
Mammalian Brain Ca2+ Channel Activity Transplanted into Xenopus laevis Oocytes

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Membrane Preparation
2.2. Xenopus Oocytes Isolation and Injection
2.3. Calcium Current Recordings
2.4. Data Analysis
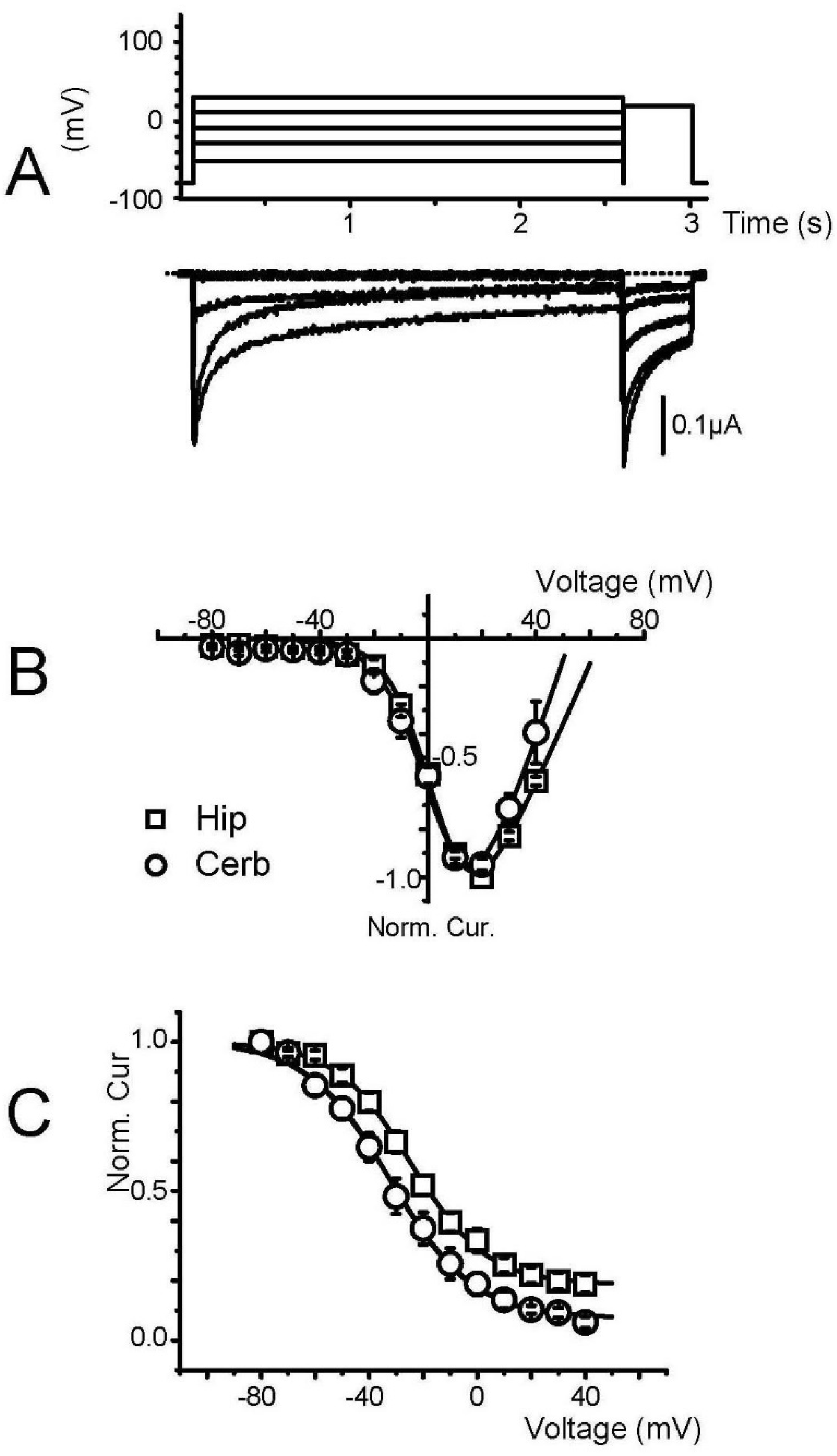
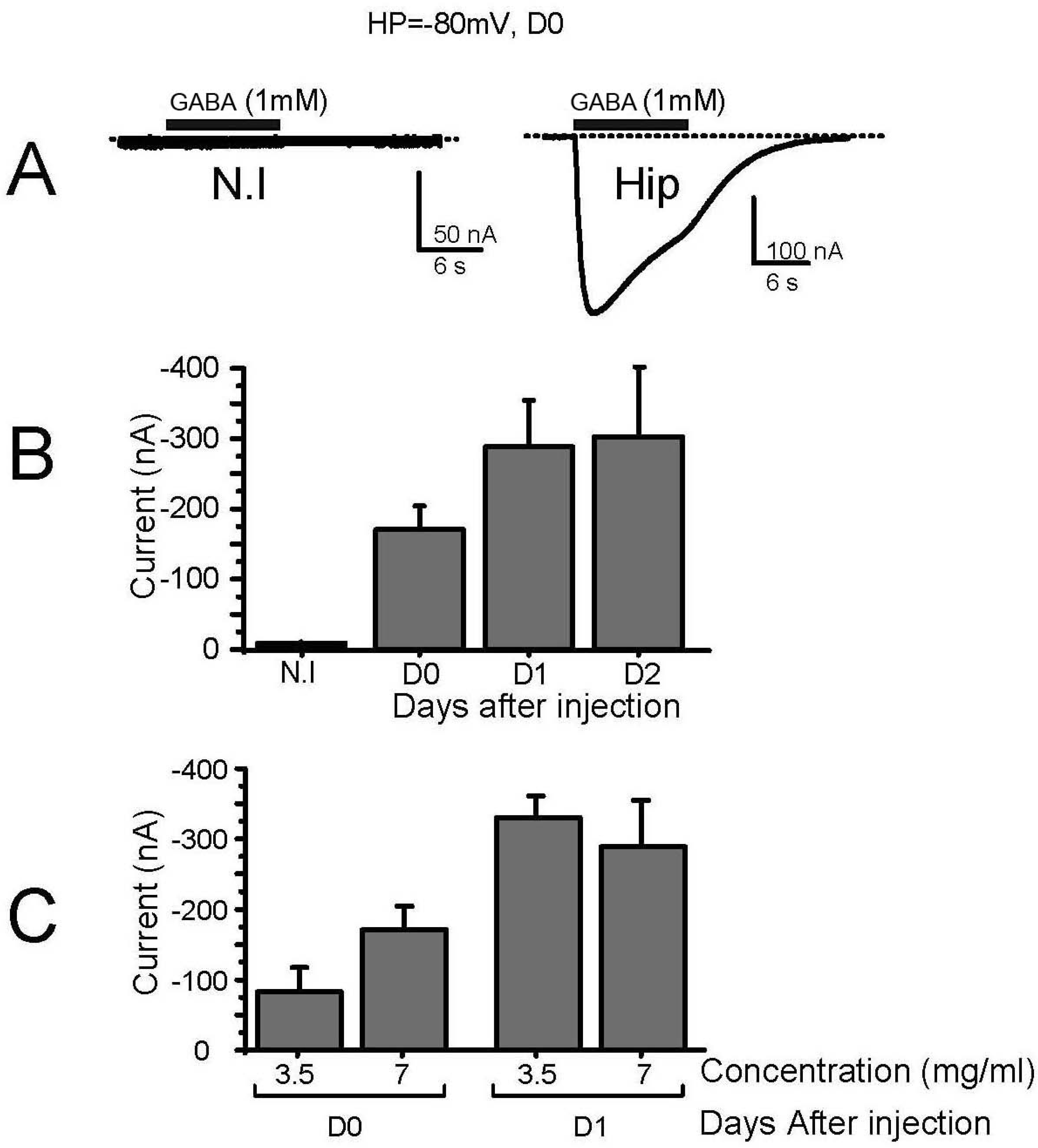
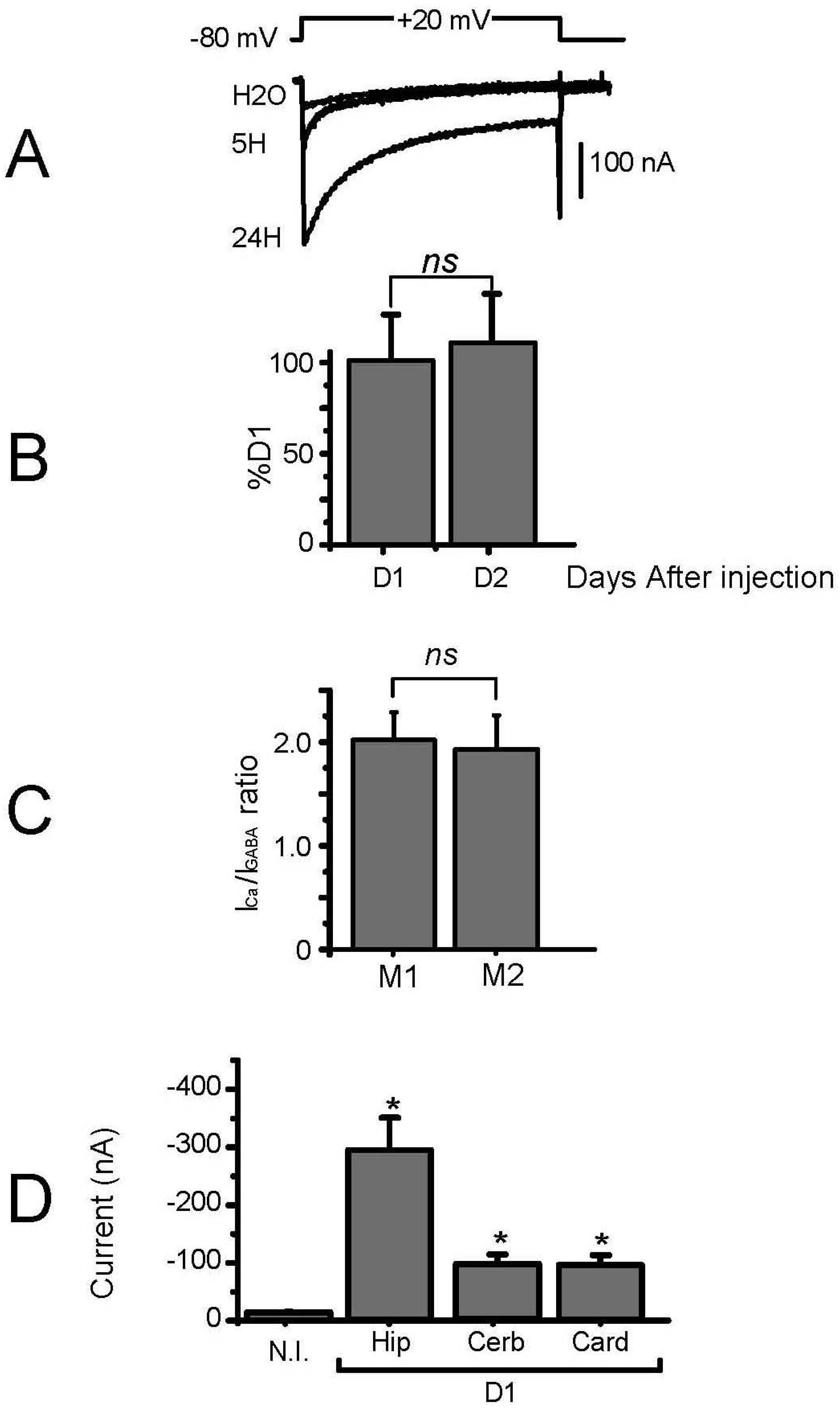
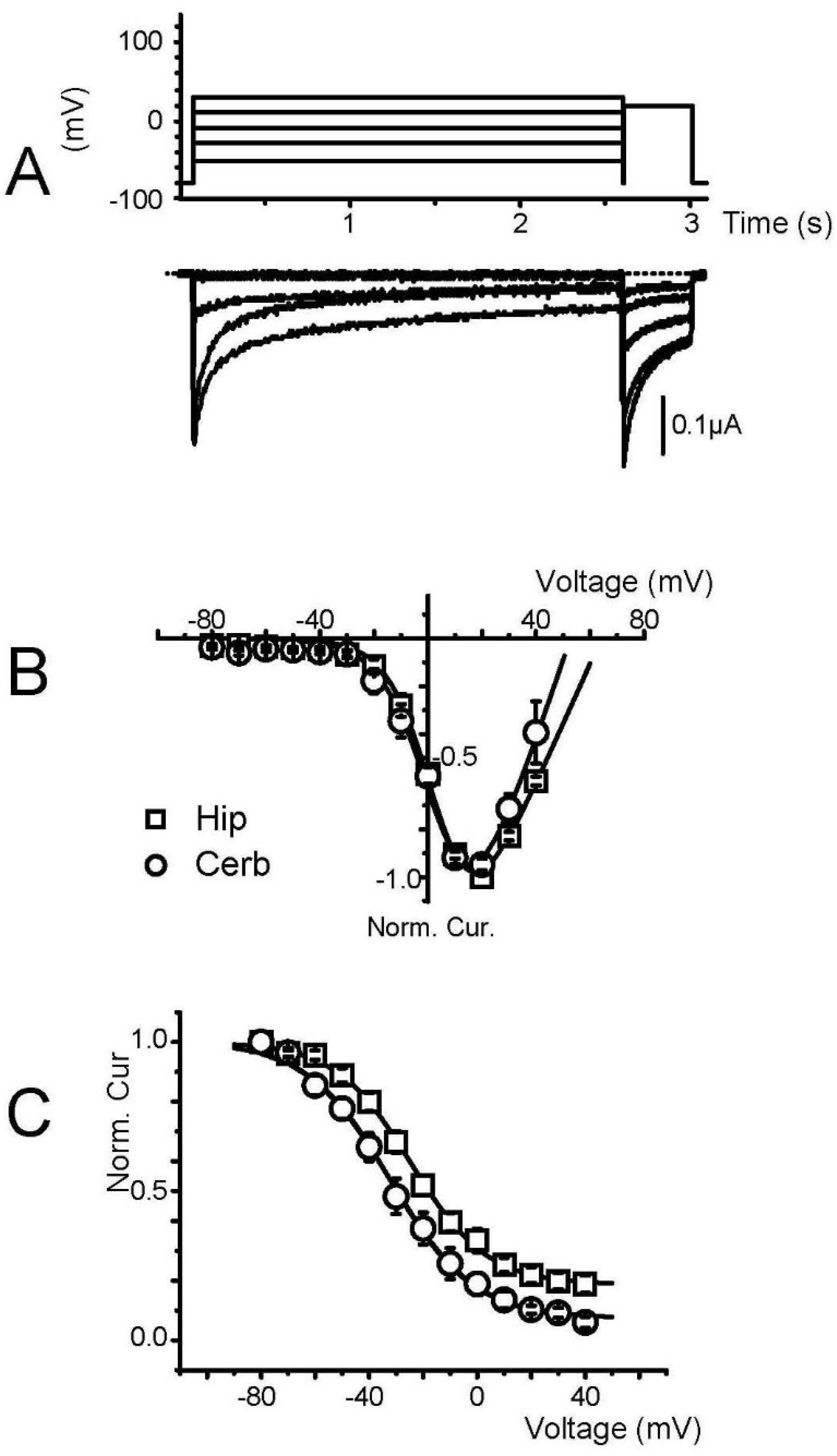
3. Results
3.1. Characterization of Membrane-Transplanted Oocytes
3.2. Expression of Voltage-Gated Ca2+ Channels in Transplanted Oocytes
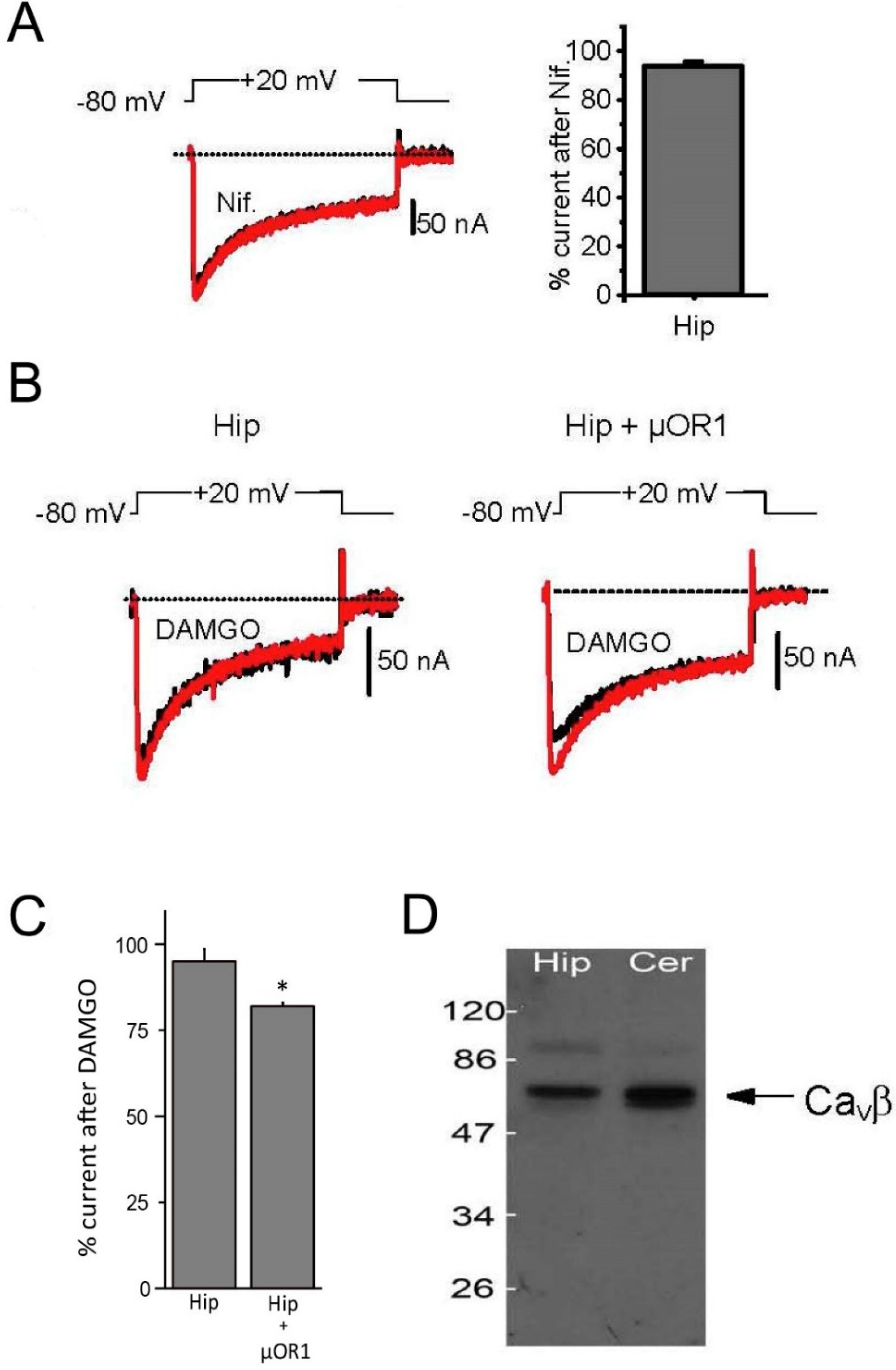
3.3. Biophysical and Pharmacological Properties of Expressed VGCC
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eusebi, F.; Palma, E.; Amici, M.; Miledi, R. Microtransplantation of ligand-gated receptor-channels from fresh or frozen nervous tissue into Xenopus oocytes: A potent tool for expanding functional information. Prog. Neurobiol. 2009, 88, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Miledi, R.; Palma, E.; Eusebi, F. Microtransplantation of neurotransmitter receptors from cells to Xenopus oocyte membranes: New procedure for ion channel studies. Methods Mol. Biol. 2006, 322, 347–355. [Google Scholar] [PubMed]
- Palma, E.; Spinelli, G.; Torchia, G.; Martinez-Torres, A.; Ragozzino, D.; Miledi, R.; Eusebi, F. Abnormal GABAA receptors from the human epileptic hippocampal subiculum microtransplanted to Xenopus oocytes. Proc. Natl. Acad. Sci. USA 2005, 102, 2514–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, E.; Esposito, V.; Mileo, A.M.; Di Gennaro, G.; Quarato, P.; Giangaspero, F.; Scoppetta, C.; Onorati, P.; Trettel, F.; Miledi, R.; et al. Expression of human epileptic temporal lobe neurotransmitter receptors in Xenopus oocytes: An innovative approach to study epilepsy. Proc. Natl. Acad. Sci. USA 2002, 99, 15078–15083. [Google Scholar] [CrossRef] [Green Version]
- Miledi, R.; Eusebi, F.; Martinez-Torres, A.; Palma, E.; Trettel, F. Expression of functional neurotransmitter receptors in Xenopus oocytes after injection of human brain membranes. Proc. Natl. Acad. Sci. USA 2002, 99, 192445299. [Google Scholar] [CrossRef] [Green Version]
- Marsal, J.; Tigyi, G.; Miledi, R. Incorporation of acetylcholine receptors and Cl-channels in Xenopus oocytes injected with Torpedo electroplaque membranes. Proc. Natl. Acad. Sci. USA 1995, 92, 5224–5228. [Google Scholar] [CrossRef] [Green Version]
- Morales, A.; Nguyen, Q.T.; Miledi, R. Electrophysiological properties of newborn and adult rat spinal cord glycine receptors expressed in Xenopus oocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 3097–3101. [Google Scholar] [CrossRef] [Green Version]
- Bernareggi, A.; Reyes-Ruiz, J.M.; Lorenzon, P.; Ruzzier, F.; Miledi, R. Microtransplantation of acetylcholine receptors from normal or denervated rat skeletal muscles to frog oocytes. J. Physiol. 2011, 589, 1133–1142. [Google Scholar] [CrossRef]
- Kepplinger, K.J.; Romanin, C. The run-down phenomenon of Ca2+ channels. In Voltage-Gated Calcium Channels; Zamponi, G.W., Ed.; Eurekah.com at Kluver Academic/Plenum Publishers: New York, NY, USA, 2005; pp. 219–230. [Google Scholar]
- Belles, B.; Malecot, C.O.; Hescheler, J.; Trautwein, W. “Run-down” of the Ca current during long whole-cell recordings in guinea pig heart cells: Role of phosphorylation and intracellular calcium. Pflug. Arch. 1988, 411, 353–360. [Google Scholar] [CrossRef]
- Wu, L.; Bauer, C.S.; Zhen, X.-G.; Xie, C.; Yang, J. Dual regulation of voltage-gated calcium channels by PtdIns (4, 5) P2. Nature 2002, 419, 947–952. [Google Scholar] [CrossRef]
- Restituito, S.; Cens, T.; Barrere, C.; Geib, S.; Galas, S.; De Waard, M.; Charnet, P. The b2a subunit is a molecular groom for the Ca2+ channel inactivation gate. J. Neurosci. 2000, 20, 9046–9052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dascal, N. The Use ofXenopusOocytes for the Study of Ion Channel. Crit. Rev. Biochem. 1987, 22, 317–387. [Google Scholar] [CrossRef] [PubMed]
- Terhag, J.; Cavara, N.A.; Hollmann, M. Cave canalem: How endogenous ion channels may interfere with heterologous expression in Xenopus oocytes. Methods 2010, 51, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Weber, W.M. Ion currents of Xenopus laevis oocytes: State of the art. Biochim. Et Biophys. Acta (BBA)-Biomembr. 1999, 1421, 213–233. [Google Scholar] [CrossRef] [Green Version]
- Charnet, P.; Bourinet, E.; Dubel, S.J.; Snutch, T.P.; Nargeot, J. Calcium currents recorded from a neuronal alpha 1C L-type calcium channel in Xenopus oocytes. FEBS Lett. 1994, 344, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Bourinet, E.; Fournier, F.; Nargeot, J.; Charnet, P. EndogenousXenopus-oocyte Ca-channels are regulated by protein kinases A and C. FEBS Lett. 1992, 299, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Bourinet, E.; Fournier, F.; Lory, P.; Charnet, P.; Nargeot, J. Protein kinase C regulation of cardiac calcium channels expressed in Xenopus oocytes. Pflug. Arch. 1992, 421, 247–255. [Google Scholar] [CrossRef]
- Sun, Y.; Shi, N.; Li, H.; Liu, K.; Zhang, Y.; Chen, W.; Sun, X. Ghrelin suppresses Purkinje neuron P-type Ca(2+) channels via growth hormone secretagogue type 1a receptor, the betagamma subunits of Go-protein, and protein kinase a pathway. Cell. Signal. 2014, 26, 2530–2538. [Google Scholar] [CrossRef]
- Salvi, J.; Bertaso, F.; Mausset-Bonnefont, A.L.; Metz, A.; Lemmers, C.; Ango, F.; Mezghrani, A. RNAi silencing of P/Q-type calcium channels in Purkinje neurons of adult mouse leads to episodic ataxia type 2. Neurobiol. Dis. 2014, 68, 47–56. [Google Scholar] [CrossRef]
- Yan, H.; Pablo, J.L.; Pitt, G.S. FGF14 regulates presynaptic Ca2+ channels and synaptic transmission. Cell. Rep. 2013, 4, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Condliffe, S.B.; Corradini, I.; Pozzi, D.; Verderio, C.; Matteoli, M. Endogenous SNAP-25 regulates native voltage-gated calcium channels in glutamatergic neurons. J. Biol. Chem. 2010, 285, 24968–24976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brittain, J.M.; Piekarz, A.D.; Wang, Y.; Kondo, T.; Cummins, T.R.; Khanna, R. An atypical role for collapsin response mediator protein 2 (CRMP-2) in neurotransmitter release via interaction with presynaptic voltage-gated calcium channels. J. Biol. Chem. 2009, 284, 31375–31390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, F.; Charnet, P.; Bourinet, E.; Vilbert, C.; Matifat, F.; Charpentier, G.; Navarre, P.; Brûlé, G.; Marlot, D. Regulation by protein kinase-C of putative P-type Ca channels expressed in Xenopus oocytes from cerebellar mRNA. FEBS Lett. 1993, 317, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Proft, J.; Weiss, N. G protein regulation of neuronal calcium channels: Back to the future. Mol. Pharmacol. 2014, 87, 890–906. [Google Scholar] [CrossRef] [Green Version]
- Beedle, A.M.; McRory, J.E.; Poirot, O.; Doering, C.J.; Altier, C.; Barrere, C.; Hamid, J.; Nargeot, J.; Bourinet, E.; Zamponi, G.W. Agonist-independent modulation of N-type calcium channels by ORL1 receptors. Nat. Neurosci. 2004, 7, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Rousset, M.; Cens, T.; Gouin-Charnet, A.; Scamps, F.; Charnet, F. Ca2+ and phosphatidylinositol 4,5-bisphosphate stabilize a G{beta}{gamma}-sensitive state of CaV2 Ca2+ channels. J. Biol. Chem. 2004, 279, 14619–14630. [Google Scholar] [CrossRef] [Green Version]
- Lambert, A.N.; Wilson, A.W. High-threshold Ca2+ currents in rat hippocampal interneurones and their selective inhibition by activation of GABA(B) receptors. J. Physiol. 1996, 492, 115–127. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rousset, M.; Humez, S.; Laurent, C.; Buée, L.; Blum, D.; Cens, T.; Vignes, M.; Charnet, P. Mammalian Brain Ca2+ Channel Activity Transplanted into Xenopus laevis Oocytes. Membranes 2022, 12, 496. https://doi.org/10.3390/membranes12050496
Rousset M, Humez S, Laurent C, Buée L, Blum D, Cens T, Vignes M, Charnet P. Mammalian Brain Ca2+ Channel Activity Transplanted into Xenopus laevis Oocytes. Membranes. 2022; 12(5):496. https://doi.org/10.3390/membranes12050496
Chicago/Turabian StyleRousset, Matthieu, Sandrine Humez, Cyril Laurent, Luc Buée, David Blum, Thierry Cens, Michel Vignes, and Pierre Charnet. 2022. "Mammalian Brain Ca2+ Channel Activity Transplanted into Xenopus laevis Oocytes" Membranes 12, no. 5: 496. https://doi.org/10.3390/membranes12050496
APA StyleRousset, M., Humez, S., Laurent, C., Buée, L., Blum, D., Cens, T., Vignes, M., & Charnet, P. (2022). Mammalian Brain Ca2+ Channel Activity Transplanted into Xenopus laevis Oocytes. Membranes, 12(5), 496. https://doi.org/10.3390/membranes12050496