
Inhibiting Biofilm Formation via Simultaneous Application of Nitric Oxide and Quorum Quenching Bacteria



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. NO Donors for Biofilm Dispersal
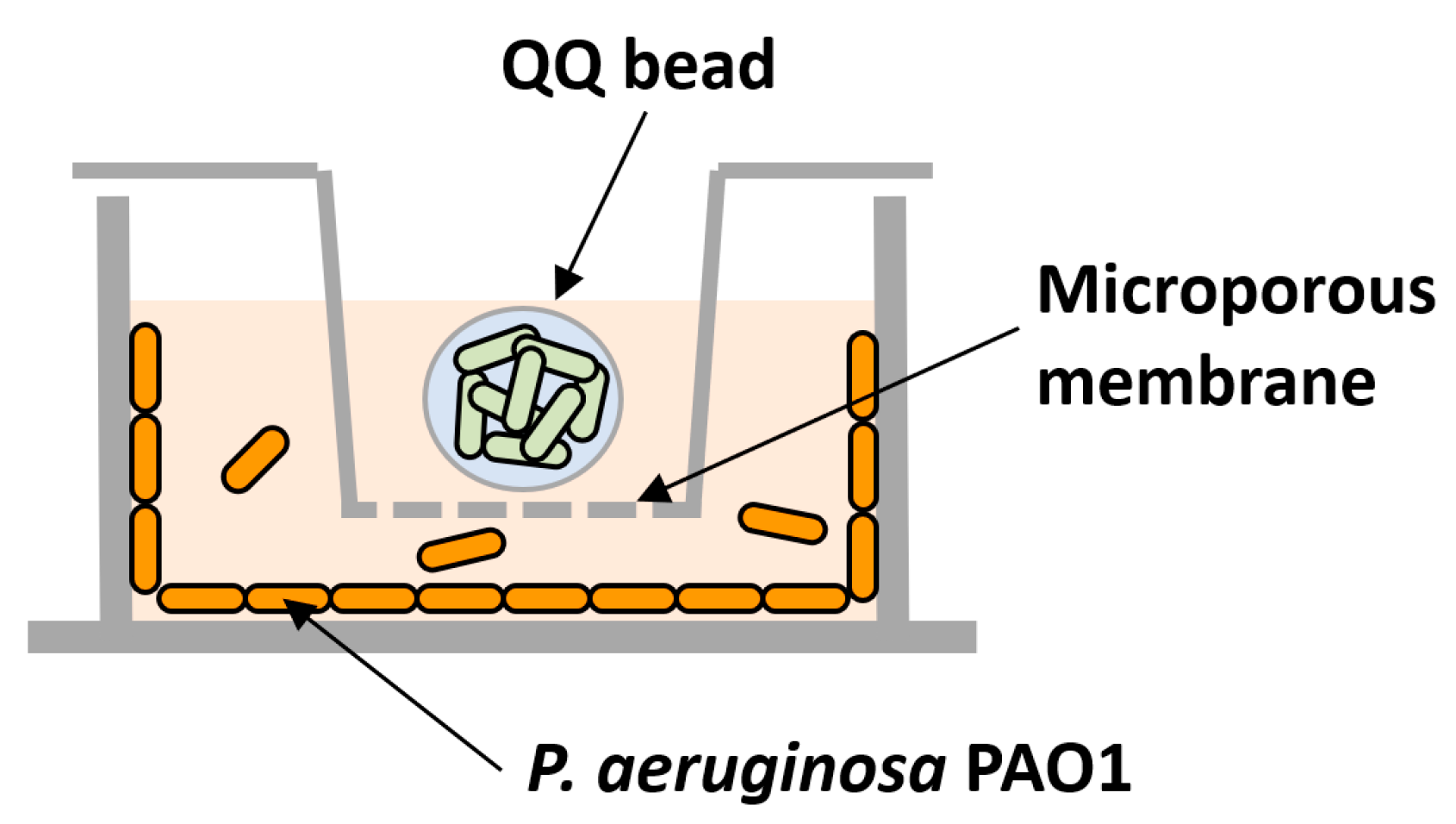
2.3. Preparation of QQ Beads
2.4. Biofilm Formation Assay
2.5. Bioassay for the Quantification of AHLs
2.6. QQ Activity Test
3. Results and Discussion
3.1. Effect of QQ Beads on Biofilm Formation
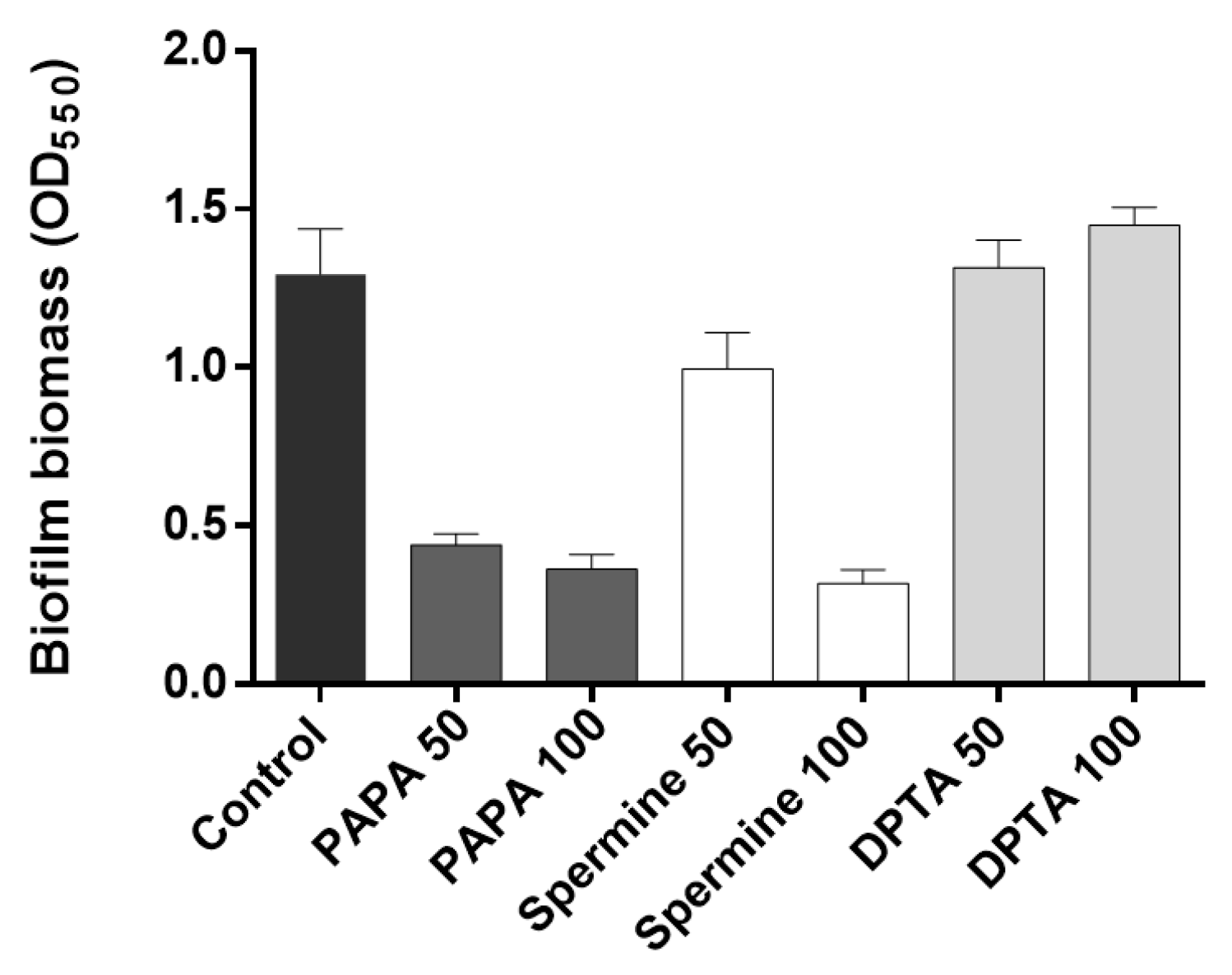
3.2. Dispersal of Biofilm by Addition of NO Donors
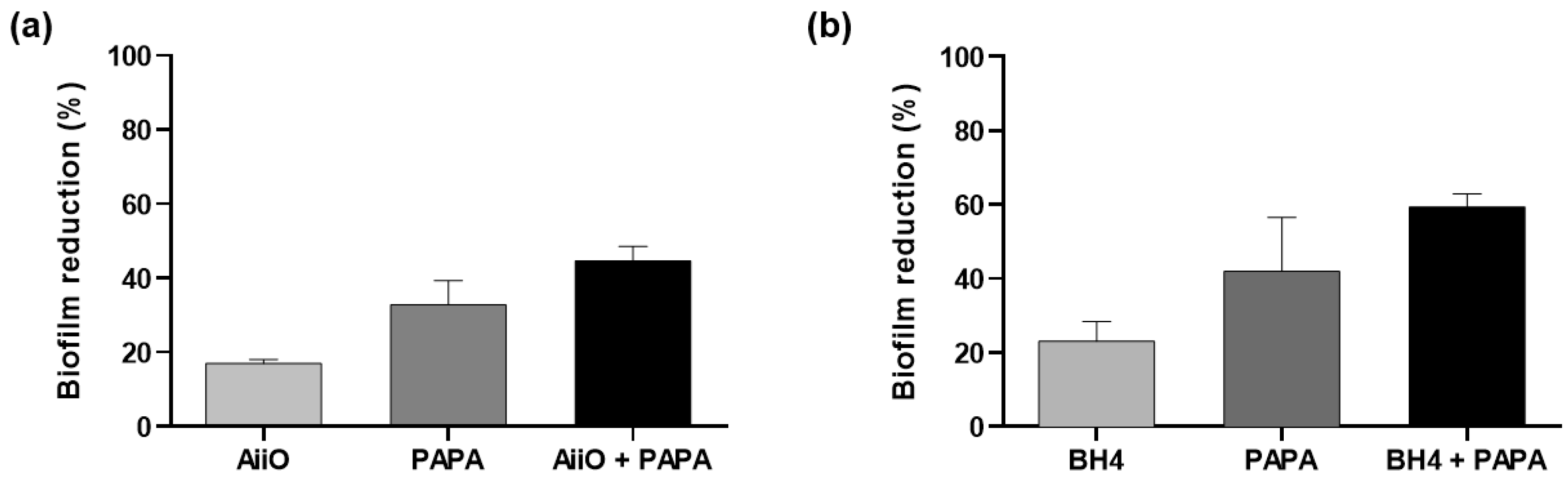
3.3. Combination of NO Treatment and QQ Alginate Beads for Biofilm Reduction
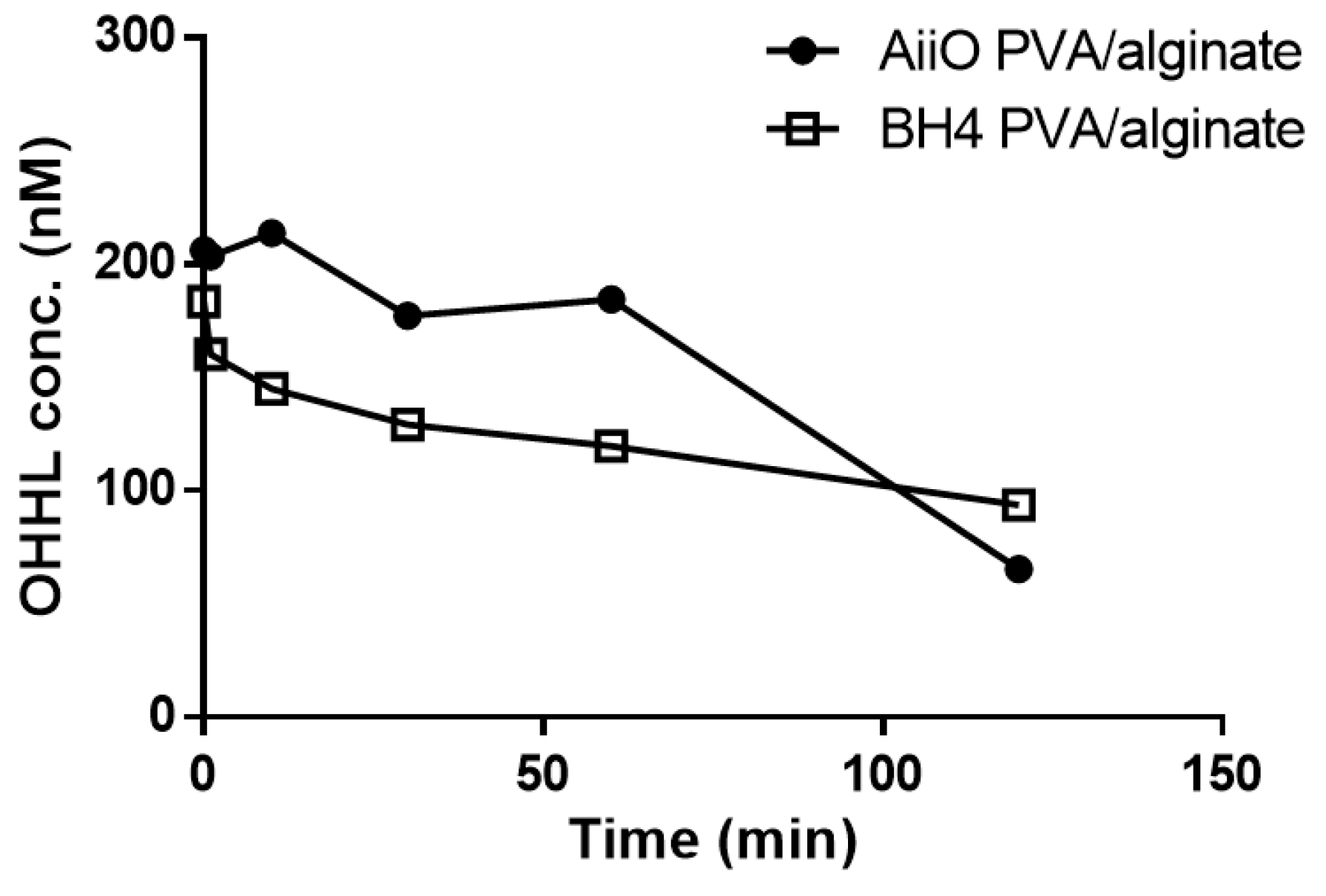
3.4. Combination of NO Treatment and QQ PVA/Alginate Beads
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Drews, A. Membrane Fouling in Membrane Bioreactors-Characterisation, Contradictions, Cause and Cures. J. Membr. Sci. 2010, 363, 1–28. [Google Scholar] [CrossRef]
- Smith, A.L.; Stadler, L.B.; Love, N.G.; Skerlos, S.J.; Raskin, L. Perspectives on Anaerobic Membrane Bioreactor Treatment of Domestic Wastewater: A Critical Review. Bioresour. Technol. 2012, 122, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.K.; Kashif, A.; Rout, P.R.; Aslam, M.; Fuwad, A.; Choi, Y.; Banu, J.R.; Park, J.H.; Kumar, G. A Brief Review of Anaerobic Membrane Bioreactors Emphasizing Recent Advancements, Fouling Issues and Future Perspectives. J. Environ. Manag. 2020, 270, 110909. [Google Scholar] [CrossRef]
- Lee, W.N.; Chang, I.S.; Hwang, B.K.; Park, P.K.; Lee, C.H.; Huang, X. Changes in Biofilm Architecture with Addition of Membrane Fouling Reducer in a Membrane Bioreactor. Process Biochem. 2007, 42, 655–661. [Google Scholar] [CrossRef]
- Jegatheesan, V.; Pramanik, B.K.; Chen, J.; Navaratna, D.; Chang, C.Y.; Shu, L. Treatment of Textile Wastewater with Membrane Bioreactor: A Critical Review. Bioresour. Technol. 2016, 204, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Krzeminski, P.; Leverette, L.; Malamis, S.; Katsou, E. Membrane Bioreactors—A Review on Recent Developments in Energy Reduction, Fouling Control, Novel Configurations, LCA and Market Prospects. J. Membr. Sci. 2017, 527, 207–227. [Google Scholar] [CrossRef]
- Le-Clech, P.; Chen, V.; Fane, T.A.G. Fouling in Membrane Bioreactors Used in Wastewater Treatment. J. Membr. Sci. 2006, 284, 17–53. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y. Biological Control of Microbial Attachment: A Promising Alternative for Mitigating Membrane Biofouling. Appl. Microbiol. Biotechnol. 2010, 86, 825–837. [Google Scholar] [CrossRef]
- Oh, H.S.; Constancias, F.; Ramasamy, C.; Tang, P.Y.P.; Yee, M.O.; Fane, A.G.; McDougald, D.; Rice, S.A. Biofouling Control in Reverse Osmosis by Nitric Oxide Treatment and Its Impact on the Bacterial Community. J. Membr. Sci. 2018, 550, 313–321. [Google Scholar] [CrossRef]
- McDougald, D.; Rice, S.A.; Barraud, N.; Steinberg, P.D.; Kjelleberg, S. Should We Stay or Should We Go: Mechanisms and Ecological Consequences for Biofilm Dispersal. Nat. Rev. Microbiol. 2012, 10, 39–50. [Google Scholar] [CrossRef]
- Cai, Y.M.; Webb, J.S. Optimization of Nitric Oxide Donors for Investigating Biofilm Dispersal Response in Pseudomonas aeruginosa Clinical Isolates. Appl. Microbiol. Biotechnol. 2020, 104, 8859–8869. [Google Scholar] [CrossRef]
- Barraud, N.; Storey, M.V.; Moore, Z.P.; Webb, J.S.; Rice, S.A.; Kjelleberg, S. Nitric Oxide-Mediated Dispersal in Single- and Multi-Species Biofilms of Clinically and Industrially Relevant Microorganisms. Microb. Biotechnol. 2009, 2, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Heil, J.; Vereecken, H.; Brüggemann, N. A Review of Chemical Reactions of Nitrification Intermediates and Their Role in Nitrogen Cycling and Nitrogen Trace Gas Formation in Soil. Eur. J. Soil Sci. 2016, 67, 23–39. [Google Scholar] [CrossRef]
- Barraud, N.; Schleheck, D.; Klebensberger, J.; Webb, J.S.; Hassett, D.J.; Rice, S.A.; Kjelleberg, S. Nitric Oxide Signaling in Pseudomonas aeruginosa Biofilms Mediates Phosphodiesterase Activity, Decreased Cyclic Di-GMP Levels, and Enhanced Dispersal. J. Bacteriol. 2009, 191, 7333–7342. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, J.; Barnes, R.J.; Tan, X.; Mcdougald, D.; Fane, A.G.; Zhuang, G.; Kjelleberg, S.; Cohen, Y.; Rice, S.A. The Application of Nitric Oxide to Control Biofouling of Membrane Bioreactors. Microb. Biotechnol. 2015, 8, 549–560. [Google Scholar] [CrossRef]
- Barnes, R.J.; Bandi, R.R.; Wong, W.S.; Barraud, N.; McDougald, D.; Fane, A.; Kjelleberg, S.; Rice, S.A. Optimal Dosing Regimen of Nitric Oxide Donor Compounds for the Reduction of Pseudomonas aeruginosa Biofilm and Isolates from Wastewater Membranes. Biofouling 2013, 29, 203–212. [Google Scholar] [CrossRef]
- Barnes, R.J.; Bandi, R.R.; Chua, F.; Low, J.H.; Aung, T.; Barraud, N.; Fane, A.G.; Kjelleberg, S.; Rice, S.A. The Roles of Pseudomonas aeruginosa Extracellular Polysaccharides in Biofouling of Reverse Osmosis Membranes and Nitric Oxide Induced Dispersal. J. Membr. Sci. 2014, 466, 161–172. [Google Scholar] [CrossRef]
- Wang, Y.; Bian, Z.; Wang, Y. Biofilm Formation and Inhibition Mediated by Bacterial Quorum Sensing. Appl. Microbiol. Biotechnol. 2022, 106, 6365–6381. [Google Scholar] [CrossRef] [PubMed]
- Anburajan, P.; Kim, Y.; Rice, S.A.; Oh, H.S. Bacterial Signaling and Signal Responses as Key Factors in Water and Wastewater Treatment. J. Water Process Eng. 2021, 44, 102434. [Google Scholar] [CrossRef]
- Noori, A.; Kim, H.; Kim, M.H.; Kim, K.; Lee, K.; Oh, H.S. Quorum Quenching Bacteria Isolated from Industrial Wastewater Sludge to Control Membrane Biofouling. Bioresour. Technol. 2022, 352, 127077. [Google Scholar] [CrossRef]
- Lee, K.; Yu, H.; Zhang, X.; Choo, K.H. Quorum Sensing and Quenching in Membrane Bioreactors: Opportunities and Challenges for Biofouling Control. Bioresour. Technol. 2018, 270, 656–668. [Google Scholar] [CrossRef]
- Passos da Silva, D.; Schofield, M.C.; Parsek, M.R.; Tseng, B.S. An Update on the Sociomicrobiology of Quorum Sensing in Gram-Negative Biofilm Development. Pathogens 2017, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Evans, L.R.; Linker, A. Production and Characterization of the Slime Polysaccharide of Pseudomonas aeruginosa. J. Bacteriol. 1973, 116, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Ryu, D.H.; Lee, S.W.; Mikolaityte, V.; Kim, Y.W.; Jeong, H.; Lee, S.J.; Lee, C.H.; Lee, J.K. Identification of a Second Type of Ahllactonase from Rhodococcus Sp. BH4, Belonging to the α/β Hydrolase Superfamily. J. Microbiol. Biotechnol. 2020, 30, 937–945. [Google Scholar] [CrossRef]
- Oh, H.S.; Tan, C.H.; Low, J.H.; Rzechowicz, M.; Siddiqui, M.F.; Winters, H.; Kjelleberg, S.; Fane, A.G.; Rice, S.A. Quorum Quenching Bacteria Can Be Used to Inhibit the Biofouling of Reverse Osmosis Membranes. Water Res. 2017, 112, 29–37. [Google Scholar] [CrossRef]
- Fuqua, C.; Winans, S.C. Conserved Cis-Acting Promoter Elements Are Required for Density-Dependent Transcription of Agrobacterium Tumefaciens Conjugal Transfer Genes. J. Bacteriol. 1996, 178, 435–440. [Google Scholar] [CrossRef]
- Lee, K.; Kim, Y.W.; Lee, S.; Lee, S.H.; Nahm, C.H.; Kwon, H.; Park, P.K.; Choo, K.H.; Koyuncu, I.; Drews, A.; et al. Stopping Autoinducer-2 Chatter by Means of an Indigenous Bacterium (Acinetobacter Sp. DKY-1): A New Antibiofouling Strategy in a Membrane Bioreactor for Wastewater Treatment. Environ. Sci. Technol. 2018, 52, 6237–6245. [Google Scholar] [CrossRef] [PubMed]
- Duarte, D.; Vale, N. Evaluation of Synergism in Drug Combinations and Reference Models for Future Orientations in Oncology. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100110. [Google Scholar] [CrossRef]
- Syafiuddin, A.; Boopathy, R.; Mehmood, M.A. Recent Advances on Bacterial Quorum Quenching as an Effective Strategy to Control Biofouling in Membrane Bioreactors. Bioresour. Technol. Rep. 2021, 15, 100745. [Google Scholar] [CrossRef]
- Diggle, S.P.; Winzer, K.; Chhabra, S.R.; Worrall, K.E.; Cámara, M.; Williams, P. The Pseudomonas aeruginosa Quinolone Signal Molecule Overcomes the Cell Density-Dependency of the Quorum Sensing Hierarchy, Regulates Rhl-Dependent Genes at the Onset of Stationary Phase and Can Be Produced in the Absence of LasR. Mol. Microbiol. 2003, 50, 29–43. [Google Scholar] [CrossRef]
- Tan, C.H.; Oh, H.S.; Sheraton, V.M.; Mancini, E.; Joachim Loo, S.C.; Kjelleberg, S.; Sloot, P.M.A.; Rice, S.A. Convection and the Extracellular Matrix Dictate Inter- And Intra-Biofilm Quorum Sensing Communication in Environmental Systems. Environ. Sci. Technol. 2020, 54, 6730–6740. [Google Scholar] [CrossRef]
- Zain, N.A.M.; Suhaimi, M.S.; Idris, A. Development and Modification of PVA-Alginate as a Suitable Immobilization Matrix. Process Biochem. 2011, 46, 2122–2129. [Google Scholar] [CrossRef]
- Elwakeel, K.Z.; Ahmed, M.M.; Akhdhar, A.; Sulaiman, M.G.M.; Khan, Z.A. Recent Advances in Alginate-Based Adsorbents for Heavy Metal Retention from Water: A Review. Desalin. Water Treat. 2022, 272, 50–74. [Google Scholar] [CrossRef]
- Bai, Y.; Wu, W. The Neutral Protease Immobilization: Physical Characterization of Sodium Alginate-Chitosan Gel Beads. Appl. Biochem. Biotechnol. 2022, 194, 2269–2283. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Purpose and Characteristics | Incubation Temp. (°C) | Antibiotics | Source |
|---|---|---|---|---|
| P. aeruginosa PAO1 | Model biofilm strain. Wild type | 37 | - | [23] |
| Rhodococcus sp. BH4 | AHL quencher strain. Wild type | 30 | - | [24] |
| E. coli TOP10-Empty | Control strain carrying pTrcHis2 plasmid; KanR, AmpR | 37 | Kanamycin (50 μg/mL) | [25] |
| E. coli TOP10-AiiO | AHL quencher strain carrying pTriHis2-AiiO plasmid; KanR, AmpR | 37 | Kanamycin (50 μg/mL) | [25] |
| A. tumefaciens A136 (Ti−)(pCF218)(pCF372) | AHL biosensor, carrying pCF218 plasmid with traR and pCF372 plasmid with PtraI-lacZ; SpR, TetR | 30 | Spectinomycin (50 μg/mL), Tetracyclin (4.5 μg/mL) | [26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Anburajan, P.; Kim, H.; Oh, H.-S. Inhibiting Biofilm Formation via Simultaneous Application of Nitric Oxide and Quorum Quenching Bacteria. Membranes 2023, 13, 836. https://doi.org/10.3390/membranes13100836
Kim Y, Anburajan P, Kim H, Oh H-S. Inhibiting Biofilm Formation via Simultaneous Application of Nitric Oxide and Quorum Quenching Bacteria. Membranes. 2023; 13(10):836. https://doi.org/10.3390/membranes13100836
Chicago/Turabian StyleKim, Youkyoung, Parthiban Anburajan, Hyeok Kim, and Hyun-Suk Oh. 2023. "Inhibiting Biofilm Formation via Simultaneous Application of Nitric Oxide and Quorum Quenching Bacteria" Membranes 13, no. 10: 836. https://doi.org/10.3390/membranes13100836
APA StyleKim, Y., Anburajan, P., Kim, H., & Oh, H.-S. (2023). Inhibiting Biofilm Formation via Simultaneous Application of Nitric Oxide and Quorum Quenching Bacteria. Membranes, 13(10), 836. https://doi.org/10.3390/membranes13100836