
Deciphering the Assembly of Enveloped Viruses Using Model Lipid Membranes


{kind=link}
{kind=link}
{kind=link}
Abstract
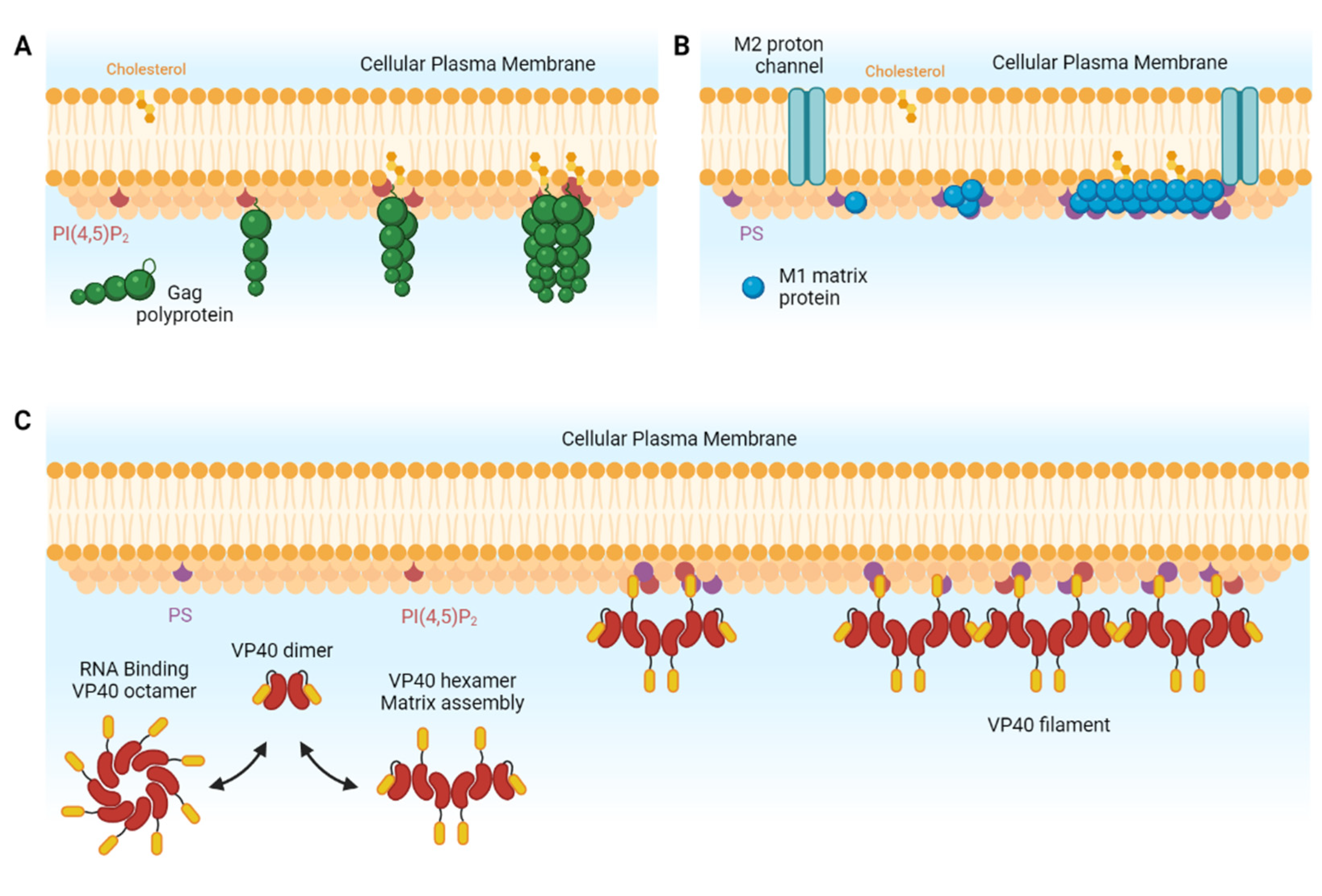
1. Introduction
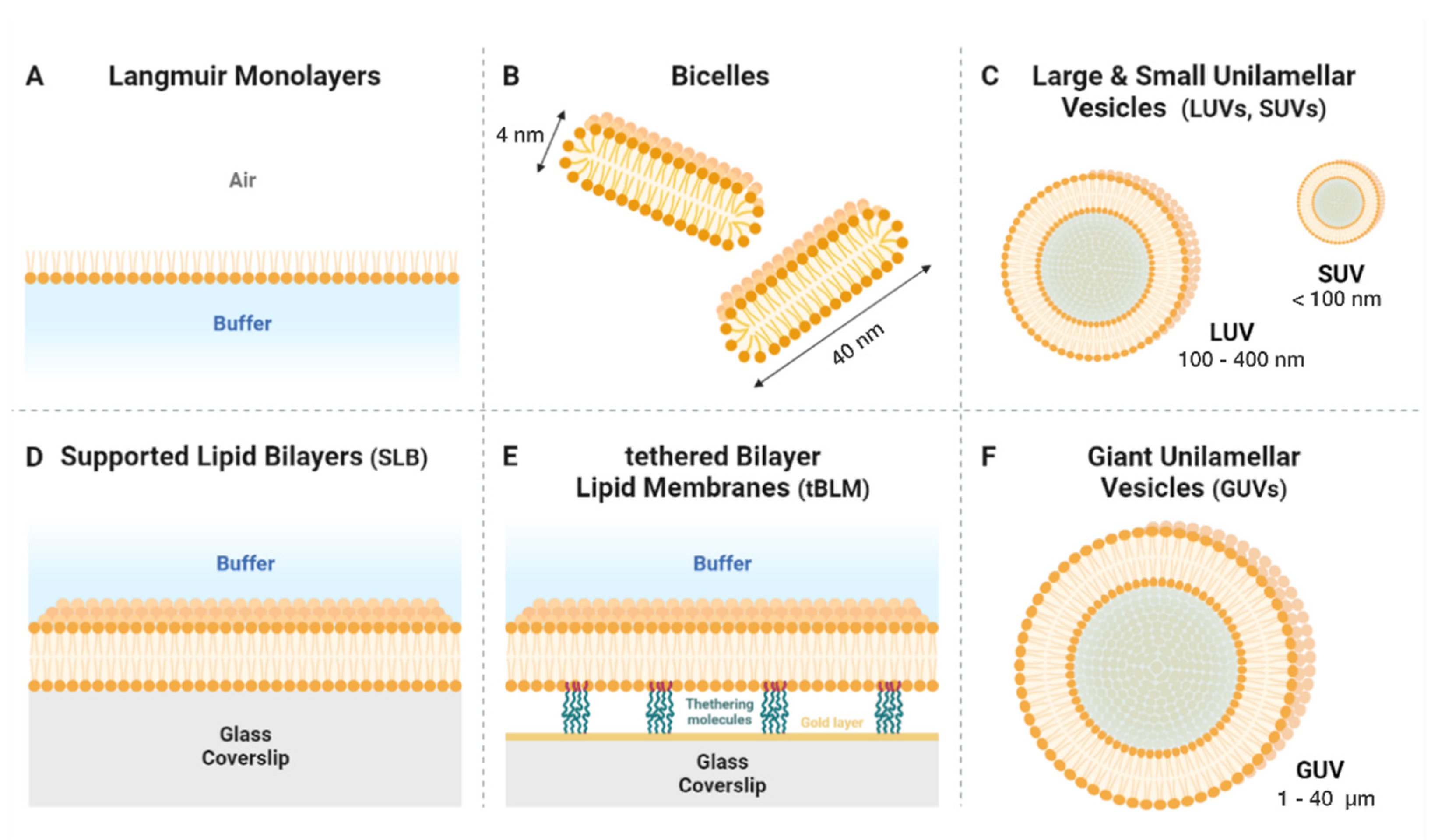
2. An Overview of the Main Types of Model Lipid Membrane
2.1. Langmuir Films
2.2. Micelles and Bicelles
2.3. Unilamellar Vesicles
2.4. Supported Lipid Bilayers
2.5. Giant Unilamellar Vesicles
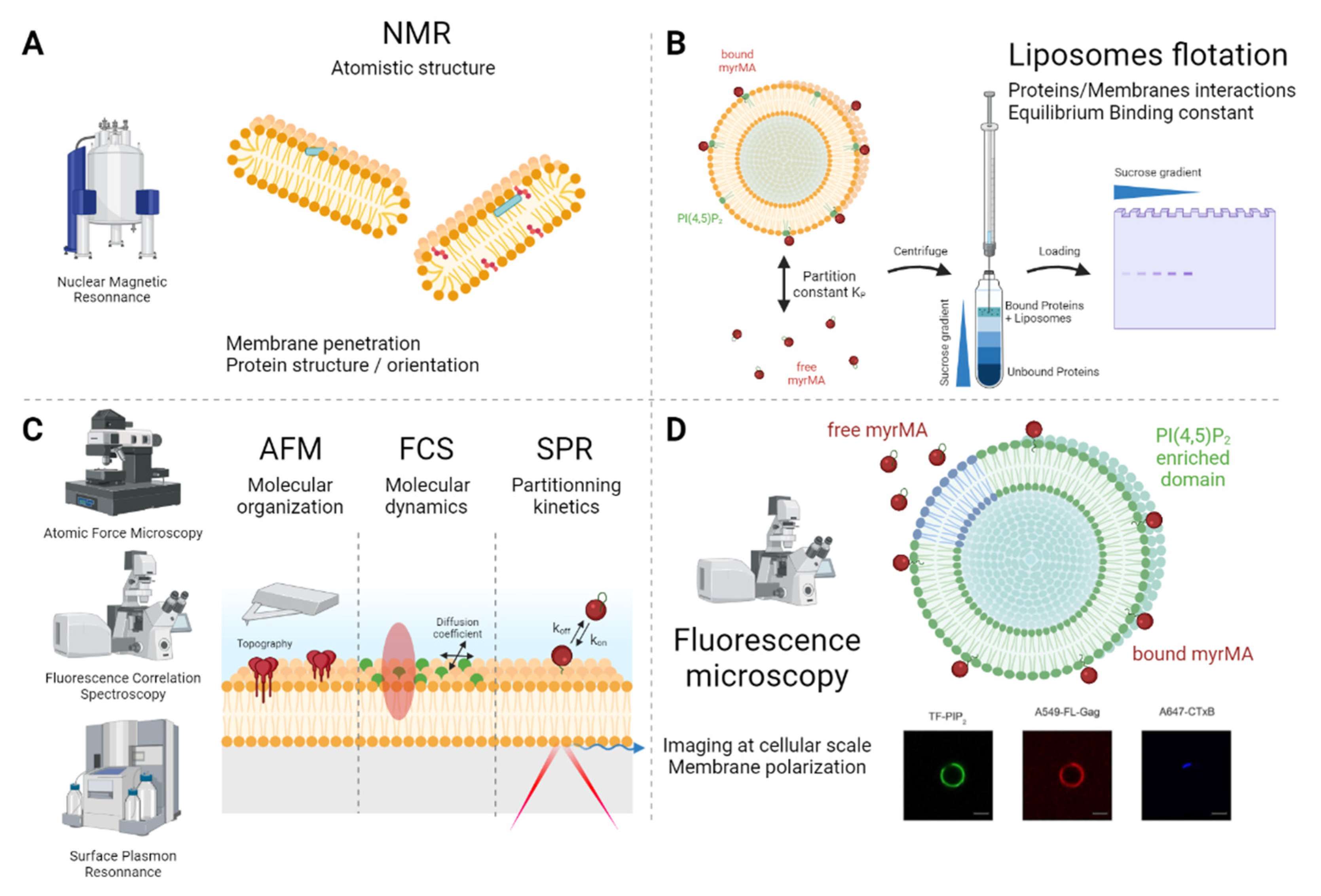
3. Artificial Lipid Membranes as Tools for Viral Assembly Research
3.1. Langmuir Monolayers: A Fine Tuning System
3.2. Using Bicelles to Elucidate the Molecular Structures of Viral Proteins on Membranes
3.3. Unilamellar Vesicles
3.4. Supported and Tethered Bilayers: Planar Membranes
3.5. Giant Unilamellar Vesicles: The Closest Model to Cell Plasma Membranes
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Glossary
| Biophysical Techniques Lexicon | ||
| HX MS | Hydrogen Exchange Mass Spectrometry | This technique exploits proteins’ fundamental ability to exchange hydrogens with their environment. Studied proteins are incorporated in deuterated water (D2O) and exchange hydrogens bonded to the backbone nitrogen with deuterium from D2O solvent over time. Incorporated deuterium atoms are consecutively used to probe protein conformation using mass spectrometry [83]. |
| X-ray Crystallography | The diffraction of incident X-rays by electrons in a sample is used to calculate the positions of atoms in molecules. Due to the fact that the signal of each individual atom is weak, this technique requires sample crystallization (all the molecules are immobilized in the same conformation in a 3D lattice) [84]. | |
| Cryo-EM | Cryo-electron microscopy | This process involves freezing samples at cryogenic temperatures so quickly that water molecules do not crystallize, which preserves the samples’ native structure. This sample processing technique is generally associated with transmission electron microscopy for structural analyses at an atomic level [85]. |
| NMR | Nuclear Magnetic Resonance | NMR is based on the spin properties of atomic nuclei, which are aligned when submitted to a magnetic field. Only a few isotopes that have a magnetic spin are used for NMR purposes (the most common is 1H, but 13C, 15N and 31P are also often used), and the analysis either relies on the natural isotopic proportion in the sample or is performed on enriched samples. Once irradiated, each non-equivalent nucleus resonates at a specific frequency, making peaks in the NMR signal, called chemical shifts. The peaks’ intensity and multiplicity are used to determine the atoms’ spatial proximity [86]. |
| OMAS-NMR | Off-Magic-Angle Spinning NMR | This is a variant of the magic-angle-pinning solid-state NMR developed to narrow the quadrupolar resonances and increase the frequency resolution/attribution. Resonance widening occurs in anisotropic systems, such as proteins bound to lipid membranes. OMAS is based on a slight deviation from the magic angle (angle < 0.01°). This significantly improves the precision and accuracy of derived rate parameters, especially for slow motion on the kilohertz time scale [87]. |
| FCS | Fluorescence Correlation Spectroscopy (STED, scanning, Spot Variation) | This microscopy technique relies on the use of fluorescently labelled particles and makes it possible to quantify their dynamics (including their concentration, diffusion coefficient and interactions) by recording and correlating the fluorescence intensity’s fluctuation over time in a given zone [88]. Using an optical microscope, a scanning mode can be added to perform scanning-FCS. An adjustable pinhole enables spot-variation-FCS. Coupled with a STED microscope, the reduced volume of detection makes it possible to study diffusion in nanoscale regions [89,90,91]. |
| STED | Stimulated Emission Depletion | This is a super-resolution fluorescence microscopy technique based on the possibility to deplete the excited fluorophore before it emits. For this, a depletion laser (emitting at a higher wavelength than the excitation wavelength) is focused on the object plane, as a a donut shape surrounding the excitation laser beam. The fluorophore exposed to this donut shape laser returns to the ground state before emitting a fluorescence photon. This result in a continuous decreasing size of the emission spot below the diffraction limit (<200 nm)with increasing depletion laser power [92]. |
| Liposome flotation assay | This method is used to characterize the interactions between proteins and membrane-mimicking liposomes (LUV). This assay is based on bound/free proteins’ separation by centrifuging the complex through a sucrose gradient. At the appropriate centrifugation speed, proteins bound to liposomes float up to the low-sucrose fraction, whereas denser free proteins remain at the bottom at high sucrose-density levels. Due to protein–LUV interactions, an apparent KD value for the liposome–protein interaction can be estimated by varying the amount of LUV at a constant protein concentration [93,94]. | |
| SPR | Surface Plasmon Resonance | SPR describes a propagation phenomenon of electrons parallel to a metal layer when excited by an incident laser at a certain angle (with the angle depending on the refractive index of the material near the metal surface). An SPR assay is based on the lowered SPR occurrence due to an analyte interaction with its target, coated at the surface of the metal layer. The amount of interacting analytes and the interaction kinetics are unveiled either by measuring the reflected light intensity or by tracking the resonance angle shift [95]. |
| RICS | Raster Image Correlation Spectroscopy | Raster image correlation spectroscopy (RICS) is a fluorescent-image-analysis method for extracting the mobility, concentration and stoichiometry of diffusing fluorescent molecules from confocal image stacks. The method works by calculating a spatial correlation function for each image and analysing their average by model fitting [96]. |
| CD Spectroscopy | Circular Dichroism Spectroscopy | Polarized absorption spectroscopy method. When illuminated by an elliptical polarized light, the absorption of the incident light will depend on chirality, but also on the folding of the structures in the case of complex macromolecules. It is generally used to characterize the conformation of a protein’s secondary structure and, in some cases, of its tertiary structure environment. It also enables access to dynamic conformational changes in proteins in or at membranes [97]. |
References
- Bernardino de la Serna, J.; Schütz, G.J.; Eggeling, C.; Cebecauer, M. There Is No Simple Model of the Plasma Membrane Organization. Front. Cell Dev. Biol. 2016, 4, 106. [Google Scholar] [CrossRef] [PubMed]
- Jouvenet, N.; Bieniasz, P.D.; Simon, S.M. Imaging the Biogenesis of Individual HIV-1 Virions in Live Cells. Nature 2008, 454, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Floderer, C.; Masson, J.-B.; Boilley, E.; Georgeault, S.; Merida, P.; El Beheiry, M.; Dahan, M.; Roingeard, P.; Sibarita, J.-B.; Favard, C.; et al. Single Molecule Localisation Microscopy Reveals How HIV-1 Gag Proteins Sense Membrane Virus Assembly Sites in Living Host CD4 T Cells. Sci. Rep. 2018, 8, 16283. [Google Scholar] [CrossRef]
- Michaels, T.C.T.; Bellaiche, M.M.J.; Hagan, M.F.; Knowles, T.P.J. Kinetic Constraints on Self-Assembly into Closed Supramolecular Structures. Sci. Rep. 2017, 7, 12295. [Google Scholar] [CrossRef] [PubMed]
- Saad, J.S.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural Basis for Targeting HIV-1 Gag Proteins to the Plasma Membrane for Virus Assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 11364–11369. [Google Scholar] [CrossRef]
- Saad, J.S.; Muriaux, D.M. Editorial: Role of Lipids in Virus Assembly. Front. Microbiol. 2015, 6, 410. [Google Scholar] [CrossRef]
- Muriaux, D.; Darlix, J.-L. Properties and Functions of the Nucleocapsid Protein in Virus Assembly. RNA Biol. 2010, 7, 744–753. [Google Scholar] [CrossRef]
- Kutluay, S.B.; Bieniasz, P.D. Analysis of the Initiating Events in HIV-1 Particle Assembly and Genome Packaging. PLoS Pathog. 2010, 6, e1001200. [Google Scholar] [CrossRef]
- Rossman, J.S.; Lamb, R.A. Influenza Virus Assembly and Budding. Virology 2011, 411, 229–236. [Google Scholar] [CrossRef]
- Motsa, B.B.; Stahelin, R.V. Lipid–Protein Interactions in Virus Assembly and Budding from the Host Cell Plasma Membrane. Biochem. Soc. Trans. 2021, 49, 1633–1641. [Google Scholar] [CrossRef]
- Panahandeh, S.; Li, S.; Marichal, L.; Leite Rubim, R.; Tresset, G.; Zandi, R. How a Virus Circumvents Energy Barriers to Form Symmetric Shells. ACS Nano 2020, 14, 3170–3180. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Shklovskii, B.I. Kinetics of Macroion Coagulation Induced by Multivalent Counterions. Phys. Rev. E 2002, 65, 031409. [Google Scholar] [CrossRef] [PubMed]
- Hogue, I.B.; Hoppe, A.; Ono, A. Quantitative Fluorescence Resonance Energy Transfer Microscopy Analysis of the Human Immunodeficiency Virus Type 1 Gag-Gag Interaction: Relative Contributions of the CA and NC Domains and Membrane Binding. J. Virol. 2009, 83, 7322–7336. [Google Scholar] [CrossRef] [PubMed]
- Gamble, T.R.; Yoo, S.; Vajdos, F.F.; von Schwedler, U.K.; Worthylake, D.K.; Wang, H.; McCutcheon, J.P.; Sundquist, W.I.; Hill, C.P. Structure of the Carboxyl-Terminal Dimerization Domain of the HIV-1 Capsid Protein. Science 1997, 278, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Hilsch, M.; Goldenbogen, B.; Sieben, C.; Höfer, C.T.; Rabe, J.P.; Klipp, E.; Herrmann, A.; Chiantia, S. Influenza A Matrix Protein M1 Multimerizes upon Binding to Lipid Membranes. Biophys. J. 2014, 107, 912–923. [Google Scholar] [CrossRef]
- Kerviel, A.; Dash, S.; Moncorgé, O.; Panthu, B.; Prchal, J.; Décimo, D.; Ohlmann, T.; Lina, B.; Favard, C.; Decroly, E.; et al. Involvement of an Arginine Triplet in M1 Matrix Protein Interaction with Membranes and in M1 Recruitment into Virus-Like Particles of the Influenza A(H1N1)Pdm09 Virus. PLoS ONE 2016, 11, e0165421. [Google Scholar] [CrossRef]
- Petrich, A.; Dunsing, V.; Bobone, S.; Chiantia, S. Influenza A M2 Recruits M1 to the Plasma Membrane: A Fluorescence Fluctuation Microscopy Study. Biophys. J. 2021, 120, 5478–5490. [Google Scholar] [CrossRef]
- Johnson, K.A.; Taghon, G.J.F.; Scott, J.L.; Stahelin, R.V. The Ebola Virus Matrix Protein, VP40, Requires Phosphatidylinositol 4,5-Bisphosphate (PI(4,5)P2) for Extensive Oligomerization at the Plasma Membrane and Viral Egress. Sci. Rep. 2016, 6, 19125. [Google Scholar] [CrossRef]
- Kielian, M.; Rey, F.A. Virus Membrane-Fusion Proteins: More than One Way to Make a Hairpin. Nat. Rev. Microbiol. 2006, 4, 67–76. [Google Scholar] [CrossRef]
- Más, V.; Melero, J.A. Entry of Enveloped Viruses into Host Cells: Membrane Fusion. In Structure and Physics of Viruses; Mateu, M.G., Ed.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2013; Volume 68, pp. 467–487. ISBN 978-94-007-6551-1. [Google Scholar]
- Mouritsen, O.G. Model Answers to Lipid Membrane Questions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004622. [Google Scholar] [CrossRef]
- Zasadzinski, J.A.; Viswanathan, R.; Madsen, L.; Garnaes, J.; Schwartz, D.K. Langmuir-Blodgett Films. Science 1994, 263, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.R.; Prosser, R.S. Bicelles: A Model Membrane System for All Seasons? Structure 1998, 6, 1227–1234. [Google Scholar] [CrossRef]
- Hutchison, J.M.; Shih, K.-C.; Scheidt, H.A.; Fantin, S.M.; Parson, K.F.; Pantelopulos, G.A.; Harrington, H.R.; Mittendorf, K.F.; Qian, S.; Stein, R.A.; et al. Bicelles Rich in Both Sphingolipids and Cholesterol and Their Use in Studies of Membrane Proteins. J. Am. Chem. Soc. 2020, 142, 12715–12729. [Google Scholar] [CrossRef]
- Schwendener, R.A. Liposomes in Biology and Medicine. In Bio-Applications of Nanoparticles; Chan, W.C.W., Ed.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2007; Volume 620, pp. 117–128. ISBN 978-0-387-76712-3. [Google Scholar]
- Tamm, L.K.; McConnell, H.M. Supported Phospholipid Bilayers. Biophys. J. 1985, 47, 105–113. [Google Scholar] [CrossRef]
- Veneziano, R.; Rossi, C.; Chenal, A.; Brenner, C.; Ladant, D.; Chopineau, J. Synthesis and Characterization of Tethered Lipid Assemblies for Membrane Protein Reconstitution (Review). Biointerphases 2017, 12, 04E301. [Google Scholar] [CrossRef] [PubMed]
- Reeves, J.P.; Dowben, R.M. Formation and Properties of Thin-Walled Phospholipid Vesicles. J. Cell. Physiol. 1969, 73, 49–60. [Google Scholar] [CrossRef]
- Weinberger, A.; Tsai, F.-C.; Koenderink, G.H.; Schmidt, T.F.; Itri, R.; Meier, W.; Schmatko, T.; Schröder, A.; Marques, C. Gel-Assisted Formation of Giant Unilamellar Vesicles. Biophys. J. 2013, 105, 154–164. [Google Scholar] [CrossRef]
- Angelova, M.I.; Dimitrov, D.S. Liposome Electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303. [Google Scholar] [CrossRef]
- Chiantia, S.; Schwille, P.; Klymchenko, A.S.; London, E. Asymmetric GUVs Prepared by MβCD-Mediated Lipid Exchange: An FCS Study. Biophys. J. 2011, 100, L1–L3. [Google Scholar] [CrossRef]
- Enoki, T.A.; Wu, J.; Heberle, F.A.; Feigenson, G.W. Investigation of the Domain Line Tension in Asymmetric Vesicles Prepared via Hemifusion. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183586. [Google Scholar] [CrossRef]
- Malbec, M.; Sourisseau, M.; Guivel-Benhassine, F.; Porrot, F.; Blanchet, F.; Schwartz, O.; Casartelli, N. HIV-1 Nef Promotes the Localization of Gag to the Cell Membrane and Facilitates Viral Cell-to-Cell Transfer. Retrovirology 2013, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Pirrone, G.F.; Emert-Sedlak, L.A.; Wales, T.E.; Smithgall, T.E.; Kent, M.S.; Engen, J.R. Membrane-Associated Conformation of HIV-1 Nef Investigated with Hydrogen Exchange Mass Spectrometry at a Langmuir Monolayer. Anal. Chem. 2015, 87, 7030–7035. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, E.; Soni, S.P.; Xue, Y.; Digman, M.A.; Gratton, E.; Stahelin, R.V. The Ebola Virus Matrix Protein Penetrates into the Plasma Membrane. J. Biol. Chem. 2013, 288, 5779–5789. [Google Scholar] [CrossRef]
- Kerviel, A.; Thomas, A.; Chaloin, L.; Favard, C.; Muriaux, D. Virus Assembly and Plasma Membrane Domains: Which Came First? Virus Res. 2013, 171, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Vlach, J.; Saad, J.S. Trio Engagement via Plasma Membrane Phospholipids and the Myristoyl Moiety Governs HIV-1 Matrix Binding to Bilayers. Proc. Natl. Acad. Sci. USA 2013, 110, 3525–3530. [Google Scholar] [CrossRef] [PubMed]
- Mercredi, P.Y.; Bucca, N.; Loeliger, B.; Gaines, C.R.; Mehta, M.; Bhargava, P.; Tedbury, P.R.; Charlier, L.; Floquet, N.; Muriaux, D.; et al. Structural and Molecular Determinants of Membrane Binding by the HIV-1 Matrix Protein. J. Mol. Biol. 2016, 428, 1637–1655. [Google Scholar] [CrossRef] [PubMed]
- Charlier, L.; Louet, M.; Chaloin, L.; Fuchs, P.; Martinez, J.; Muriaux, D.; Favard, C.; Floquet, N. Coarse-Grained Simulations of the HIV-1 Matrix Protein Anchoring: Revisiting Its Assembly on Membrane Domains. Biophys. J. 2014, 106, 577–585. [Google Scholar] [CrossRef]
- Wang, T.; Hong, M. Investigation of the Curvature Induction and Membrane Localization of the Influenza Virus M2 Protein Using Static and Off-Magic-Angle Spinning Solid-State Nuclear Magnetic Resonance of Oriented Bicelles. Biochemistry 2015, 54, 2214–2226. [Google Scholar] [CrossRef]
- Zhou, W.; Resh, M.D. Differential Membrane Binding of the Human Immunodeficiency Virus Type 1 Matrix Protein. J. Virol. 1996, 70, 8540–8548. [Google Scholar] [CrossRef]
- Dalton, A.K.; Ako-Adjei, D.; Murray, P.S.; Murray, D.; Vogt, V.M. Electrostatic Interactions Drive Membrane Association of the Human Immunodeficiency Virus Type 1 Gag MA Domain. J. Virol. 2007, 81, 6434–6445. [Google Scholar] [CrossRef]
- Ehrlich, L.S.; Fong, S.; Scarlata, S.; Zybarth, G.; Carter, C. Partitioning of HIV-1 Gag and Gag-Related Proteins to Membranes. Biochemistry 1996, 35, 3933–3943. [Google Scholar] [CrossRef] [PubMed]
- Yandrapalli, N.; Lubart, Q.; Tanwar, H.S.; Picart, C.; Mak, J.; Muriaux, D.; Favard, C. Self Assembly of HIV-1 Gag Protein on Lipid Membranes Generates PI(4,5)P2/Cholesterol Nanoclusters. Sci. Rep. 2016, 6, 39332. [Google Scholar] [CrossRef] [PubMed]
- Dick, R.A.; Goh, S.L.; Feigenson, G.W.; Vogt, V.M. HIV-1 Gag Protein Can Sense the Cholesterol and Acyl Chain Environment in Model Membranes. Proc. Natl. Acad. Sci. USA 2012, 109, 18761–18766. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, V.; Hogue, I.B.; Boyko, V.; Hu, W.-S.; Ono, A. Interaction between the Human Immunodeficiency Virus Type 1 Gag Matrix Domain and Phosphatidylinositol-(4,5)-Bisphosphate Is Essential for Efficient Gag Membrane Binding. J. Virol. 2008, 82, 2405–2417. [Google Scholar] [CrossRef] [PubMed]
- Bouamr, F.; Scarlata, S.; Carter, C. Role of Myristylation in HIV-1 Gag Assembly. Biochemistry 2003, 42, 6408–6417. [Google Scholar] [CrossRef]
- Pérez Socas, L.B.; Ambroggio, E.E. The Influence of Myristoylation, Liposome Surface Charge and Nucleic Acid Interaction in the Partition Properties of HIV-1 Gag-N-Terminal Peptides to Membranes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183421. [Google Scholar] [CrossRef]
- Chukkapalli, V.; Oh, S.J.; Ono, A. Opposing Mechanisms Involving RNA and Lipids Regulate HIV-1 Gag Membrane Binding through the Highly Basic Region of the Matrix Domain. Proc. Natl. Acad. Sci. USA 2010, 107, 1600–1605. [Google Scholar] [CrossRef]
- Sumner, C.; Kotani, O.; Liu, S.; Musier-Forsyth, K.; Sato, H.; Ono, A. Molecular Determinants in TRNA D-Arm Required for Inhibition of HIV-1 Gag Membrane Binding. J. Mol. Biol. 2022, 434, 167390. [Google Scholar] [CrossRef]
- Todd, G.C.; Duchon, A.; Inlora, J.; Olson, E.D.; Musier-Forsyth, K.; Ono, A. Inhibition of HIV-1 Gag–Membrane Interactions by Specific RNAs. RNA 2017, 23, 395–405. [Google Scholar] [CrossRef]
- Keller, H.; Kräusslich, H.-G.; Schwille, P. Multimerizable HIV Gag Derivative Binds to the Liquid-Disordered Phase in Model Membranes. Cell. Microbiol. 2013, 15, 237–247. [Google Scholar] [CrossRef]
- Urbančič, I.; Brun, J.; Shrestha, D.; Waithe, D.; Eggeling, C.; Chojnacki, J. Lipid Composition but Not Curvature Is the Determinant Factor for the Low Molecular Mobility Observed on the Membrane of Virus-Like Vesicles. Viruses 2018, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Gregoriades, A. Interaction of Influenza M Protein with Viral Lipid and Phosphatidylcholine Vesicles. J. Virol. 1980, 36, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Oxford, J.S.; Hockley, D.J.; Heath, T.D.; Patterson, S. The Interaction of Influenza Virus Haemagglutinin with Phospholipid Vesicles—Morphological and Immunological Studies. J. Gen. Virol. 1981, 52, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Baudin, F.; Petit, I.; Weissenhorn, W.; Ruigrok, R.W. In Vitro Dissection of the Membrane and RNP Binding Activities of Influenza Virus M1 Protein. Virology 2001, 281, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.W.; Barge, A.; Durrer, P.; Brunner, J.; Ma, K.; Whittaker, G.R. Membrane Interaction of Influenza Virus M1 Protein. Virology 2000, 267, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Kordyukova, L.V.; Konarev, P.V.; Fedorova, N.V.; Shtykova, E.V.; Ksenofontov, A.L.; Loshkarev, N.A.; Dadinova, L.A.; Timofeeva, T.A.; Abramchuk, S.S.; Moiseenko, A.V.; et al. The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer. Membranes 2021, 11, 772. [Google Scholar] [CrossRef]
- Huang, R.T.C.; Warn, K.; Klenk, H.-D.; Rott, R. Association of the Envelope Glycoproteins of Influenza Virus with Liposomes—a Model Study on Viral Envelope Assembly. Virology 1979, 97, 212–217. [Google Scholar] [CrossRef]
- Bailey, A.; Zhukovsky, M.; Gliozzi, A.; Chernomordik, L.V. Liposome Composition Effects on Lipid Mixing between Cells Expressing Influenza Virus Hemagglutinin and Bound Liposomes. Arch. Biochem. Biophys. 2005, 439, 211–221. [Google Scholar] [CrossRef]
- Rossman, J.S.; Jing, X.; Leser, G.P.; Lamb, R.A. Influenza Virus M2 Protein Mediates ESCRT-Independent Membrane Scission. Cell 2010, 142, 902–913. [Google Scholar] [CrossRef]
- Martyna, A.; Gómez-Llobregat, J.; Lindén, M.; Rossman, J.S. Curvature Sensing by a Viral Scission Protein. Biochemistry 2016, 55, 3493–3496. [Google Scholar] [CrossRef]
- Martyna, A.; Bahsoun, B.; Madsen, J.J.; Jackson, F.S.J.S.; Badham, M.D.; Voth, G.A.; Rossman, J.S. Cholesterol Alters the Orientation and Activity of the Influenza Virus M2 Amphipathic Helix in the Membrane. J. Phys. Chem. B 2020, 124, 6738–6747. [Google Scholar] [CrossRef] [PubMed]
- Ruigrok, R.W.H.; Schoehn, G.; Dessen, A.; Forest, E.; Volchkov, V.; Dolnik, O.; Klenk, H.-D.; Weissenhorn, W. Structural Characterization and Membrane Binding Properties of the Matrix Protein VP40 of Ebola Virus11Edited by J. Karn. J. Mol. Biol. 2000, 300, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Scianimanico, S. Membrane Association Induces a Conformational Change in the Ebola Virus Matrix Protein. EMBO J. 2000, 19, 6732–6741. [Google Scholar] [CrossRef] [PubMed]
- Valbuena, A.; Maity, S.; Mateu, M.G.; Roos, W.H. Visualization of Single Molecules Building a Viral Capsid Protein Lattice through Stochastic Pathways. ACS Nano 2020, 14, 8724–8734. [Google Scholar] [CrossRef] [PubMed]
- Favard, C.; Chojnacki, J.; Merida, P.; Yandrapalli, N.; Mak, J.; Eggeling, C.; Muriaux, D. HIV-1 Gag Specifically Restricts PI(4,5)P2 and Cholesterol Mobility in Living Cells Creating a Nanodomain Platform for Virus Assembly. Sci. Adv. 2019, 5, eaaw8651. [Google Scholar] [CrossRef] [PubMed]
- Miles, P.; Cassidy, P.; Donlon, L.; Yarkoni, O.; Frankel, D. In Vitro Assembly of a Viral Envelope. Soft Matter 2015, 11, 7722–7727. [Google Scholar] [CrossRef] [PubMed]
- Bobone, S.; Hilsch, M.; Storm, J.; Dunsing, V.; Herrmann, A.; Chiantia, S. Phosphatidylserine Lateral Organization Influences the Interaction of Influenza Virus Matrix Protein 1 with Lipid Membranes. J. Virol. 2017, 91, e00267-17. [Google Scholar] [CrossRef]
- Höfer, C.T.; Di Lella, S.; Dahmani, I.; Jungnick, N.; Bordag, N.; Bobone, S.; Huang, Q.; Keller, S.; Herrmann, A.; Chiantia, S. Structural Determinants of the Interaction between Influenza A Virus Matrix Protein M1 and Lipid Membranes. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1123–1134. [Google Scholar] [CrossRef]
- Cremer, P.S.; Boxer, S.G. Formation and Spreading of Lipid Bilayers on Planar Glass Supports. J. Phys. Chem. B 1999, 103, 2554–2559. [Google Scholar] [CrossRef]
- Wen, Y.; Feigenson, G.W.; Vogt, V.M.; Dick, R.A. Mechanisms of PI(4,5)P2 Enrichment in HIV-1 Viral Membranes. J. Mol. Biol. 2020, 432, 5343–5364. [Google Scholar] [CrossRef]
- Gui, D.; Gupta, S.; Xu, J.; Zandi, R.; Gill, S.; Huang, I.-C.; Rao, A.L.N.; Mohideen, U. A Novel Minimal in Vitro System for Analyzing HIV-1 Gag-Mediated Budding. J. Biol. Phys. 2015, 41, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Van Engelenburg, S.B.; Shtengel, G.; Sengupta, P.; Waki, K.; Jarnik, M.; Ablan, S.D.; Freed, E.O.; Hess, H.F.; Lippincott-Schwartz, J. Distribution of ESCRT Machinery at HIV Assembly Sites Reveals Virus Scaffolding of ESCRT Subunits. Science 2014, 343, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Carlson, L.-A.; Hurley, J.H. In Vitro Reconstitution of the Ordered Assembly of the Endosomal Sorting Complex Required for Transport at Membrane-Bound HIV-1 Gag Clusters. Proc. Natl. Acad. Sci. USA 2012, 109, 16928–16933. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, I.; Ludwig, K.; Chiantia, S. Influenza A Matrix Protein M1 Induces Lipid Membrane Deformation via Protein Multimerization. Biosci. Rep. 2019, 39, BSR20191024. [Google Scholar] [CrossRef] [PubMed]
- Saletti, D.; Radzimanowski, J.; Effantin, G.; Midtvedt, D.; Mangenot, S.; Weissenhorn, W.; Bassereau, P.; Bally, M. The Matrix Protein M1 from Influenza C Virus Induces Tubular Membrane Invaginations in an in Vitro Cell Membrane Model. Sci. Rep. 2017, 7, 40801. [Google Scholar] [CrossRef] [PubMed]
- Soni, S.P.; Adu-Gyamfi, E.; Yong, S.S.; Jee, C.S.; Stahelin, R.V. The Ebola Virus Matrix Protein Deeply Penetrates the Plasma Membrane: An Important Step in Viral Egress. Biophys. J. 2013, 104, 1940–1949. [Google Scholar] [CrossRef] [PubMed]
- Soni, S.P.; Stahelin, R.V. The Ebola Virus Matrix Protein VP40 Selectively Induces Vesiculation from Phosphatidylserine-Enriched Membranes. J. Biol. Chem. 2014, 289, 33590–33597. [Google Scholar] [CrossRef]
- Podkalicka, J.; Blouin, C.M. GPMVs as a Tool to Study Caveolin-Interacting Partners. In Caveolae; Blouin, C.M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; Volume 2169, pp. 81–88. ISBN 978-1-07-160731-2. [Google Scholar]
- Levental, K.R.; Levental, I. Giant Plasma Membrane Vesicles: Models for Understanding Membrane Organization. In Current Topics in Membranes; Elsevier: Amsterdam, The Netherlands, 2015; Volume 75, pp. 25–57. ISBN 978-0-12-803295-4. [Google Scholar]
- Gerstle, Z.; Desai, R.; Veatch, S.L. Giant Plasma Membrane Vesicles: An Experimental Tool for Probing the Effects of Drugs and Other Conditions on Membrane Domain Stability. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 603, pp. 129–150. ISBN 978-0-12-814574-6. [Google Scholar]
- Marcsisin, S.R.; Engen, J.R. Hydrogen Exchange Mass Spectrometry: What Is It and What Can It Tell Us? Anal. Bioanal. Chem. 2010, 397, 967–972. [Google Scholar] [CrossRef]
- Molecular Virology of Human Pathogenic Viruses; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 978-0-12-800838-6.
- Milne, J.L.S.; Borgnia, M.J.; Bartesaghi, A.; Tran, E.E.H.; Earl, L.A.; Schauder, D.M.; Lengyel, J.; Pierson, J.; Patwardhan, A.; Subramaniam, S. Cryo-Electron Microscopy—A Primer for the Non-Microscopist. FEBS J. 2013, 280, 28–45. [Google Scholar] [CrossRef]
- Howard, M.J. Protein NMR Spectroscopy. Curr. Biol. 1998, 8, R331–R333. [Google Scholar] [CrossRef]
- Huang, Y.; Vold, R.L.; Hoatson, G.L. Investigation of Multiaxis Molecular Motion by Off-Magic Angle Spinning Deuteron NMR. J. Chem. Phys. 2006, 124, 104504. [Google Scholar] [CrossRef] [PubMed]
- Magde, D.; Elson, E.; Webb, W.W. Thermodynamic Fluctuations in a Reacting System—Measurement by Fluorescence Correlation Spectroscopy. Phys. Rev. Lett. 1972, 29, 705–708. [Google Scholar] [CrossRef]
- Wawrezinieck, L.; Rigneault, H.; Marguet, D.; Lenne, P.-F. Fluorescence Correlation Spectroscopy Diffusion Laws to Probe the Submicron Cell Membrane Organization. Biophys. J. 2005, 89, 4029–4042. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, C.; Ringemann, C.; Medda, R.; Schwarzmann, G.; Sandhoff, K.; Polyakova, S.; Belov, V.N.; Hein, B.; von Middendorff, C.; Schönle, A.; et al. Direct Observation of the Nanoscale Dynamics of Membrane Lipids in a Living Cell. Nature 2009, 457, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Favard, C.; Wenger, J.; Lenne, P.-F.; Rigneault, H. FCS Diffusion Laws in Two-Phase Lipid Membranes: Determination of Domain Mean Size by Experiments and Monte Carlo Simulations. Biophys. J. 2011, 100, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Vicidomini, G.; Bianchini, P.; Diaspro, A. STED Super-Resolved Microscopy. Nat. Methods 2018, 15, 173–182. [Google Scholar] [CrossRef]
- Zhao, H.; Lappalainen, P. A Simple Guide to Biochemical Approaches for Analyzing Protein–Lipid Interactions. Mol. Biol. Cell 2012, 23, 2823–2830. [Google Scholar] [CrossRef]
- Hamard-Peron, E.; Juillard, F.; Saad, J.S.; Roy, C.; Roingeard, P.; Summers, M.F.; Darlix, J.-L.; Picart, C.; Muriaux, D. Targeting of Murine Leukemia Virus Gag to the Plasma Membrane Is Mediated by PI(4,5)P2/PS and a Polybasic Region in the Matrix. J. Virol. 2010, 84, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Nano-Inspired Biosensors for Protein Assay with Clinical Applications; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-12-815053-5.
- Digman, M.A.; Brown, C.M.; Sengupta, P.; Wiseman, P.W.; Horwitz, A.R.; Gratton, E. Measuring Fast Dynamics in Solutions and Cells with a Laser Scanning Microscope. Biophys. J. 2005, 89, 1317–1327. [Google Scholar] [CrossRef]
- Miles, A.J.; Janes, R.W.; Wallace, B.A. Tools and Methods for Circular Dichroism Spectroscopy of Proteins: A Tutorial Review. Chem. Soc. Rev. 2021, 50, 8400–8413. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brémaud, E.; Favard, C.; Muriaux, D. Deciphering the Assembly of Enveloped Viruses Using Model Lipid Membranes. Membranes 2022, 12, 441. https://doi.org/10.3390/membranes12050441
Brémaud E, Favard C, Muriaux D. Deciphering the Assembly of Enveloped Viruses Using Model Lipid Membranes. Membranes. 2022; 12(5):441. https://doi.org/10.3390/membranes12050441
Chicago/Turabian StyleBrémaud, Erwan, Cyril Favard, and Delphine Muriaux. 2022. "Deciphering the Assembly of Enveloped Viruses Using Model Lipid Membranes" Membranes 12, no. 5: 441. https://doi.org/10.3390/membranes12050441
APA StyleBrémaud, E., Favard, C., & Muriaux, D. (2022). Deciphering the Assembly of Enveloped Viruses Using Model Lipid Membranes. Membranes, 12(5), 441. https://doi.org/10.3390/membranes12050441