.; Sezgin, E. Nanoscale Dynamics of Cholesterol in the Cell Membrane. J. Biol. Chem. 2019, 294, 12599–12609. [Google Scholar] [CrossRef]
- Raguz, M.; Kumar, S.N.; Zareba, M.; Ilic, N.; Mainali, L.; Subczynski, W.K. Confocal Microscopy Confirmed That in Phosphatidylcholine Giant Unilamellar Vesicles with Very High Cholesterol Content Pure Cholesterol Bilayer Domains Form. Cell Biochem. Biophys. 2019, 77, 309–317. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; Subczynski, W.K. Formation of Cholesterol Bilayer Domains Precedes Formation of Cholesterol Crystals in Cholesterol/Dimyristoylphosphatidylcholine Membranes: EPR and DSC Studies. J. Phys. Chem. B 2013, 117, 8994–9003. [Google Scholar] [CrossRef] [PubMed]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of Membranes Derived from the Total Lipids Extracted from Clear and Cataractous Lenses of 61–70-Year-Old Human Donors. Eur. Biophys. J. 2015, 44, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes: A Review. Cell Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Raguz, M.; Widomska, J.; Mainali, L.; Konovalov, A. Functions of Cholesterol and the Cholesterol Bilayer Domain Specific to the Fiber-Cell Plasma Membrane of the Eye Lens. J. Membr. Biol. 2012, 245, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Widomska, J.; Subczynski, W.K. Why Is Very High Cholesterol Content Beneficial for the Eye Lens but Negative for Other Organs? Nutrients 2019, 11, 1083. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Mainali, L.; Raguz, M.; O’Brien, W.J. Organization of Lipids in Fiber-Cell Plasma Membranes of the Eye Lens. Exp. Eye Res. 2017, 156, 79–86. [Google Scholar] [CrossRef]
- Preston Mason, R.; Tulenko, T.N.; Jacob, R.F. Direct Evidence for Cholesterol Crystalline Domains in Biological Membranes: Role in Human Pathobiology. Biochim. Biophys. Acta Biomembr. 2003, 1610, 198–207. [Google Scholar] [CrossRef]
- Ertel, A.; Marangoni, A.G.; Marsh, J.; Hallett, F.R.; Wood, J.M. Mechanical Properties of Vesicles. I. Coordinated Analysis of Osmotic Swelling and Lysis. Biophys. J. 1993, 64, 426–434. [Google Scholar] [CrossRef]
- Reeves, J.P.; Dowben, R.M. Formation and Properties of Thin-Walled Phospholipid Vesicles. J. Cell. Physiol. 1969, 73, 49–60. [Google Scholar] [CrossRef]
- Buzás, E.I.; Tóth, E.Á.; Sódar, B.W.; Szabó-Taylor, K.É. Molecular Interactions at the Surface of Extracellular Vesicles. Semin. Immunopathol. 2018, 40, 453–464. [Google Scholar] [CrossRef]
- Menger, F.M.; Angelova, M.I. Giant Vesicles: Imitating the Cytological Processes of Cell Membranes. Acc. Chem. Res. 1998, 31, 789–797. [Google Scholar] [CrossRef]
- Valkenier, H.; López Mora, N.; Kros, A.; Davis, A.P. Visualization and Quantification of Transmembrane Ion Transport into Giant Unilamellar Vesicles. Angew. Chemie Int. Ed. 2015, 54, 2137–2141. [Google Scholar] [CrossRef]
- Chacko, I.A.; Ghate, V.M.; Dsouza, L.; Lewis, S.A. Lipid Vesicles: A Versatile Drug Delivery Platform for Dermal and Transdermal Applications. Colloids Surf. B Biointerfaces 2020, 195, 111262. [Google Scholar] [CrossRef] [PubMed]
- Mihailescu, M.; Krepkiy, D.; Milescu, M.; Gawrisch, K.; Swartz, K.J.; White, S. Structural Interactions of a Voltage Sensor Toxin with Lipid Membranes. Proc. Natl. Acad. Sci. USA 2014, 111, 5463–5470. [Google Scholar] [CrossRef] [PubMed]
- Karal, M.A.S.; Ahammed, S.; Levadny, V.; Belaya, M.; Ahamed, M.K.; Ahmed, M.; Bin Mahbub, Z.; Ullah, A.K.M.A. Deformation and Poration of Giant Unilamellar Vesicles Induced by Anionic Nanoparticles. Chem. Phys. Lipids 2020, 230, 104916. [Google Scholar] [CrossRef]
- Luisi, P.L. Toward the Engineering of Minimal Living Cells. Anat. Rec. 2002, 268, 208–214. [Google Scholar] [CrossRef]
- Akbarzadeh, A.; Rezaei-Sadabady, R.; Davaran, S.; Joo, S.W.; Zarghami, N.; Hanifehpour, Y.; Samiei, M.; Kouhi, M.; Nejati-Koshki, K. Liposome: Classification, Preparation, and Applications. Nanoscale Res. Lett. 2013, 8, 102. [Google Scholar] [CrossRef]
- Walde, P. Preparation of Vesicles (Liposomes). Encycl. Nanosci. Nanotechnol. 2004, 8, 43–79. [Google Scholar]
- Buboltz, J.T.; Feigenson, G.W. A Novel Strategy for the Preparation of Liposomes: Rapid Solvent Exchange. Biochim. Biophys. Acta Biomembr. 1999, 1417, 232–245. [Google Scholar] [CrossRef]
- Buboltz, J.T. A More Efficient Device for Preparing Model-Membrane Liposomes by the Rapid Solvent Exchange Method. Rev. Sci. Instrum. 2009, 80, 124301. [Google Scholar] [CrossRef]
- Baykal-Caglar, E.; Hassan-Zadeh, E.; Saremi, B.; Huang, J. Preparation of Giant Unilamellar Vesicles from Damp Lipid Film for Better Lipid Compositional Uniformity. Biochim. Biophys. Acta Biomembr. 2012, 1818, 2598–2604. [Google Scholar] [CrossRef] [PubMed]
- Lind, T.K.; Cárdenas, M. Understanding the Formation of Supported Lipid Bilayers via Vesicle Fusion—A Case That Exemplifies the Need for the Complementary Method Approach (Review). Biointerphases 2016, 11, 020801. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.; Chitneni, M.; Lee, K.; Ming, L.; Yuen, K. Evaluation of Extrusion Technique for Nanosizing Liposomes. Pharmaceutics 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Beales, P.A.; Ciani, B.; Cleasby, A.J. Nature’s Lessons in Design: Nanomachines to Scaffold, Remodel and Shape Membrane Compartments. Phys. Chem. Chem. Phys. 2015, 17, 15489–15507. [Google Scholar] [CrossRef] [PubMed]
- Walde, P.; Cosentino, K.; Engel, H.; Stano, P. Giant Vesicles: Preparations and Applications. ChemBioChem 2010, 11, 848–865. [Google Scholar] [CrossRef]
- van Swaay, D.; DeMello, A. Microfluidic Methods for Forming Liposomes. Lab Chip 2013, 13, 752. [Google Scholar] [CrossRef]
- Patil, Y.P.; Jadhav, S. Novel Methods for Liposome Preparation. Chem. Phys. Lipids 2014, 177, 8–18. [Google Scholar] [CrossRef]
- Rodriguez, N.; Pincet, F.; Cribier, S. Giant Vesicles Formed by Gentle Hydration and Electroformation: A Comparison by Fluorescence Microscopy. Colloids Surf. B Biointerfaces 2005, 42, 125–130. [Google Scholar] [CrossRef]
- Angelova, M.I.; Dimitrov, D.S. Liposome Electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303. [Google Scholar] [CrossRef]
- Dimitrov, D.S.; Angelova, M.I. Lipid Swelling and Liposome Formation on Solid Surfaces in External Electric Fields. In New Trends in Colloid Science; Steinkopff: Darmstadt, Germany, 1987; Volume 56, pp. 48–56. [Google Scholar]
- Boban, Z.; Mardešić, I.; Subczynski, W.K.; Raguz, M. Giant Unilamellar Vesicle Electroformation: What to Use, What to Avoid, and How to Quantify the Results. Membranes 2021, 11, 860. [Google Scholar] [CrossRef]
- Boban, Z.; Puljas, A.; Kovač, D.; Subczynski, W.K.; Raguz, M. Effect of Electrical Parameters and Cholesterol Concentration on Giant Unilamellar Vesicles Electroformation. Cell Biochem. Biophys. 2020, 78, 157–164. [Google Scholar] [CrossRef]
- Boban, Z.; Mardešić, I.; Subczynski, W.K.; Jozić, D.; Raguz, M. Optimization of Giant Unilamellar Vesicle Electroformation for Phosphatidylcholine/Sphingomyelin/Cholesterol Ternary Mixtures. Membranes 2022, 12, 525. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.A.; Zheng, S.; Toppozini, L.A.; Alsop, R.J.; Dies, H.; Wang, A.; Jago, N.; Moore, M.; Rheinstädter, M.C. Solubility of Cholesterol in Lipid Membranes and the Formation of Immiscible Cholesterol Plaques at High Cholesterol Concentrations. Soft Matter 2013, 9, 9342. [Google Scholar] [CrossRef]
- Raguz, M.; Mainali, L.; Widomska, J.; Subczynski, W.K. Using Spin-Label Electron Paramagnetic Resonance (EPR) to Discriminate and Characterize the Cholesterol Bilayer Domain. Chem. Phys. Lipids 2011, 164, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Raguz, M.; Widomska, J.; Dillon, J.; Gaillard, E.R.; Subczynski, W.K. Characterization of Lipid Domains in Reconstituted Porcine Lens Membranes Using EPR Spin-Labeling Approaches. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1079–1090. [Google Scholar] [CrossRef] [PubMed]
- Raguz, M.; Mainali, L.; Widomska, J.; Subczynski, W.K. The Immiscible Cholesterol Bilayer Domain Exists as an Integral Part of Phospholipid Bilayer Membranes. Biochim. Biophys. Acta Biomembr. 2011, 1808, 1072–1080. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Properties of Membranes Derived from the Total Lipids Extracted from the Human Lens Cortex and Nucleus. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1432–1440. [Google Scholar] [CrossRef]
- Mainali, L.; O’Brien, W.J.; Subczynski, W.K. Detection of Cholesterol Bilayer Domains in Intact Biological Membranes: Methodology Development and Its Application to Studies of Eye Lens Fiber Cell Plasma Membranes. Exp. Eye Res. 2019, 178, 72–81. [Google Scholar] [CrossRef]
- Mainali, L.; Raguz, M.; O’Brien, W.J.; Subczynski, W.K. Changes in the Properties and Organization of Human Lens Lipid Membranes Occurring with Age. Curr. Eye Res. 2017, 42, 721–731. [Google Scholar] [CrossRef]
- Richter, R.P.; Bérat, R.; Brisson, A.R. Formation of Solid-Supported Lipid Bilayers: An Integrated View. Langmuir 2006, 22, 3497–3505. [Google Scholar] [CrossRef]
- Brian, A.A.; McConnell, H.M. Allogeneic Stimulation of Cytotoxic T Cells by Supported Planar Membranes. Proc. Natl. Acad. Sci. USA 1984, 81, 6159–6163. [Google Scholar] [CrossRef] [PubMed]
- Lind, T.K.; Cárdenas, M.; Wacklin, H.P. Formation of Supported Lipid Bilayers by Vesicle Fusion: Effect of Deposition Temperature. Langmuir 2014, 30, 7259–7263. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Cho, N.-J. Supported Lipid Bilayer Formation: Beyond Vesicle Fusion. Langmuir 2020, 36, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Tero, R. Substrate Effects on the Formation Process, Structure and Physicochemical Properties of Supported Lipid Bilayers. Materials 2012, 5, 2658–2680. [Google Scholar] [CrossRef]
- Koenig, B.W.; Krueger, S.; Orts, W.J.; Majkrzak, C.F.; Berk, N.F.; Silverton, J.V.; Gawrisch, K. Neutron Reflectivity and Atomic Force Microscopy Studies of a Lipid Bilayer in Water Adsorbed to the Surface of a Silicon Single Crystal. Langmuir 1996, 12, 1343–1350. [Google Scholar] [CrossRef]
- Khadka, N.K.; Timsina, R.; Rowe, E.; O’Dell, M.; Mainali, L. Mechanical Properties of the High Cholesterol-Containing Membrane: An AFM Study. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183625. [Google Scholar] [CrossRef]
- Waldie, S.; Moulin, M.; Porcar, L.; Pichler, H.; Strohmeier, G.A.; Skoda, M.; Forsyth, V.T.; Haertlein, M.; Maric, S.; Cárdenas, M. The Production of Matchout-Deuterated Cholesterol and the Study of Bilayer-Cholesterol Interactions. Sci. Rep. 2019, 9, 5118. [Google Scholar] [CrossRef]
- Waldie, S.; Lind, T.K.; Browning, K.; Moulin, M.; Haertlein, M.; Forsyth, V.T.; Luchini, A.; Strohmeier, G.A.; Pichler, H.; Maric, S.; et al. Localization of Cholesterol within Supported Lipid Bilayers Made of a Natural Extract of Tailor-Deuterated Phosphatidylcholine. Langmuir 2018, 34, 472–479. [Google Scholar] [CrossRef]
- Tabaei, S.R.; Choi, J.; Haw Zan, G.; Zhdanov, V.P.; Cho, N. Solvent-Assisted Lipid Bilayer Formation on Silicon Dioxide and Gold. Langmuir 2014, 30, 10363–10373. [Google Scholar] [CrossRef]
- Tabaei, S.R.; Jackman, J.A.; Kim, S.-O.; Liedberg, B.; Knoll, W.; Parikh, A.N.; Cho, N. Formation of Cholesterol-Rich Supported Membranes Using Solvent-Assisted Lipid Self-Assembly. Langmuir 2014, 30, 13345–13352. [Google Scholar] [CrossRef]
- Hohner, A.O.; David, M.P.C.; Rädler, J.O. Controlled Solvent-Exchange Deposition of Phospholipid Membranes onto Solid Surfaces. Biointerphases 2010, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, R.; Last, J.A.; Slade, A.L.; Ista, L.K.; Bisong, P.; O’Brien, M.J.; Brueck, S.R.J.; Sasaki, D.Y.; Lopez, G.P. Using Bicellar Mixtures To Form Supported and Suspended Lipid Bilayers on Silicon Chips. Langmuir 2006, 22, 8163–8168. [Google Scholar] [CrossRef] [PubMed]
- Alassi, A.; Benammar, M.; Brett, D. Quartz Crystal Microbalance Electronic Interfacing Systems: A Review. Sensors 2017, 17, 2799. [Google Scholar] [CrossRef] [PubMed]
- Sut, T.N.; Jackman, J.A.; Yoon, B.K.; Park, S.; Kolahdouzan, K.; Ma, G.J.; Zhdanov, V.P.; Cho, N. Influence of NaCl Concentration on Bicelle-Mediated SLB Formation. Langmuir 2019, 35, 10658–10666. [Google Scholar] [CrossRef]
- Dudás, E.F.; Wacha, A.; Bóta, A.; Bodor, A. Peptide-Bicelle Interaction: Following Variations in Size and Morphology by a Combined NMR-SAXS Approach. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183095. [Google Scholar] [CrossRef]
- Ziblat, R.; Fargion, I.; Leiserowitz, L.; Addadi, L. Spontaneous Formation of Two-Dimensional and Three-Dimensional Cholesterol Crystals in Single Hydrated Lipid Bilayers. Biophys. J. 2012, 103, 255–264. [Google Scholar] [CrossRef]
- Ziblat, R.; Leiserowitz, L.; Addadi, L. Crystalline Lipid Domains: Characterization by X-Ray Diffraction and Their Relation to Biology. Angew. Chemie Int. Ed. 2011, 50, 3620–3629. [Google Scholar] [CrossRef]
- Ziblat, R.; Leiserowitz, L.; Addadi, L. Crystalline Domain Structure and Cholesterol Crystal Nucleation in Single Hydrated DPPC:Cholesterol:POPC Bilayers. J. Am. Chem. Soc. 2010, 132, 9920–9927. [Google Scholar] [CrossRef]
- Pincet, F.; Adrien, V.; Yang, R.; Delacotte, J.; Rothman, J.E.; Urbach, W.; Tareste, D. FRAP to Characterize Molecular Diffusion and Interaction in Various Membrane Environments. PLoS ONE 2016, 11, e0158457. [Google Scholar] [CrossRef]
- Litz, J.P.; Thakkar, N.; Portet, T.; Keller, S.L. Depletion with Cyclodextrin Reveals Two Populations of Cholesterol in Model Lipid Membranes. Biophys. J. 2016, 110, 635–645. [Google Scholar] [CrossRef]
- Sut, T.N.; Yoon, B.K.; Park, S.; Jackman, J.A. Versatile Formation of Supported Lipid Bilayers from Bicellar Mixtures of Phospholipids and Capric Acid. Sci. Rep. 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sut, T.N.; Park, S.; Choe, Y.; Cho, N.-J. Characterizing the Supported Lipid Membrane Formation from Cholesterol-Rich Bicelles. Langmuir 2019, 35, 15063–15070. [Google Scholar] [CrossRef] [PubMed]
- Heberle, F.A.; Feigenson, G.W. Phase Separation in Lipid Membranes. Cold Spring Harb. Perspect. Biol. 2011, 3, a004630. [Google Scholar] [CrossRef]
- Simons, K.; Vaz, W.L.C. Model Systems, Lipid Rafts, and Cell Membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef]
- Huang, J.; Feigenson, G.W. A Microscopic Interaction Model of Maximum Solubility of Cholesterol in Lipid Bilayers. Biophys. J. 1999, 76, 2142–2157. [Google Scholar] [CrossRef]
- Almeida, P.F.F.; Pokorny, A.; Hinderliter, A. Thermodynamics of Membrane Domains. Biochim. Biophys. Acta Biomembr. 2005, 1720, 1–13. [Google Scholar] [CrossRef]
- Plesnar, E.; Subczynski, W.K.; Pasenkiewicz-Gierula, M. Saturation with Cholesterol Increases Vertical Order and Smoothes the Surface of the Phosphatidylcholine Bilayer: A Molecular Simulation Study. Biochim. Biophys. Acta Biomembr. 2012, 1818, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Brzustowicz, M.R.; Cherezov, V.; Caffrey, M.; Stillwell, W.; Wassall, S.R. Molecular Organization of Cholesterol in Polyunsaturated Membranes: Microdomain Formation. Biophys. J. 2002, 82, 285–298. [Google Scholar] [CrossRef]
- Buzhynskyy, N.; Sens, P.; Behar-Cohen, F.; Scheuring, S. Eye Lens Membrane Junctional Microdomains: A Comparison between Healthy and Pathological Cases. New J. Phys. 2011, 13, 085016. [Google Scholar] [CrossRef]
- Tulenko, T.N.; Chen, M.; Mason, P.E.; Mason, R.P. Physical Effects of Cholesterol on Arterial Smooth Muscle Membranes: Evidence of Immiscible Cholesterol Domains and Alterations in Bilayer Width during Atherogenesis. J. Lipid Res. 1998, 39, 947–956. [Google Scholar] [CrossRef]
- Jacob, R.F.; Cenedella, R.J.; Mason, R.P. Direct Evidence for Immiscible Cholesterol Domains in Human Ocular Lens Fiber Cell Plasma Membranes. J. Biol. Chem. 1999, 274, 31613–31618. [Google Scholar] [CrossRef] [PubMed]
- Mainali, L.; Pasenkiewicz-Gierula, M.; Subczynski, W.K. Formation of Cholesterol Bilayer Domains Precedes Formation of Cholesterol Crystals in Membranes Made of the Major Phospholipids of Human Eye Lens Fiber Cell Plasma Membranes. Curr. Eye Res. 2020, 45, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M. Hypothetical Pathway for Formation of Cholesterol Microcrystals Initiating the Atherosclerotic Process. Cell Biochem. Biophys. 2020, 78, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Kar, R.; Batra, N.; Riquelme, M.A.; Jiang, J.X. Biological Role of Connexin Intercellular Channels and Hemichannels. Arch. Biochem. Biophys. 2012, 524, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Timsina, R.; Mainali, L. Association of Alpha-Crystallin with Fiber Cell Plasma Membrane of the Eye Lens Accompanied by Light Scattering and Cataract Formation. Membranes 2021, 11, 447. [Google Scholar] [CrossRef]
- Timsina, R.; Wellisch, S.; Haemmerle, D.; Mainali, L. Binding of Alpha-Crystallin to Cortical and Nuclear Lens Lipid Membranes Derived from a Single Lens. Int. J. Mol. Sci. 2022, 23, 11295. [Google Scholar] [CrossRef]
- Bassnett, S. Lens Organelle Degradation. Exp. Eye Res. 2002, 74, 1–6. [Google Scholar] [CrossRef]
- Wride, M.A. Lens Fibre Cell Differentiation and Organelle Loss: Many Paths Lead to Clarity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1219–1233. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mardešić, I.; Boban, Z.; Subczynski, W.K.; Raguz, M. Membrane Models and Experiments Suitable for Studies of the Cholesterol Bilayer Domains. Membranes 2023, 13, 320. https://doi.org/10.3390/membranes13030320
Mardešić I, Boban Z, Subczynski WK, Raguz M. Membrane Models and Experiments Suitable for Studies of the Cholesterol Bilayer Domains. Membranes. 2023; 13(3):320. https://doi.org/10.3390/membranes13030320
Chicago/Turabian StyleMardešić, Ivan, Zvonimir Boban, Witold Karol Subczynski, and Marija Raguz. 2023. "Membrane Models and Experiments Suitable for Studies of the Cholesterol Bilayer Domains" Membranes 13, no. 3: 320. https://doi.org/10.3390/membranes13030320
APA StyleMardešić, I., Boban, Z., Subczynski, W. K., & Raguz, M. (2023). Membrane Models and Experiments Suitable for Studies of the Cholesterol Bilayer Domains. Membranes, 13(3), 320. https://doi.org/10.3390/membranes13030320