
Characterization of the Role of Extracellular Vesicles Released from Chicken Tracheal Cells in the Antiviral Responses against Avian Influenza Virus


Abstract
1. Introduction
2. Material and Methods
2.1. Avian Influenza Virus (AIV)
2.2. Toll-Like Receptor (TLR) Ligands
2.3. Tracheal Organ Culture (TOC)
2.4. Chicken Macrophage Cell Line
2.5. Determining Protein Content of EVs Released from TOC
2.5.1. TOC Infection with AIV and Stimulation with TLR Ligands
2.5.2. EV Isolation
2.5.3. Sample Preparation for Mass Spectrometry Analysis
2.5.4. Mass Spectrometry-Based Proteomics
2.5.5. Protein Functional Analyses
2.6. Treatment of Chicken Macrophages with EVs Released from TOC
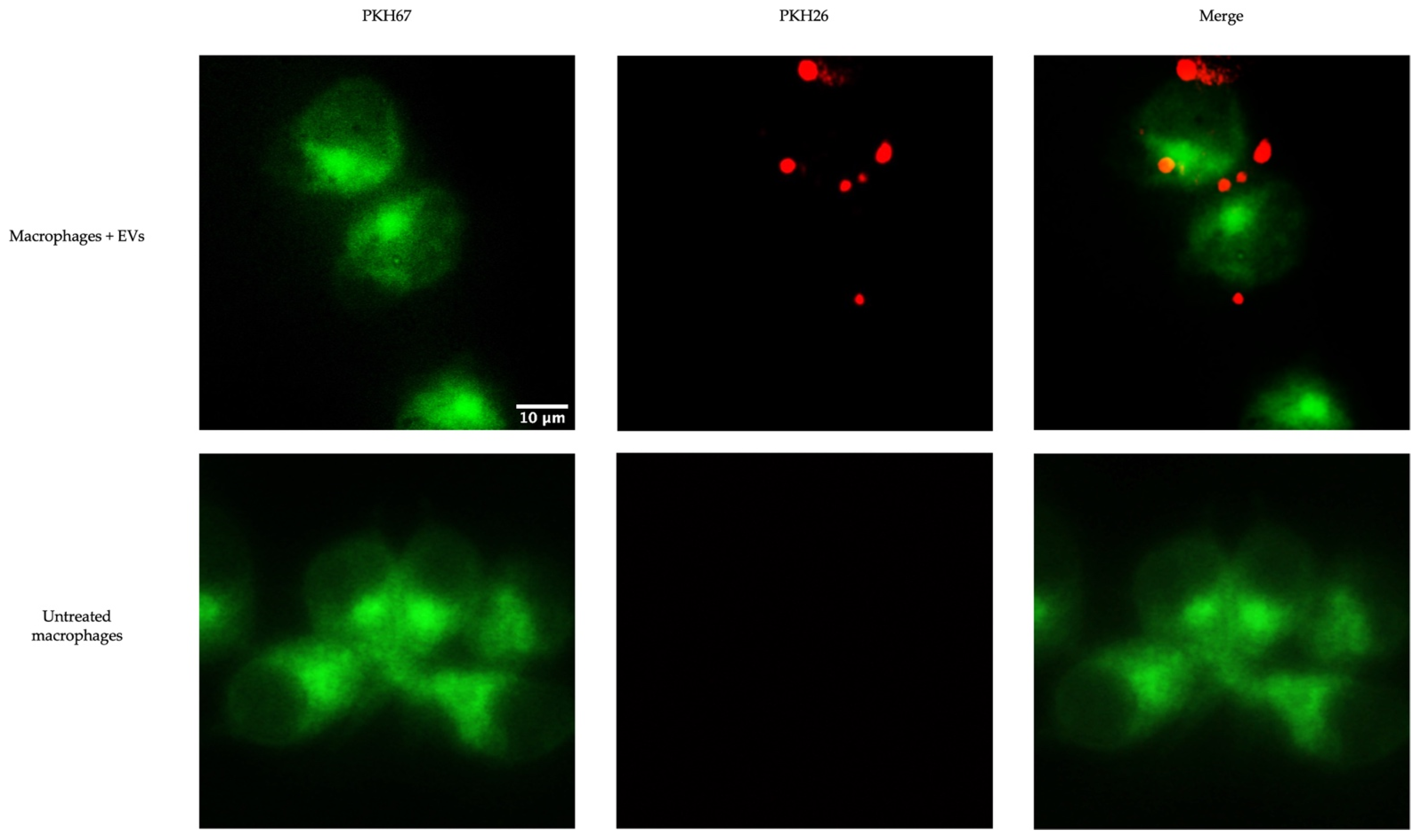
2.6.1. EV Uptake by Chicken Macrophages
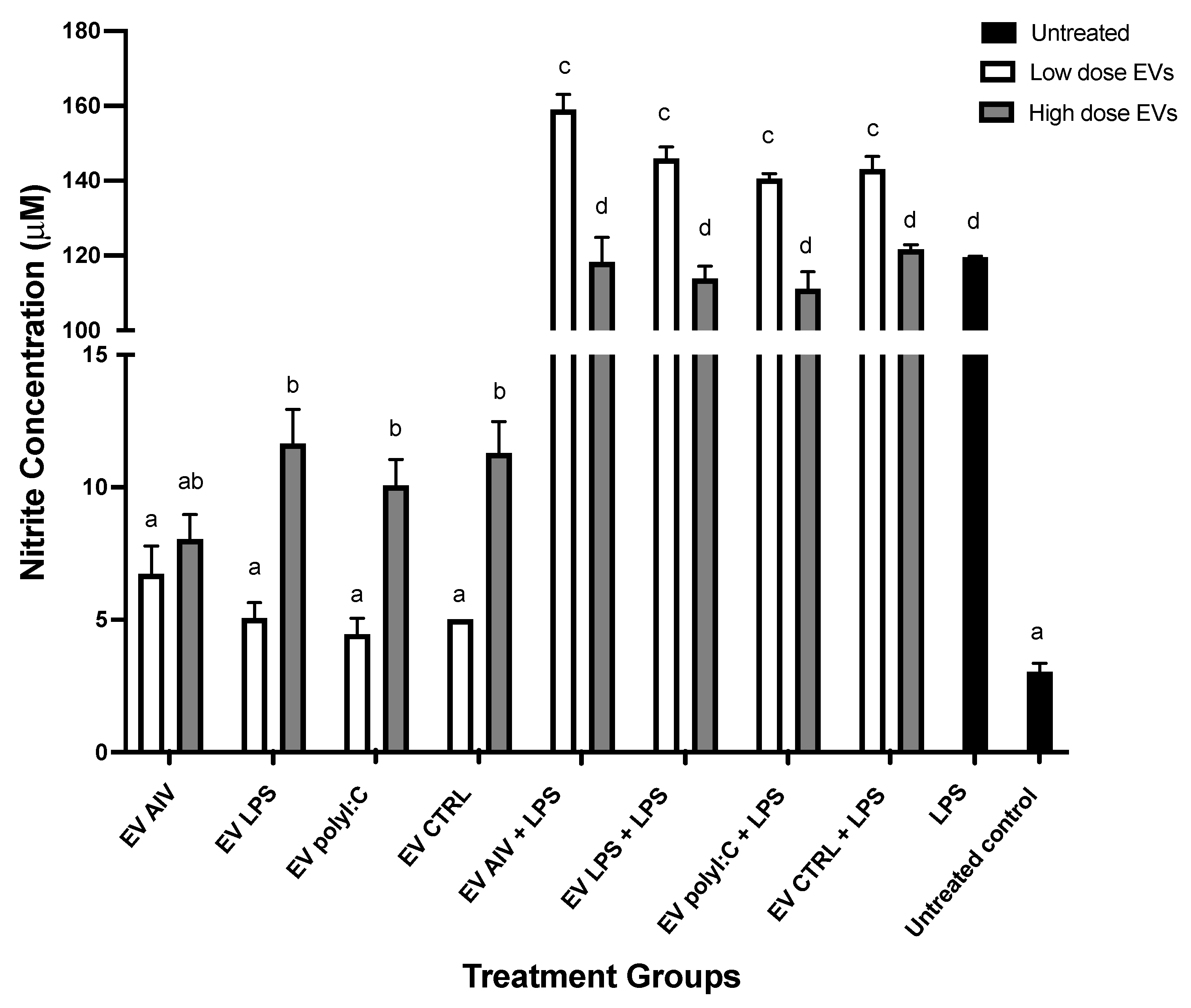
2.6.2. Nitric Oxide (NO) Production by Chicken Macrophages Treated with EVs
2.6.3. Phagocytosis by Chicken Macrophages Treated with EVs
2.6.4. Gene Expression of Chicken Macrophages Treated with EVs
2.7. Statistical Analysis
3. Results
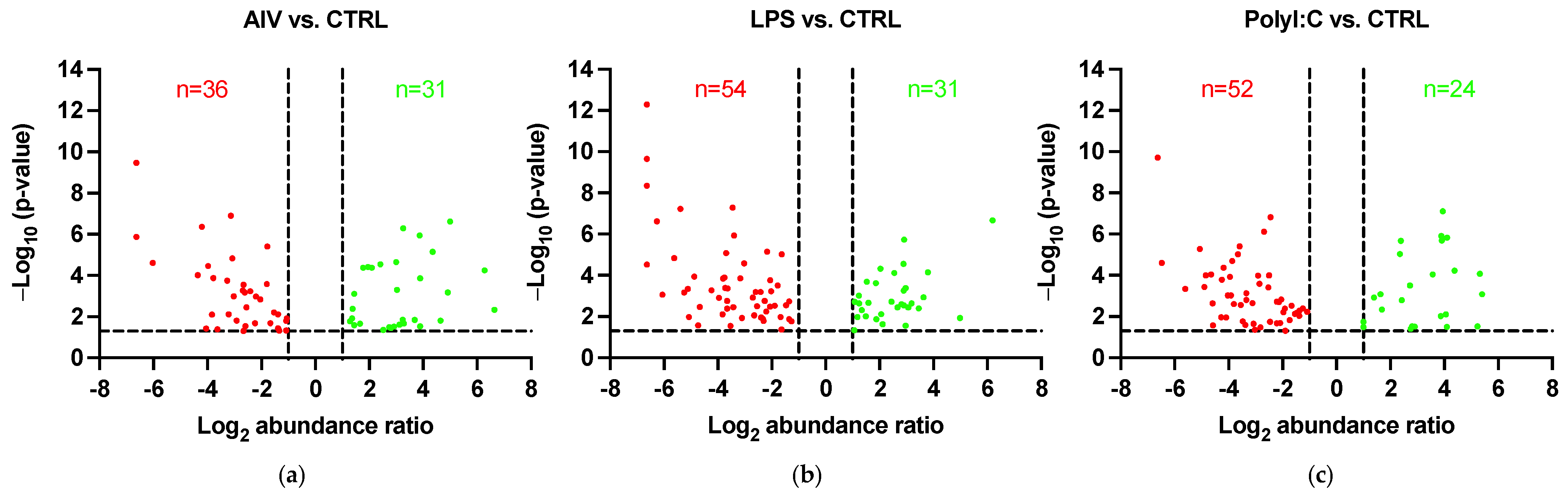
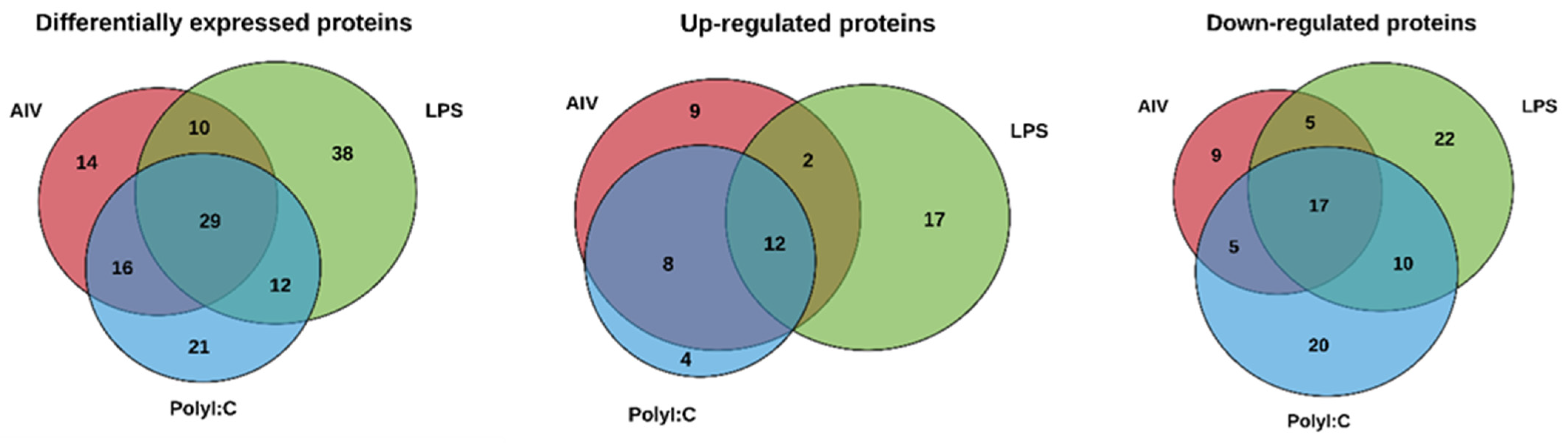
3.1. EVs Released from TOCs Treated with AIV, LPS, and PolyI:C Have Distinctive Protein Profiles
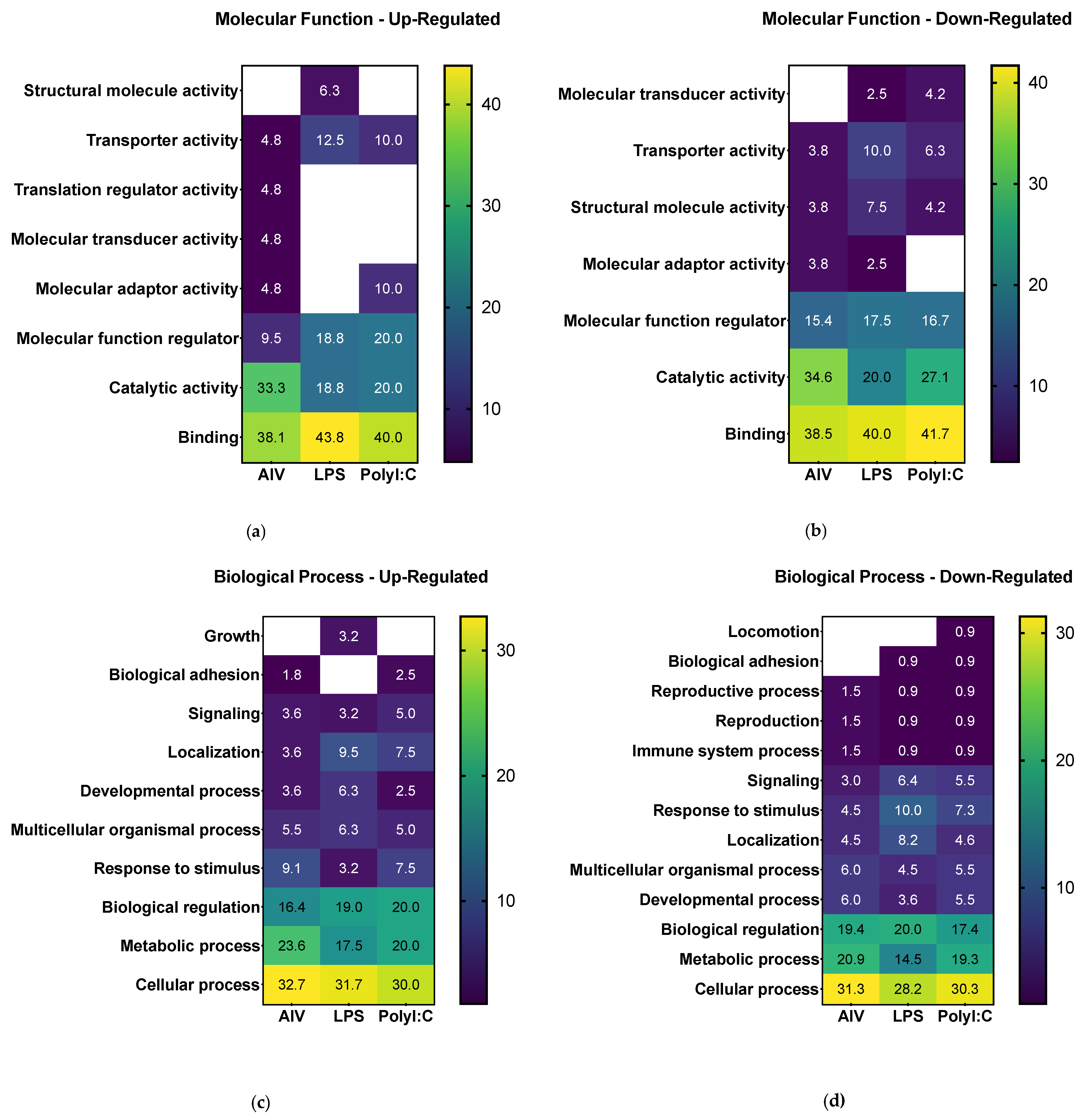
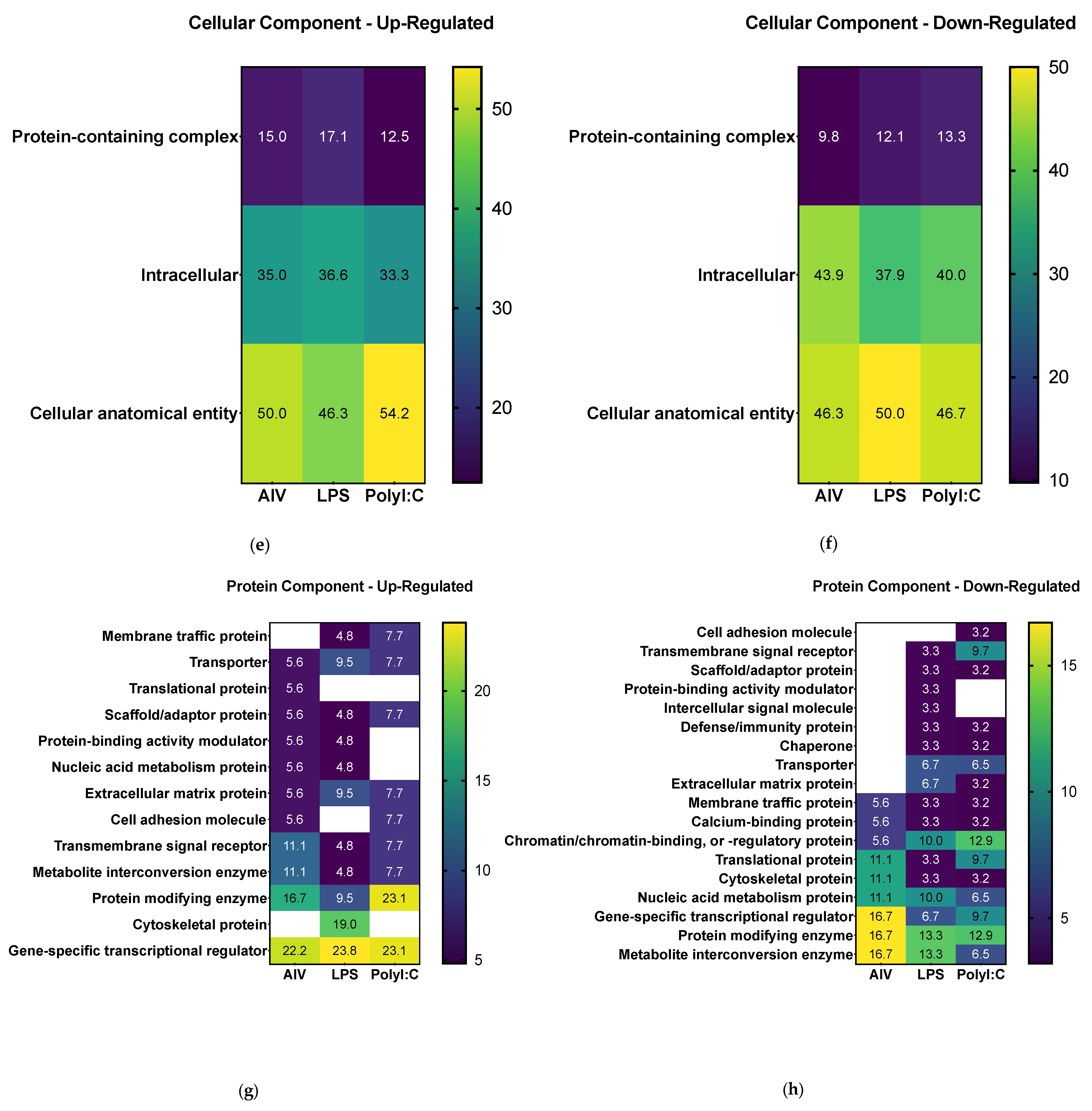
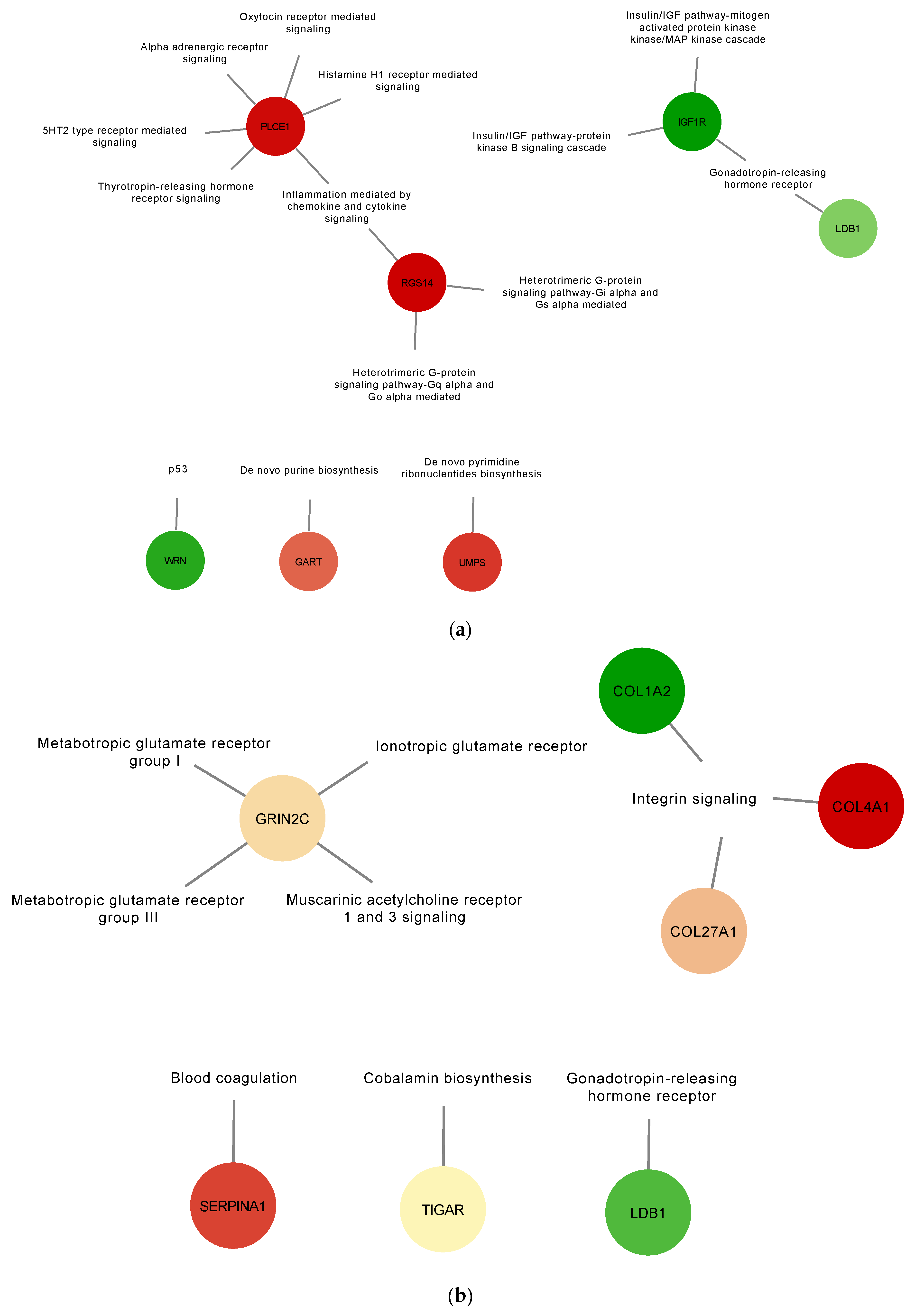
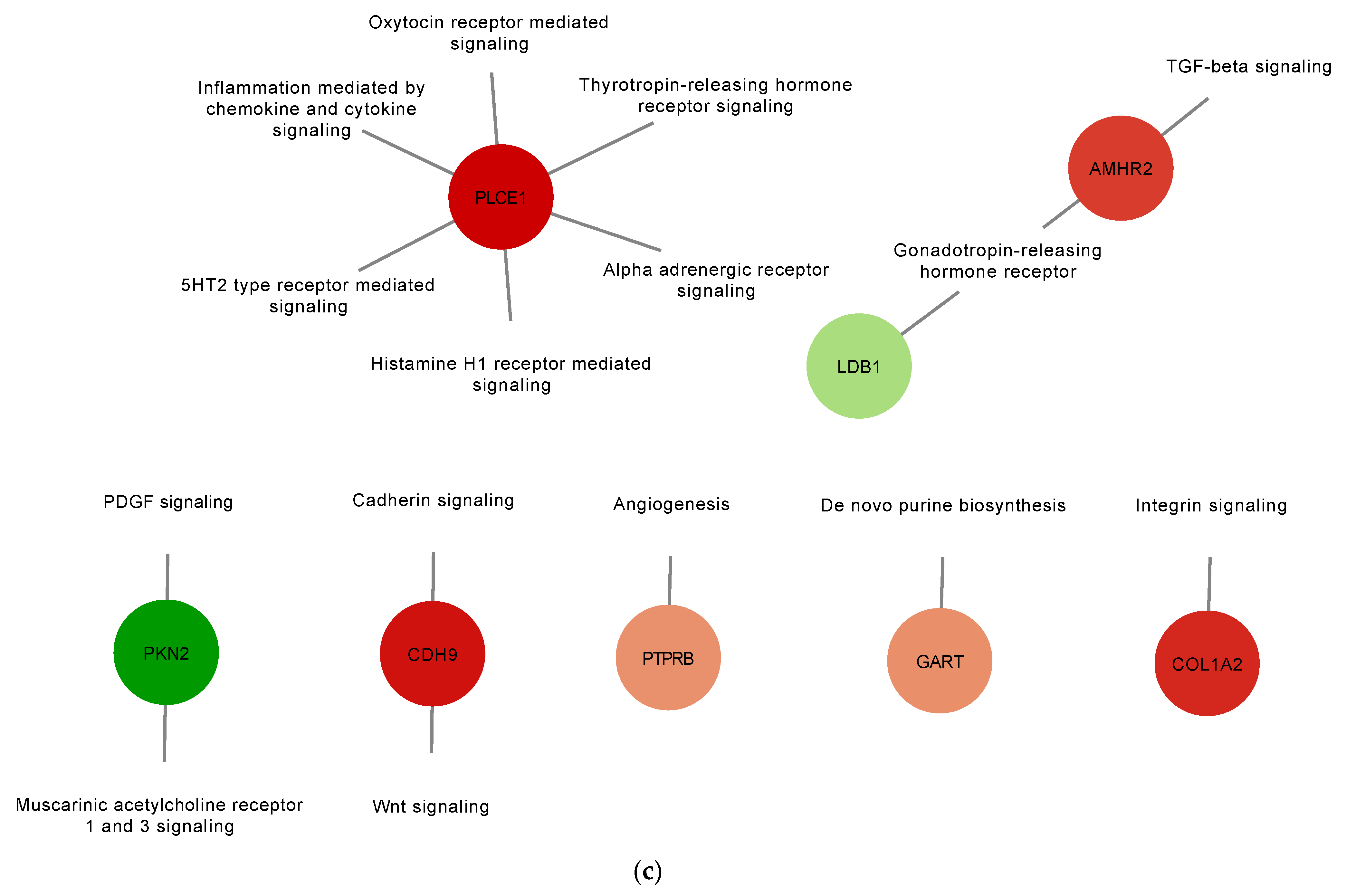
3.2. Proteins Found in EVs Released from TOCs Have Functions in Cell Signaling and Immune System Processes
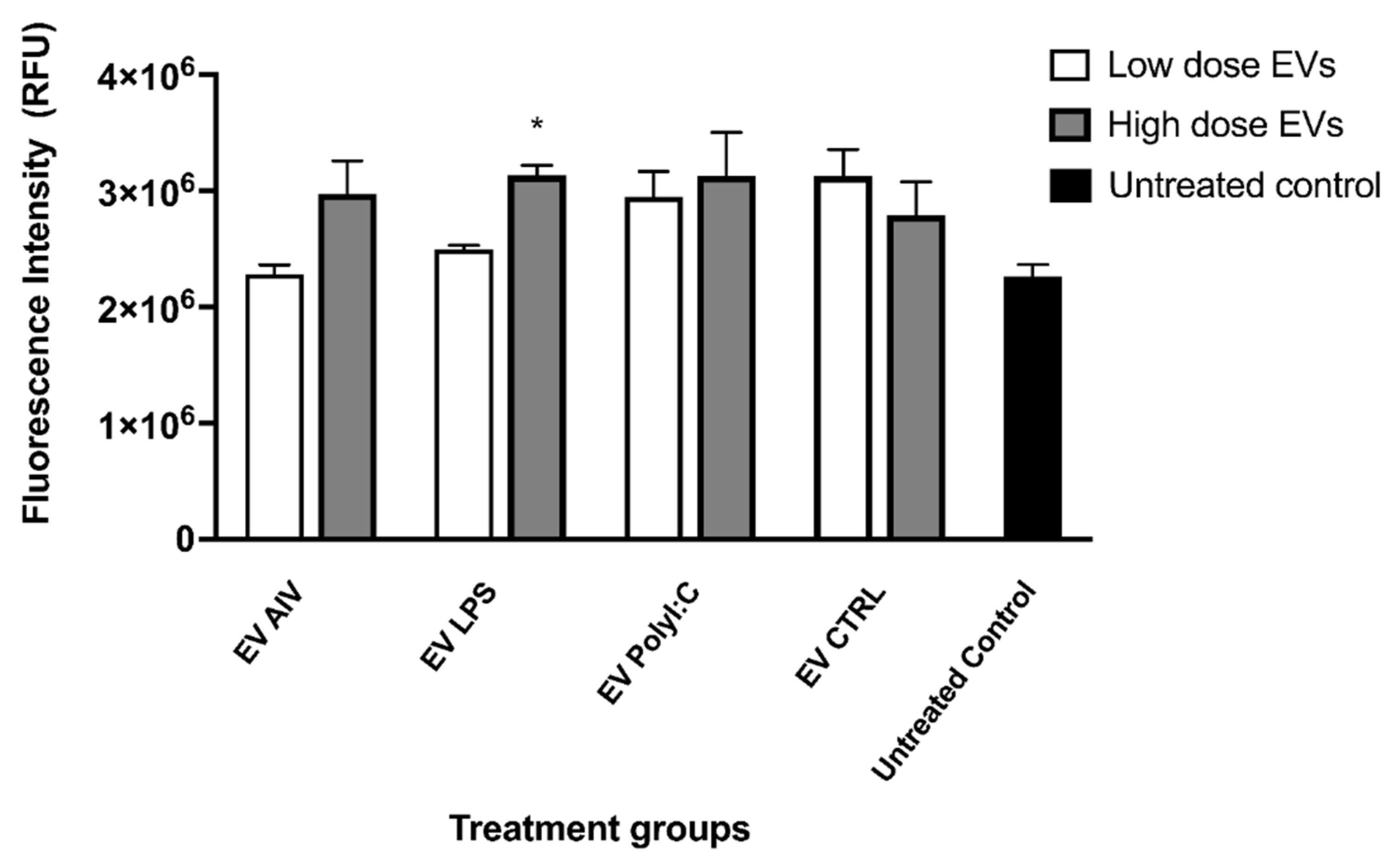
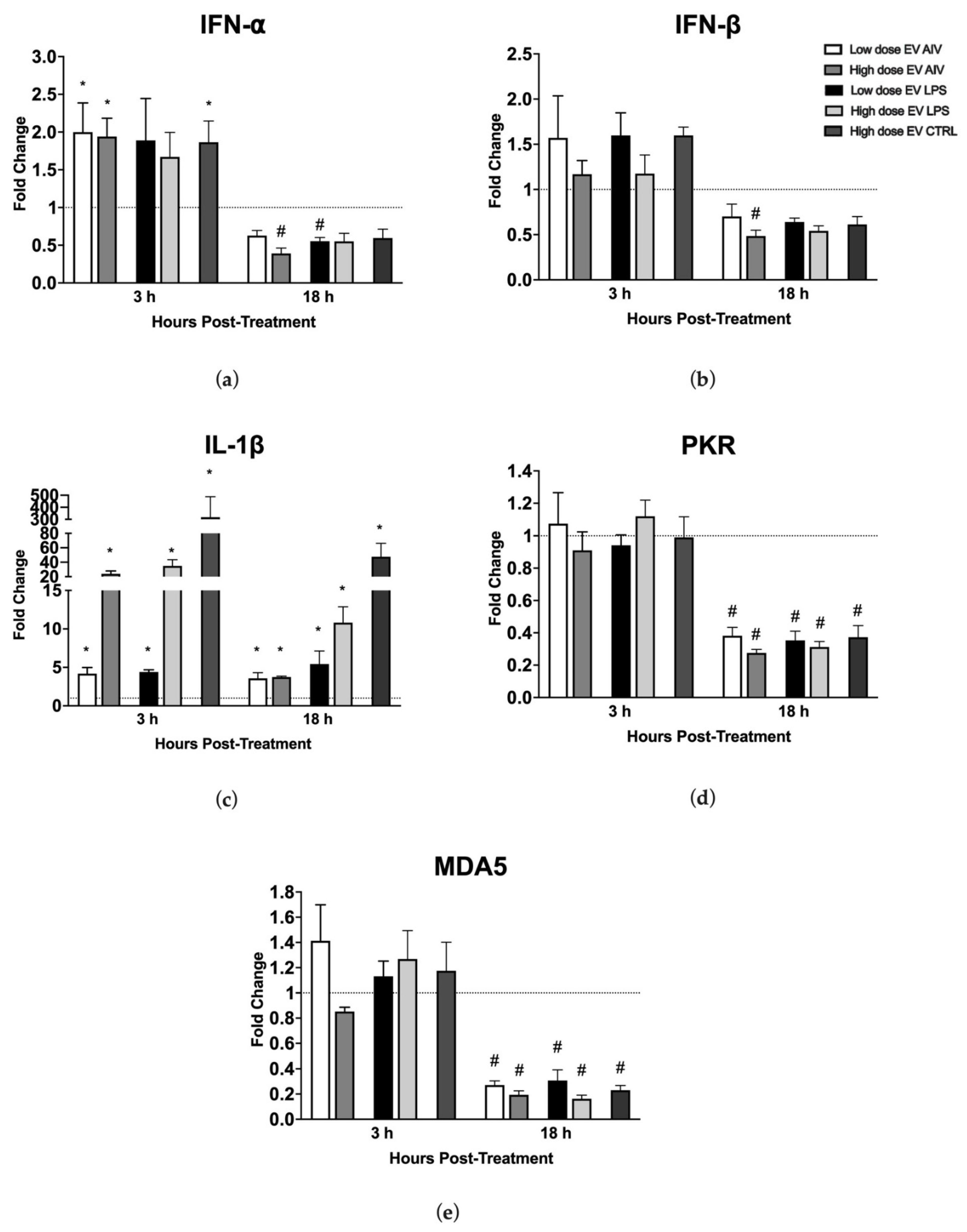
3.3. EVs Released from TOCs Impact Chicken Macrophage Functions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miura, T.A. Respiratory Epithelial Cells as Master Communicators during Viral Infections. Curr. Clin. Microbiol. Rep. 2019, 6, 10–17. [Google Scholar] [CrossRef]
- Yan, N.; Chen, Z.J. Intrinsic antiviral immunity. Nat. Immunol. 2012, 13, 214–222. [Google Scholar] [CrossRef]
- McNab, F.; Mayer-Barber, K.; Sher, A.; Wack, A.; O’Garra, A. Type I interferons in infectious disease. Nat. Rev. Immunol. 2015, 15, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Rivera, A.; Siracusa, M.C.; Yap, G.S.; Gause, W.C. Innate cell communication kick-starts pathogen-specific immunity. Nat. Immunol. 2016, 17, 356–363. [Google Scholar] [CrossRef]
- Qureshi, M.A.; Heggen, C.L.; Hussain, I. Avian macrophage: Effector functions in health and disease. Dev. Comp. Immunol. 2000, 24, 103–119. [Google Scholar] [CrossRef]
- Dreux, M.; Garaigorta, U.; Boyd, B.; Décembre, E.; Chung, J.; Whitten-Bauer, C.; Wieland, S.; Chisari, F.V. Short-Range Exosomal Transfer of Viral RNA from Infected Cells to Plasmacytoid Dendritic Cells Triggers Innate Immunity. Cell Host Microbe 2012, 12, 558–570. [Google Scholar] [CrossRef]
- Brennan, K. A comparison of methods for the isolation and separation of extracellular vesicles from protein and lipid particles in human serum. Sci. Rep. 2020, 10, 1039. [Google Scholar]
- Yáñez-Mó, M.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; Colás, E.; Cordeiro-da, A.; Fais, S.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Vizio, D.D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal experimental requirements for definition of extracellular vesicles and their functions: A position statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef] [PubMed]
- Toribio, V.; Morales, S.; López-Martín, S.; Cardeñes, B.; Cabañas, C.; Yáñez-Mó, M. Development of a quantitative method to measure EV uptake. Sci. Rep. 2019, 9, 10522. [Google Scholar] [CrossRef] [PubMed]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Quaglia, M.; Dellepiane, S.; Guglielmetti, G.; Merlotti, G.; Castellano, G.; Cantaluppi, V. Extracellular Vesicles as Mediators of Cellular Crosstalk Between Immune System and Kidney Graft. Front. Immunol. 2020, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Felekkis, K.; Touvana, E.; Stefanou, C.; Deltas, C. MicroRNAs: A newly described class of encoded molecules that play a role in health and disease. Hippokratia 2010, 14, 236–240. [Google Scholar]
- Chahar, H.S.; Corsello, T.; Kudlicki, A.S.; Komaravelli, N.; Casola, A. Respiratory Syncytial Virus Infection Changes Cargo Composition of Exosome Released from Airway Epithelial Cells. Sci. Rep. 2018, 8, 387. [Google Scholar] [CrossRef]
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Dillon, S.T.; Reeves, R.K.; Manickam, C.; Morgello, S.; Kirk, G.D.; Mehta, S.H.; Gabuzda, D. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci. Rep. 2018, 8, 7227. [Google Scholar] [CrossRef]
- Bello-Morales, R. Extracellular Vesicles in Viral Spread and Antiviral Response. Viruses 2020, 12, 623. [Google Scholar] [CrossRef]
- Petrik, J. Immunomodulatory effects of exosomes produced by virus-infected cells. Transfus. Apher. Sci. 2016, 55, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Qiao, Y.; Li, X.; Chen, J.; Ding, J.; Bai, L.; Shen, F.; Shi, B.; Liu, J.; Peng, L.; et al. Exosomes Exploit the Virus Entry Machinery and Pathway To Transmit Alpha Interferon-Induced Antiviral Activity. J. Virol. 2018, 92, e01578-18. [Google Scholar] [CrossRef]
- Hong, Y.; Truong, A.D.; Lee, J.; Vu, T.H.; Lee, S.; Song, K.D.; Lillehoj, H.S.; Hong, Y.H. Exosomal miRNA profiling from H5N1 avian influenza virus-infected chickens. Vet. Res. 2021, 52, 36. [Google Scholar] [CrossRef]
- del Cacho, E.; Gallego, M.; Lee, S.H.; Lillehoj, H.S.; Quilez, J.; Lillehoj, E.P.; Sánchez-Acedo, C. Induction of Protective Immunity against Eimeria tenella, Eimeria maxima, and Eimeria acervulina Infections Using Dendritic Cell-Derived Exosomes. Infect. Immun. 2012, 80, 1909–1916. [Google Scholar] [CrossRef]
- del Cacho, E.; Gallego, M.; Lillehoj, H.S.; Quilez, J.; Lillehoj, E.P.; Sánchez-Acedo, C. Induction of protective immunity against experimental Eimeria tenella infection using serum exosomes. Vet. Parasitol. 2016, 224, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fu, Y.; Zou, M.; Sun, Y.; Yin, X.; Niu, L.; Peng, X. Analysis of deep sequencing Exosome-microRNA expression profile from Chicken Type Ⅱ Pneumocytes derived reveals potential role of gga-miRNA-451 in inflammation. J. Cell. Mol. Med. 2020, 24, 6178–6190. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, G.; Wang, Z.; Zhang, H.; Zhang, L.; Cheng, Z. Chicken biliary exosomes enhance CD4+T proliferation and inhibit ALV-J replication in liver. Biochem. Cell Biol. 2014, 92, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Neerukonda, S.N.; Tavlarides-Hontz, P.; McCarthy, F.; Pendarvis, K.; Parcells, M.S. Comparison of the Transcriptomes and Proteomes of Serum Exosomes from Marek’s Disease Virus-Vaccinated and Protected and Lymphoma-Bearing Chickens. Genes 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhuang, P.; Cheng, Z.; Yang, J.; Bi, J.; Wang, G. Avian leukosis virus subgroup J and reticuloendotheliosis virus coinfection induced TRIM62 regulation of the actin cytoskeleton. J. Vet. Sci. 2020, 21, e49. [Google Scholar] [CrossRef]
- Ye, F.; Wang, Y.; He, Q.; Cui, C.; Yu, H.; Lu, Y.; Zhu, S.; Xu, H.; Zhao, X.; Yin, H.; et al. Exosomes transmit viral genetic information and immune signals may cause immunosuppression and immune tolerance in ALV-J infected hd11 cells. Int. J. Biol. Sci. 2020, 16, 904–920. [Google Scholar] [CrossRef]
- Hong, Y.; Lee, J.; Vu, T.H.; Lee, S.; Lillehoj, H.S.; Hong, Y.H. Exosomes of lipopolysaccharide-stimulated chicken macrophages modulate immune response through the MyD88/NF-κB signaling pathway. Dev. Comp. Immunol. 2021, 115, 103908. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Lee, J.; Vu, T.H.; Lee, S.; Lillehoj, H.S.; Hong, Y.H. Immunomodulatory effects of poly(I:C)-stimulated exosomes derived from chicken macrophages. Poult. Sci. 2021, 100, 101247. [Google Scholar] [CrossRef]
- Barjesteh, N.; Behboudi, S.; Brisbin, J.T.; Villanueva, A.I.; Nagy, É.; Sharif, S. TLR ligands induce antiviral responses in chicken macrophages. PLoS ONE 2014, 9, e105713. [Google Scholar] [CrossRef]
- Barjesteh, N.; Shojadoost, B.; Brisbin, J.T.; Emam, M.; Hodgins, D.C.; Nagy, É.; Sharif, S. Reduction of avian influenza virus shedding by administration of Toll-like receptor ligands to chickens. Vaccine 2015, 33, 4843–4849. [Google Scholar] [CrossRef]
- Barjesteh, N.; Alkie, T.N.; Hodgins, D.C.; Nagy, É.; Sharif, S. Local innate responses to TLR ligands in the chicken trachea. Viruses 2016, 8, 207. [Google Scholar] [CrossRef]
- Barjesteh, N.; Taha-Abdelaziz, K.; Kulkarni, R.R.; Sharif, S. Innate antiviral responses are induced by TLR3 and TLR4 ligands in chicken tracheal epithelial cells: Communication between epithelial cells and macrophages. Virology 2019, 534, 132–142. [Google Scholar] [CrossRef]
- O’Dowd, K.; Emam, M.; El Khili, M.R.; Emad, A.; Ibeagha-Awemu, E.M.; Gagnon, C.A.; Barjesteh, N. Distinct miRNA Profile of Cellular and Extracellular Vesicles Released from Chicken Tracheal Cells Following Avian Influenza Virus Infection. Vaccines 2020, 8, 438. [Google Scholar] [CrossRef] [PubMed]
- Szretter, K.J.; Balish, A.L.; Katz, J.M. Influenza: Propagation, quantification, and storage. Curr. Protoc. Microbiol. 2006, 3, 15G-1. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.; Cox, N.; Stohr, K. WHO Manual on Animal Influenza Diagnosis and Surveillance: World Health Organization, Department of Communicable Disease Surveillance and Response. WHO/CDS/CDR/2002.5. World Health Organ. 2002, 1, 48–54. [Google Scholar]
- Qureshi, M.A.; Miller, L.; Lillehoj, H.S.; Ficken, M.D. Establishment and characterization of a chicken mononuclear cell line. Vet. Immunol. Immunopathol. 1990, 26, 237–250. [Google Scholar] [CrossRef]
- Hossain, M.J.; Mori, I.; Dong, L.; Liu, B.; Kimura, Y. Fetal calf serum inhibits virus genome expression in Madin-Darby canine kidney cells persistently infected with influenza A virus. Med. Microbiol. Immunol. 2008, 197, 21–27. [Google Scholar] [CrossRef] [PubMed]
- The, M.; MacCoss, M.J.; Noble, W.S.; Käll, L. Fast and Accurate Protein False Discovery Rates on Large-Scale Proteomics Data Sets with Percolator 3.0. J. Am. Soc. Mass Spectrom. 2016, 27, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Orsburn, B.C. Proteome discoverer-a community enhanced data processing suite for protein informatics. Proteomes 2021, 9, 15. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Lässer, C.; Seyed Alikhani, V.; Ekström, K.; Eldh, M.; Torregrosa Paredes, P.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human saliva, plasma and breast milk exosomes contain RNA: Uptake by macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Preibisch, S.; Saalfeld, S.; Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics 2009, 25, 1463–1465. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- St Paul, M.; Mallick, A.I.; Haq, K.; Orouji, S.; Abdul-Careem, M.F.; Sharif, S. In vivo administration of ligands for chicken toll-like receptors 4 and 21 induces the expression of immune system genes in the spleen. Vet. Immunol. Immunopathol. 2011, 144, 228–237. [Google Scholar] [CrossRef]
- Villanueva, A.I.; Kulkarni, R.R.; Sharif, S. Synthetic double-stranded RNA oligonucleotides are immunostimulatory for chicken spleen cells. Dev. Comp. Immunol. 2011, 35, 28–34. [Google Scholar] [CrossRef]
- Raposo, G.; Stahl, P.D. Extracellular vesicles: A new communication paradigm? Nat. Rev. Mol. Cell Biol. 2019, 20, 509–510. [Google Scholar] [CrossRef] [PubMed]
- Margolis, L.; Sadovsky, Y. The biology of extracellular vesicles: The known unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef] [PubMed]
- White, J.R.; Dauros-Singorenko, P.; Hong, J.; Vanholsbeeck, F.; Phillips, A.; Swift, S. The complex, bidirectional role of extracellular vesicles in infection. Biochem. Soc. Trans. 2021, 49, 881–891. [Google Scholar] [CrossRef]
- Gupta, R.; Radicioni, G.; Abdelwahab, S.; Dang, H.; Carpenter, J.; Chua, M.; Mieczkowski, P.A.; Sheridan, J.T.; Randell, S.H.; Kesimer, M. Intercellular communication between airway epithelial cells is mediated by exosome-like vesicles. Am. J. Respir. Cell Mol. Biol. 2019, 60, 209–220. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, X.; Yao, J.; Guo, H.; Yin, L.; Leung, W.; Xu, C. Role of Extracellular Vesicles in Influenza Virus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Dorshorst, B.; Harun-Or-Rashid, M.; Bagherpoor, A.J.; Rubin, C.J.; Ashwell, C.; Gourichon, D.; Tixier-Boichard, M.; Hallböök, F.; Andersson, L. A Genomic Duplication is Associated with Ectopic Eomesodermin Expression in the Embryonic Chicken Comb and Two Duplex-comb Phenotypes. PLoS Genet. 2015, 11, e1004947. [Google Scholar] [CrossRef]
- Li, J.; He, Y.; Hao, J.; Ni, L.; Dong, C. High Levels of Eomes Promote Exhaustion of Anti-tumor CD8+ T Cells. Front. Immunol. 2018, 9, 2981. [Google Scholar] [CrossRef]
- Pearce, E.L.; Mullen, A.C.; Martins, G.A.; Krawczyk, C.M.; Hutchins, A.S.; Zediak, V.P.; Banica, M.; DiCioccio, C.B.; Gross, D.A.; Mao, C.A.; et al. Control of Effector CD8+ T Cell Function by the Transcription Factor Eomesodermin. Science 2003, 302, 1041–1043. [Google Scholar] [CrossRef]
- Kurachi, M. CD8 + T cell exhaustion. Semin. Immunopathol. 2019, 41, 327–337. [Google Scholar] [CrossRef]
- Cui, X.; Xin, H.; Peng, H.; Chen, Y. Comprehensive bioinformatics analysis of the mRNA profile of PLCE1 knockdown in esophageal squamous cell carcinoma. Mol. Med. Rep. 2017, 16, 5871–5880. [Google Scholar] [CrossRef][Green Version]
- Shimaoka, M.; Kawamoto, E.; Gaowa, A.; Okamoto, T.; Park, E.J. Connexins and integrins in exosomes. Cancers 2019, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Albacete-Albacete, L. Extracellular Vesicles: An Emerging Mechanism Governing the Secretion and Biological Roles of Tenascin-C. Front. Immunol. 2021, 12, 671485. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef]
- Koliha, N. Analysis of the microRNA profile and origin of exosomes in plasma of melanoma patients and healthy individuals. Friedrich-Alexander-Univ. Erlangen-Nürnb. 2016, 115, 6779–6789. [Google Scholar]
- Li, Y.H.; Yan, Z.Q.; Brauner, A.; Tullus, K. Activation of macrophage nuclear factor-κB and induct ion of inducible nitric oxide synthase by LPS. Respir. Res. 2002, 3, 23. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation David. Nat. Rev. Immunol. 2009, 8, 958–969. [Google Scholar] [CrossRef]
- Kopincová, J.; Púzserová, A.; Bernátová, I. Biochemical aspects of nitric oxide synthase feedback regulation by nitric oxide. Interdiscip. Toxicol. 2011, 4, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, D.; Rubbenstroth, D.; Martinez-Sobrido, L.; Munir, M. Avian interferons and their antiviral effectors. Front. Immunol. 2017, 8, 49. [Google Scholar] [CrossRef]
- Kwon, S.; Newcomb, R.L.; George, S.C. Mechanisms of synergistic cytokine-induced nitric oxide production in human alveolar epithelial cells. Nitric Oxide 2001, 5, 534–546. [Google Scholar] [CrossRef]
- Dauber, B.; Wolff, T. Activation of the antiviral kinase PKR and viral countermeasures. Viruses 2009, 1, 523–544. [Google Scholar] [CrossRef]
- Dias Junior, A.G.; Sampaio, N.G.; Rehwinkel, J. A Balancing Act: MDA5 in Antiviral Immunity and Autoinflammation. Trends Microbiol. 2019, 27, 75–85. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence | Annealing Temperature (°C) | Reference |
|---|---|---|---|
| β-actin | F: 5′-CAACACAGTGCTGTCTGGTGGTA-3′ | 60 | [49] |
| R: 5′-ATCGTACTCCTGCTTGCTGATCC-3′ | |||
| IFN-α | F: 5′-ATCCTGCTGCTCACGCTCCTTCT-3′ | 64 | [49] |
| R: 5′-GGTGTTGCTGGTGTCCAGGATG-3′ | |||
| IFN-β | F: 5′-GCCTCCAGCTCCTTCAGAATAC G-3′ | 64 | [50] |
| R: 5′-CTGGATCTGGTTGAGGAGGCTGT-3′ | |||
| IL-1β | F: 5′-GTGAGGCTCAACATTGCGCTGTA-3′ | 64 | [49] |
| R: 5′-TGTCCAGGCGGTAGAAGATGAAG-3′ | |||
| MDA5 | F: 5′-TGGTACAGGCGTTGGTAAGAG-3′ | 60 | [30] |
| R: 5′-GAGCACATCCGCAGGTAGAG-3′ | |||
| PKR | F: 5′-GCAAAACCAGCACTGAATGGG-3′ | 60 | [30] |
| R: 5′-CGTAAATGCTGTTCCACTAACGG-3′ |
| Treatment Group | Accession | Protein Name | Gene Symbol | Abundance Ratio | Log2 Abundance Ratio | p-Value |
|---|---|---|---|---|---|---|
| AIV (31 proteins) | A0A1D5PJC0 | Uncharacterized protein | - | 100.00 | 6.64 | 4.57 × 10−3 |
| A0A3Q2TWB1 | Phosphoinositide 5-phosphatase | INPP5B | 77.51 | 6.28 | 5.62 × 10−5 | |
| A0A1D5P5K6 | PHD-type domain-containing protein | TCF20 | 32.00 | 5.00 | 2.39 × 10−7 | |
| A0A3Q2U363 | Uncharacterized protein | ADGRA1 | 30.12 | 4.91 | 6.68 × 10−4 | |
| E1BU50 | Uncharacterized protein | LOC419409 | 24.96 | 4.64 | 1.52 × 10−2 | |
| A0A1D5P4T1 | Uncharacterized protein | SYTL2 | 20.43 | 4.35 | 7.06 × 10−6 | |
| A0A3Q2UIP4 | AT-rich interactive domain-containing protein 5B | ARID5B | 14.84 | 3.89 | 1.36 × 10−4 | |
| E1BQH9 | Uncharacterized protein | UGGT2 | 14.84 | 3.89 | 2.92 × 10−2 | |
| F1P186 | Aa_trans domain-containing protein | SLC38A7 | 14.67 | 3.87 | 1.15 × 10−6 | |
| A0A1D5P8Q3 | Uncharacterized protein | PAPLN | 12.78 | 3.68 | 1.42 × 10−2 | |
| A0A1D5PVH7 | Tyrosine-protein kinase receptor | IGF1R | 9.73 | 3.28 | 2.14 × 10−2 | |
| F1NZ61 | ZnMc domain-containing protein | MMP27 | 9.59 | 3.26 | 5.12 × 10−7 | |
| A0A1L1RVC2 | hSH3 domain-containing protein | C8H1orf168 | 9.47 | 3.24 | 1.37 × 10−2 | |
| E1BSS2 | Ubiquitin specific peptidase 53 | USP53 | 9.46 | 3.24 | 2.00 × 10−2 | |
| A0A1L1RJ96 | FSA_C domain-containing protein | KIAA1109 | 8.67 | 3.12 | 2.45 × 10−2 | |
| A0A3Q2UBZ4 | Uncharacterized protein | MSLNL | 8.18 | 3.03 | 4.95 × 10−4 | |
| F1NG87 | TPR_REGION domain-containing protein | TTC28 | 8.01 | 3.00 | 2.24 × 10−5 | |
| F1NX10 | Uncharacterized protein | FRAS1 | 7.57 | 2.92 | 2.99 × 10−2 | |
| F1NCE1 | Eukaryotic translation initiation factor 3 subunit D | EIF3D | 6.95 | 2.80 | 3.51 × 10−2 | |
| A0A1L1RQF9 | DNA helicase | WRN | 6.67 | 2.74 | 3.19 × 10−2 | |
| A0A1L1RIR0 | FHA domain-containing protein | MCRS1 | 5.72 | 2.52 | 4.38 × 10−2 | |
| A0A1D5PHD3 | C2H2-type domain-containing protein | ZNF318 | 5.30 | 2.41 | 2.86 × 10−5 | |
| Q2PC93 | SCO-spondin | SSPO | 4.26 | 2.09 | 4.20 × 10−5 | |
| A0A3Q3A731 | Uncharacterized protein | - | 3.86 | 1.95 | 3.86 × 10−5 | |
| E1C1W0 | Uncharacterized protein | TRERF1 | 3.42 | 1.77 | 4.11 × 10−5 | |
| A0A3Q2UAW2 | Rho-GAP domain-containing protein | SYDE1 | 3.16 | 1.66 | 2.17 × 10−2 | |
| O42252 | LIM domain-binding protein 1 | LDB1 | 2.71 | 1.44 | 7.69 × 10−4 | |
| E1BQX8 | GRIP domain-containing protein | GOLGA1 | 2.71 | 1.44 | 2.59 × 10−2 | |
| A0A3Q2U7G6 | AF-4_C domain-containing protein | AFF1 | 2.61 | 1.38 | 4.13 × 10−3 | |
| R4GGB8 | Uncharacterized protein | GAS2L3 | 2.57 | 1.36 | 1.22 × 10−2 | |
| A0A1D5PIM5 | CARMIL_C domain-containing protein | CARMIL3 | 2.42 | 1.28 | 1.66 × 10−2 | |
| LPS (31 proteins) | A0A1D5P5K6 | PHD-type domain-containing protein | TCF20 | 73.16 | 6.19 | 2.15 × 10−7 |
| E1C4T7 | Uncharacterized protein | SETX | 31.65 | 4.98 | 1.18 × 10−2 | |
| A0A1D5P4T1 | Uncharacterized protein | SYTL2 | 13.80 | 3.79 | 7.10 × 10−5 | |
| A0A3Q2UHE4 | SH3 domain-containing protein | CTTN | 12.36 | 3.63 | 1.15 × 10−3 | |
| A0A3Q2U363 | Uncharacterized protein | ADGRA1 | 10.95 | 3.45 | 4.03 × 10−3 | |
| A0A1D5P268 | Uncharacterized protein | SHROOM4 | 9.10 | 3.19 | 2.28 × 10−3 | |
| A0A1D5PFS7 | SCA7 domain-containing protein | ATXN7 | 8.36 | 3.06 | 3.63 × 10−3 | |
| F1NF87 | Exocyst complex component 5 | EXOC5 | 7.78 | 2.96 | 4.17 × 10−4 | |
| A0A3Q2U9L9 | Uncharacterized protein | TRAPPC8 | 7.77 | 2.96 | 2.80 × 10−2 | |
| F1NZ61 | ZnMc domain-containing protein | MMP27 | 7.54 | 2.91 | 1.84 × 10−6 | |
| A0A3Q2UIP4 | AT-rich interactive domain-containing protein 5B | ARID5B | 7.49 | 2.90 | 2.93 × 10−3 | |
| A0A1D5NXL6 | Uncharacterized protein | PHF3 | 7.39 | 2.89 | 5.47 × 10−4 | |
| F1P186 | Aa_trans domain-containing protein | SLC38A7 | 7.35 | 2.88 | 2.75 × 10−5 | |
| A0A5H1ZRJ7 | Collagen alpha-2(I) chain | COL1A2 | 6.99 | 2.81 | 2.57 × 10−3 | |
| A0A1D5P7D0 | Uncharacterized protein | NUMA1 | 6.31 | 2.66 | 3.58 × 10−3 | |
| F1NG87 | TPR_REGION domain-containing protein | TTC28 | 5.84 | 2.55 | 7.78 × 10−5 | |
| A0A1D5P111 | Uncharacterized protein | KCNK5 | 5.43 | 2.44 | 1.89 × 10−3 | |
| A0A3Q2UAW2 | Rho-GAP domain-containing protein | SYDE1 | 4.32 | 2.11 | 2.35 × 10−2 | |
| A0A3Q2UPH9 | Dystrophin | DMD | 4.17 | 2.06 | 7.72 × 10−3 | |
| Q2PC93 | SCO-spondin | SSPO | 4.09 | 2.03 | 4.79 × 10−5 | |
| A0A3Q2U741 | UDENN domain-containing protein | ST5 | 3.67 | 1.88 | 1.35 × 10−2 | |
| A0A1D5PHD3 | C2H2-type domain-containing protein | ZNF318 | 3.63 | 1.86 | 2.38 × 10−4 | |
| A0A1L1RZA6 | Uncharacterized protein | LOC420370; ZNF746 | 3.00 | 1.59 | 2.13 × 10−3 | |
| E1C1W0 | Uncharacterized protein | TRERF1 | 2.86 | 1.52 | 2.03 × 10−4 | |
| A0A3Q2UAU4 | Uncharacterized protein | TMOD4 | 2.80 | 1.49 | 9.82 × 10−3 | |
| O42252 | LIM domain-binding protein 1 | LDB1 | 2.54 | 1.34 | 4.79 × 10−3 | |
| A0A3Q3A731 | Uncharacterized protein | - | 2.37 | 1.24 | 2.27 × 10−3 | |
| A0A3Q2TYZ1 | Uncharacterized protein | MPDZ | 2.34 | 1.23 | 9.71 × 10−4 | |
| E1C371 | Uncharacterized protein | LATS1 | 2.27 | 1.18 | 1.06 × 10−2 | |
| F1NCA2 | Glycerol-3-phosphate dehydrogenase | GPD2 | 2.12 | 1.08 | 1.93 × 10−3 | |
| A0A3Q2U7G6 | AF-4_C domain-containing protein | AFF1 | 2.09 | 1.06 | 4.64 × 10−2 | |
| PolyI:C (24 proteins) | A0A3Q2U363 | Uncharacterized protein | ADGRA1 | 42.37 | 5.40 | 8.29 × 10−4 |
| R4GGB8 | Uncharacterized protein | GAS2L3 | 39.94 | 5.32 | 8.45 × 10−5 | |
| A0A1D5PJC0 | Uncharacterized protein | - | 37.61 | 5.23 | 2.98 × 10−2 | |
| E1BYS6 | Protein kinase C | PKN2 | 20.82 | 4.38 | 5.94 × 10−5 | |
| A0A1D5P5K6 | PHD-type domain-containing protein | TCF20 | 17.10 | 4.10 | 1.49 × 10−6 | |
| E1BU50 | Uncharacterized protein | LOC419409 | 17.00 | 4.09 | 3.21 × 10−2 | |
| A0A1D5PJI9 | Uncharacterized protein | VPS13A | 16.70 | 4.06 | 7.82 × 10−3 | |
| F1NZ61 | ZnMc domain-containing protein | MMP27 | 15.32 | 3.94 | 7.70 × 10−8 | |
| F1NG87 | TPR_REGION domain-containing protein | TTC28 | 15.03 | 3.91 | 2.05 × 10−6 | |
| F1P186 | Aa_trans domain-containing protein | SLC38A7 | 14.81 | 3.89 | 1.24 × 10−6 | |
| E1C1H9 | PLD phosphodiesterase domain-containing protein | PLD5 | 14.58 | 3.87 | 9.52 × 10−3 | |
| E1BQX8 | GRIP domain-containing protein | GOLGA1 | 11.87 | 3.57 | 9.02 × 10−5 | |
| E1BSS2 | Ubiquitin specific peptidase 53 | USP53 | 7.44 | 2.90 | 3.05 × 10−2 | |
| A0A1L1RVC2 | hSH3 domain-containing protein | C8H1orf168 | 6.94 | 2.80 | 2.96 × 10−2 | |
| A0A1L1RJ96 | FSA_C domain-containing protein | KIAA1109 | 6.72 | 2.75 | 3.96 × 10−2 | |
| A0A1D5P4T1 | Uncharacterized protein | SYTL2 | 6.60 | 2.72 | 3.09 × 10−4 | |
| A0A3Q2UBZ4 | Uncharacterized protein | MSLNL | 5.35 | 2.42 | 1.61 × 10−3 | |
| E1C1W0 | Uncharacterized protein | TRERF1 | 5.23 | 2.39 | 2.09 × 10−6 | |
| A0A3Q3A731 | Uncharacterized protein | - | 5.08 | 2.35 | 9.16 × 10−6 | |
| A0A1D5P3H2 | Guanylate cyclase | LOC107055115 | 3.20 | 1.68 | 4.59 × 10−3 | |
| A0A1D5PHD3 | C2H2-type domain-containing protein | ZNF318 | 3.09 | 1.63 | 8.23 × 10−4 | |
| O42252 | LIM domain-binding protein 1 | LDB1 | 2.63 | 1.39 | 1.18 × 10−3 | |
| Q2PC93 | SCO-spondin | SSPO | 2.01 | 1.00 | 1.81 × 10−2 | |
| A0A3Q2U7G6 | AF-4_C domain-containing protein | AFF1 | 2.00 | 1.00 | 3.24 × 10−2 |
| Treatment Group | Accession | Protein Name | Gene Symbol | Abundance Ratio | Log2 Abundance Ratio | p-Value |
|---|---|---|---|---|---|---|
| AIV (36 proteins) | R4GH67 | T-box_assoc domain-containing protein | EOMES | 0.01 | −6.64 | 3.38 × 10−10 |
| A0A1D5PVY2 | Uncharacterized protein | EHMT1 | 0.01 | −6.64 | 1.35 × 10−6 | |
| E1C1D1 | Annexin | ANXA7 | 0.02 | −6.03 | 2.43 × 10−5 | |
| E1C5B4 | Enhancer of polycomb homolog | EPC1 | 0.05 | −4.36 | 9.67 × 10−5 | |
| F1NJM6 | Uncharacterized protein | LOC100857368; ZCCHC6 | 0.05 | −4.21 | 4.31 × 10−7 | |
| A0A3Q2TTI8 | Zinc finger protein 644 | ZNF644 | 0.06 | −4.05 | 3.77 × 10−2 | |
| A0A1D5PPP1 | Uncharacterized protein | RGS14 | 0.06 | −3.98 | 3.40 × 10−5 | |
| A0A1D5PQ57 | Phosphoinositide phospholipase C | PLCE1 | 0.07 | −3.84 | 7.81 × 10−3 | |
| E1BU62 | DOP1 leucine zipper like protein B | 0.07 | −3.78 | 1.33 × 10−4 | ||
| F1NY55 | LEM domain-containing protein | ANKLE2 | 0.08 | −3.63 | 4.01 × 10−2 | |
| E1C8W5 | Uncharacterized protein | CHST15 | 0.1 | −3.27 | 1.75 × 10−4 | |
| F1NPH9 | OMPdecase | UMPS | 0.11 | −3.21 | 7.57 × 10−3 | |
| E1C8S3 | S1 motif domain-containing protein | SRBD1 | 0.11 | −3.13 | 1.27 × 10−7 | |
| A0A1D5PJ72 | Capping protein inhibiting regulator of actin dynamics | 0.12 | −3.08 | 1.46 × 10−5 | ||
| A0A1D5P7I8 | Zinc finger protein 516 | ZNF516 | 0.12 | −3.03 | 1.01 × 10−3 | |
| A0A3Q2U3A1 | Uncharacterized protein | 0.13 | −2.92 | 1.53 × 10−2 | ||
| A0A3Q3ANH5 | Uncharacterized protein | 0.16 | −2.69 | 5.39 × 10−4 | ||
| E1BQG1 | Uncharacterized protein | TNRC6B | 0.16 | −2.66 | 2.82 × 10−4 | |
| A0A1D5P251 | Uncharacterized protein | MAP1S | 0.16 | −2.66 | 4.85 × 10−2 | |
| F1NF87 | Exocyst complex component 5 | EXOC5 | 0.16 | −2.62 | 6.70 × 10−4 | |
| A0A3Q2UHA1 | Uncharacterized protein | FAAP100 | 0.17 | −2.59 | 2.83 × 10−2 | |
| A0A1L1RX59 | Diadenosine tetraphosphate synthetase | GARS | 0.17 | −2.56 | 3.52 × 10−3 | |
| R4GGE1 | Uncharacterized protein | SATB1 | 0.19 | −2.41 | 5.86 × 10−4 | |
| A0A3Q3A6T8 | Uncharacterized protein | ALMS1 | 0.21 | −2.24 | 2.06 × 10−2 | |
| A0A1D5PPH7 | UnbV_ASPIC domain-containing protein | CRTAC1 | 0.22 | −2.2 | 1.03 × 10−3 | |
| A0A1D5NU15 | Uncharacterized protein | MAGI1 | 0.24 | −2.04 | 1.43 × 10−3 | |
| F1NEI8 | Poly(A)-specific ribonuclease PARN | PARN | 0.29 | −1.8 | 2.59 × 10−4 | |
| E1BYA8 | Uncharacterized protein | ERCC6 | 0.29 | −1.78 | 3.96 × 10−6 | |
| A0A1D5P3H2 | Guanylate cyclase | LOC107055115 | 0.31 | −1.67 | 2.09 × 10−2 | |
| A0A3Q2TSU2 | HECT-type E3 ubiquitin transferase | NEDD4L | 0.35 | −1.53 | 6.02 × 10−3 | |
| A0A1D5P0N4 | Transcriptional activator Myb | MYB | 0.39 | −1.38 | 7.61 × 10−3 | |
| A0A3Q2U624 | Uncharacterized protein | WDR62 | 0.39 | −1.38 | 3.58 × 10−2 | |
| A0A1D5P124 | Uncharacterized protein | ANK2 | 0.39 | −1.34 | 4.80 × 10−2 | |
| A0A1D5PCT6 | Kinesin-like protein | KIF23 | 0.47 | −1.08 | 1.56 × 10−2 | |
| H9KYN7 | Peptidase S1 domain-containing protein | LOC431235; CTRB2; LOC100859877; CTRB1 | 0.47 | −1.08 | 4.57 × 10−2 | |
| F1NWT3 | F-box domain-containing protein | FBXO5 | 0.48 | −1.06 | 1.20 × 10−2 | |
| LPS (54 proteins) | A0A1D5NUX8 | Uncharacterized protein | SLC25A10 | 0.01 | −6.64 | 5.15 × 10−13 |
| A0A1D5P8P3 | Collagen IV NC1 domain-containing protein | COL4A1 | 0.01 | −6.64 | 2.19 × 10−10 | |
| R4GH67 | T-box_assoc domain-containing protein | EOMES | 0.01 | −6.64 | 4.54 × 10−9 | |
| A0A1D5PCP1 | Uncharacterized protein | ERICH3 | 0.01 | −6.64 | 3.02 × 10−5 | |
| F1NN69 | Beta-1,4-N-acetylgalactosaminyltransferase | B4GALNT3 | 0.01 | −6.26 | 2.38 × 10−7 | |
| A0A3Q2TZW2 | Transcription initiation factor TFIID subunit | RHOGL | 0.02 | −6.06 | 8.70 × 10−4 | |
| A0A1D5NZF5 | BHLH domain-containing protein | USF3 | 0.02 | −5.62 | 1.45 × 10−5 | |
| E1C7T1 | SERPIN domain-containing protein | SERPINA1; SPIA1 | 0.02 | −5.39 | 5.88 × 10−8 | |
| A0A3Q2UIH4 | Zyxin | ZYX | 0.03 | −5.25 | 6.78 × 10−4 | |
| F1NT94 | Histone acetyltransferase | KAT6A | 0.03 | −5.12 | 4.61 × 10−4 | |
| F1NY55 | LEM domain-containing protein | ANKLE2 | 0.03 | −5.07 | 1.07 × 10−2 | |
| F1NLF0 | Uncharacterized protein | EPS15 | 0.03 | −4.88 | 1.16 × 10−4 | |
| E1BU88 | Treslin_N domain-containing protein | C10H15orf42; TICRR | 0.04 | −4.73 | 2.73 × 10−2 | |
| R4GLP0 | Cytochrome c oxidase polypeptide VIIc | COX7C | 0.04 | −4.67 | 3.42 × 10−3 | |
| E1BU62 | DOP1 leucine zipper like protein B | 0.05 | −4.24 | 5.30 × 10−4 | ||
| E1BTE7 | AAA domain-containing protein | TOR3A | 0.06 | −3.96 | 1.21 × 10−3 | |
| F1NV58 | Uncharacterized protein | SPTBN5 | 0.07 | −3.83 | 7.92 × 10−3 | |
| A0A3Q2U9U5 | Ig-like domain-containing protein | ILDR2 | 0.07 | −3.81 | 1.45 × 10−4 | |
| A0A1D5NW78 | Uncharacterized protein | SPAG17 | 0.07 | −3.76 | 1.30 × 10−4 | |
| A0A3Q3A6T8 | Uncharacterized protein | ALMS1 | 0.08 | −3.73 | 4.11 × 10−4 | |
| A0A1D5P0N4 | Transcriptional activator Myb | MYB | 0.08 | −3.69 | 8.29 × 10−6 | |
| E1C309 | Uncharacterized protein | SLC25A19 | 0.08 | −3.66 | 1.76 × 10−3 | |
| P20740 | Ovostatin | LOC396151; OVST | 0.08 | −3.66 | 4.13× 10−3 | |
| E1C1D1 | Annexin | ANXA7 | 0.08 | −3.64 | 4.26 × 10−4 | |
| A0A1D5PVP9 | Protogenin | PRTG | 0.09 | −3.53 | 2.84 × 10−2 | |
| E1C8S3 | S1 motif domain-containing protein | SRBD1 | 0.09 | −3.45 | 5.11 × 10−8 | |
| F1NDM4 | Origin recognition complex subunit 2 | ORC2 | 0.09 | −3.43 | 3.49 × 10−3 | |
| F1NJM6 | Uncharacterized protein | LOC100857368; ZCCHC6 | 0.1 | −3.4 | 1.16 × 10−6 | |
| F1NHH4 | Fibrillar collagen NC1 domain-containing protein | COL27A1 | 0.11 | −3.17 | 1.39 × 10−4 | |
| E1BQP5 | Uncharacterized protein | WDR72 | 0.12 | −3.11 | 1.17 × 10−2 | |
| A0A3Q2TSU2 | HECT-type E3 ubiquitin transferase | NEDD4L | 0.12 | −3.03 | 2.64 × 10−5 | |
| R4GGE1 | Uncharacterized protein | SATB1 | 0.15 | −2.71 | 1.19 × 10−3 | |
| A0A1D5PVY2 | Uncharacterized protein | EHMT1 | 0.16 | −2.65 | 8.64 × 10−3 | |
| E1C8W5 | Uncharacterized protein | CHST15 | 0.17 | −2.6 | 6.38 × 10−4 | |
| R4GFN5 | Uncharacterized protein | GRIN2C; LOC431090 | 0.17 | −2.55 | 3.10 × 10−3 | |
| F1NWT3 | F-box domain-containing protein | FBXO5 | 0.19 | −2.41 | 6.47 × 10−4 | |
| A0A1D5P124 | Uncharacterized protein | ANK2 | 0.19 | −2.4 | 1.12 × 10−2 | |
| E1C5B4 | Enhancer of polycomb homolog | EPC1 | 0.2 | −2.33 | 1.26 × 10−2 | |
| A0A3Q2U888 | Rhomboid domain-containing protein | RHBDF2 | 0.2 | −2.31 | 1.61 × 10−2 | |
| P08287 | Histone H1.11L | HIST1H111L; HIST1H1C | 0.21 | −2.26 | 1.73 × 10−3 | |
| A0A3Q2U624 | Uncharacterized protein | WDR62 | 0.22 | −2.21 | 5.68 × 10−3 | |
| A0A3Q2TXE9 | Uncharacterized protein | 0.22 | −2.17 | 7.11 × 10−6 | ||
| E1BQG1 | Uncharacterized protein | TNRC6B | 0.24 | −2.07 | 3.35 × 10−3 | |
| R4GIZ6 | TIGAR | C1H12orf5; TIGAR | 0.24 | −2.06 | 1.72 × 10−4 | |
| H9KYN7 | Peptidase S1 domain-containing protein | LOC431235; CTRB2; LOC100859877; CTRB1 | 0.25 | −2.02 | 6.02 × 10−4 | |
| A0A1D5PPH7 | UnbV_ASPIC domain-containing protein | CRTAC1 | 0.27 | −1.89 | 2.97 × 10−3 | |
| A0A3Q3AGH4 | AIG1-type G domain-containing protein | GIMAP7L5 | 0.29 | −1.78 | 3.10 × 10−4 | |
| E1C908 | Uncharacterized protein | FYB | 0.31 | −1.68 | 1.07 × 10−2 | |
| A0A3Q3ANH5 | Uncharacterized protein | 0.32 | −1.64 | 4.30 × 10−2 | ||
| E1BYA8 | Uncharacterized protein | ERCC6 | 0.32 | −1.63 | 9.60 × 10−6 | |
| A0A1D5PE26 | Uncharacterized protein | DENND4A | 0.36 | −1.47 | 2.83 × 10−3 | |
| F1NEI8 | Poly(A)-specific ribonuclease PARN | PARN | 0.39 | −1.36 | 1.86 × 10−3 | |
| A0A1L1RKD5 | Uncharacterized protein | WISP1 | 0.39 | −1.36 | 1.33 × 10−2 | |
| Q5F393 | Nuclear receptor coactivator | NCOA1 | 0.41 | −1.27 | 1.70 × 10−2 | |
| PolyI:C (52 proteins) | R4GH67 | T-box_assoc domain-containing protein | EOMES | 0.01 | −6.64 | 1.93 × 10−10 |
| E1BU62 | DOP1 leucine zipper like protein B | 0.01 | −6.48 | 2.51 × 10−5 | ||
| E1C309 | Uncharacterized protein | SLC25A19 | 0.02 | −5.61 | 4.58 × 10−4 | |
| A0A1D5NY78 | SCD domain-containing protein | STAG1 | 0.03 | −5.07 | 5.22 × 10−6 | |
| A0A3Q2UFS2 | AIP3 domain-containing protein | SRCIN1 | 0.03 | −4.9 | 3.68 × 10−4 | |
| A0A1D5PES4 | Receptor protein-tyrosine kinase | EPHA5 | 0.04 | −4.85 | 1.01 × 10−4 | |
| A0A1D5PVY2 | Uncharacterized protein | EHMT1 | 0.04 | −4.65 | 9.19 × 10−5 | |
| A0A1D5PQ57 | Phosphoinositide phospholipase C | PLCE1 | 0.04 | −4.6 | 2.26 × 10−3 | |
| A0A3Q3A9V6 | LIM zinc-binding domain-containing protein | ZNF185L | 0.04 | −4.58 | 2.73 × 10−2 | |
| E1C264 | Uncharacterized protein | CDH9 | 0.05 | −4.28 | 1.09 × 10−2 | |
| E1C1D1 | Annexin | ANXA7 | 0.05 | −4.25 | 1.62 × 10−4 | |
| A0A3Q2U9U5 | Ig-like domain-containing protein | ILDR2 | 0.06 | −4.19 | 4.27 × 10−5 | |
| A0A1D5NVF3 | Sorting nexin-17 | SNX17 | 0.06 | −4.09 | 1.12 × 10−2 | |
| A0A1D5P8D2 | JmjC domain-containing protein | JMJD1C | 0.06 | −4.03 | 9.26 × 10−4 | |
| F1NLF0 | Uncharacterized protein | EPS15 | 0.07 | −3.95 | 1.18 × 10−4 | |
| A0A5H1ZRJ7 | Collagen alpha-2(I) chain | COL1A2 | 0.07 | −3.92 | 9.29 × 10−4 | |
| A0A3Q2TSU2 | HECT-type E3 ubiquitin transferase | NEDD4L | 0.07 | −3.87 | 2.01 × 10−5 | |
| E1BQP5 | Uncharacterized protein | WDR72 | 0.07 | −3.81 | 2.45 × 10−3 | |
| F1NCA2 | Glycerol-3-phosphate dehydrogenase | GPD2 | 0.08 | −3.66 | 9.44 × 10−6 | |
| A0A1D5PJ72 | Capping protein inhibiting regulator of actin dynamics | 0.08 | −3.6 | 3.91 × 10−6 | ||
| E1BQF4 | TGF-beta receptor type-2 | AMHR2 | 0.09 | −3.55 | 2.75 × 10−3 | |
| A0A3Q2TU08 | Uncharacterized protein | CDC5L | 0.09 | −3.48 | 1.71 × 10−2 | |
| F1NV58 | Uncharacterized protein | SPTBN5 | 0.1 | −3.38 | 2.55 × 10−2 | |
| E1BTE7 | AAA domain-containing protein | TOR3A | 0.1 | −3.35 | 7.38 × 10−4 | |
| A0A3Q2U624 | Uncharacterized protein | WDR62 | 0.1 | −3.35 | 1.55 × 10−3 | |
| E1C5B4 | Enhancer of polycomb homolog | EPC1 | 0.12 | −3.12 | 2.24 × 10−3 | |
| A0A1D5P1A1 | Uncharacterized protein | C5H11ORF9 | 0.12 | −3.08 | 2.26 × 10−2 | |
| A0A3Q2UHA1 | Uncharacterized protein | FAAP100 | 0.12 | −3.03 | 4.32 × 10−2 | |
| R4GGE1 | Uncharacterized protein | SATB1 | 0.13 | −2.91 | 1.04 × 10−4 | |
| P08287 | Histone H1.11L | HIST1H111L; HIST1H1C | 0.14 | −2.86 | 2.58 × 10−4 | |
| A0A3Q2UG75 | Ras-responsive element-binding protein 1 | RREB1 | 0.14 | −2.82 | 3.39 × 10−2 | |
| E1C8S3 | S1 motif domain-containing protein | SRBD1 | 0.16 | −2.69 | 7.57 × 10−7 | |
| A0A3Q3ANH5 | Uncharacterized protein | 0.17 | −2.53 | 3.83 × 10−4 | ||
| F1NWT3 | F-box domain-containing protein | FBXO5 | 0.18 | −2.5 | 1.02 × 10−4 | |
| A0A3Q2U0V9 | Uncharacterized protein | SLC11A2 | 0.18 | −2.47 | 1.82 × 10−2 | |
| E1BYA8 | Uncharacterized protein | ERCC6 | 0.18 | −2.45 | 1.51 × 10−7 | |
| A0A1D5P0N4 | Transcriptional activator Myb | MYB | 0.22 | −2.22 | 1.91 × 10−3 | |
| A0A3Q3A6T8 | Uncharacterized protein | ALMS1 | 0.22 | −2.22 | 2.09 × 10−2 | |
| A0A1D5PD16 | Uncharacterized protein | DNAH10 | 0.23 | −2.14 | 2.01 × 10−3 | |
| A0A1L1RX59 | Diadenosine tetraphosphate synthetase | GARS | 0.24 | −2.09 | 2.08 × 10−2 | |
| A0A1D5PBZ9 | Protein-tyrosine-phosphatase | PTPRB | 0.24 | −2.05 | 1.48 × 10−3 | |
| F1P5A5 | 28S ribosomal protein S31, mitochondrial | MRPS31 | 0.26 | −1.97 | 6.18 × 10−3 | |
| E1BQG1 | Uncharacterized protein | TNRC6B | 0.26 | −1.92 | 4.09 × 10−3 | |
| A0A3Q2U888 | Rhomboid domain-containing protein | RHBDF2 | 0.27 | −1.9 | 4.90 × 10−2 | |
| A0A1D5P7I8 | Zinc finger protein 516 | ZNF516 | 0.3 | −1.74 | 1.50 × 10−2 | |
| H9KYN7 | Peptidase S1 domain-containing protein | LOC431235; CTRB2; LOC100859877; CTRB1 | 0.31 | −1.67 | 3.00 × 10−3 | |
| Q5F393 | Nuclear receptor coactivator | NCOA1 | 0.35 | −1.54 | 7.34 × 10−3 | |
| A0A1D5PGG8 | Nucleolar complex protein 3 homolog | NOC3L | 0.36 | −1.48 | 6.74 × 10−3 | |
| A0A3Q2U9J3 | Ubiquitinyl hydrolase 1 | OTUD7A | 0.38 | −1.38 | 9.29 × 10−3 | |
| F1NQ24 | Uncharacterized protein | DENND4C | 0.39 | −1.37 | 5.08 × 10−3 | |
| F1NEI8 | Poly(A)-specific ribonuclease PARN | PARN | 0.42 | −1.25 | 4.00 × 10−3 | |
| A0A3Q2TXE9 | Uncharacterized protein | 0.47 | −1.09 | 5.90 × 10−3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Dowd, K.; Sánchez, L.; Ben Salem, J.; Beaudry, F.; Barjesteh, N. Characterization of the Role of Extracellular Vesicles Released from Chicken Tracheal Cells in the Antiviral Responses against Avian Influenza Virus. Membranes 2022, 12, 53. https://doi.org/10.3390/membranes12010053
O’Dowd K, Sánchez L, Ben Salem J, Beaudry F, Barjesteh N. Characterization of the Role of Extracellular Vesicles Released from Chicken Tracheal Cells in the Antiviral Responses against Avian Influenza Virus. Membranes. 2022; 12(1):53. https://doi.org/10.3390/membranes12010053
Chicago/Turabian StyleO’Dowd, Kelsey, Laura Sánchez, Jennifer Ben Salem, Francis Beaudry, and Neda Barjesteh. 2022. "Characterization of the Role of Extracellular Vesicles Released from Chicken Tracheal Cells in the Antiviral Responses against Avian Influenza Virus" Membranes 12, no. 1: 53. https://doi.org/10.3390/membranes12010053
APA StyleO’Dowd, K., Sánchez, L., Ben Salem, J., Beaudry, F., & Barjesteh, N. (2022). Characterization of the Role of Extracellular Vesicles Released from Chicken Tracheal Cells in the Antiviral Responses against Avian Influenza Virus. Membranes, 12(1), 53. https://doi.org/10.3390/membranes12010053