
Red Blood Cell Proteasome in Beta-Thalassemia Trait: Topology of Activity and Networking in Blood Bank Conditions

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. Sample Fractionation
2.3. Proteasome Activities
2.4. Physiological and Proteomic Parameters
2.5. Statistical and Network Analysis
3. Results
3.1. Time-Course Evaluation of Proteasome Activity during RBC Storage
3.2. Proteasome Activity in βThal+ versus Control RBCs
3.3. Correlations between Proteasome Activities and Reactive Oxygen Species in βThal+ and Control RBCs
3.4. Biological Networks
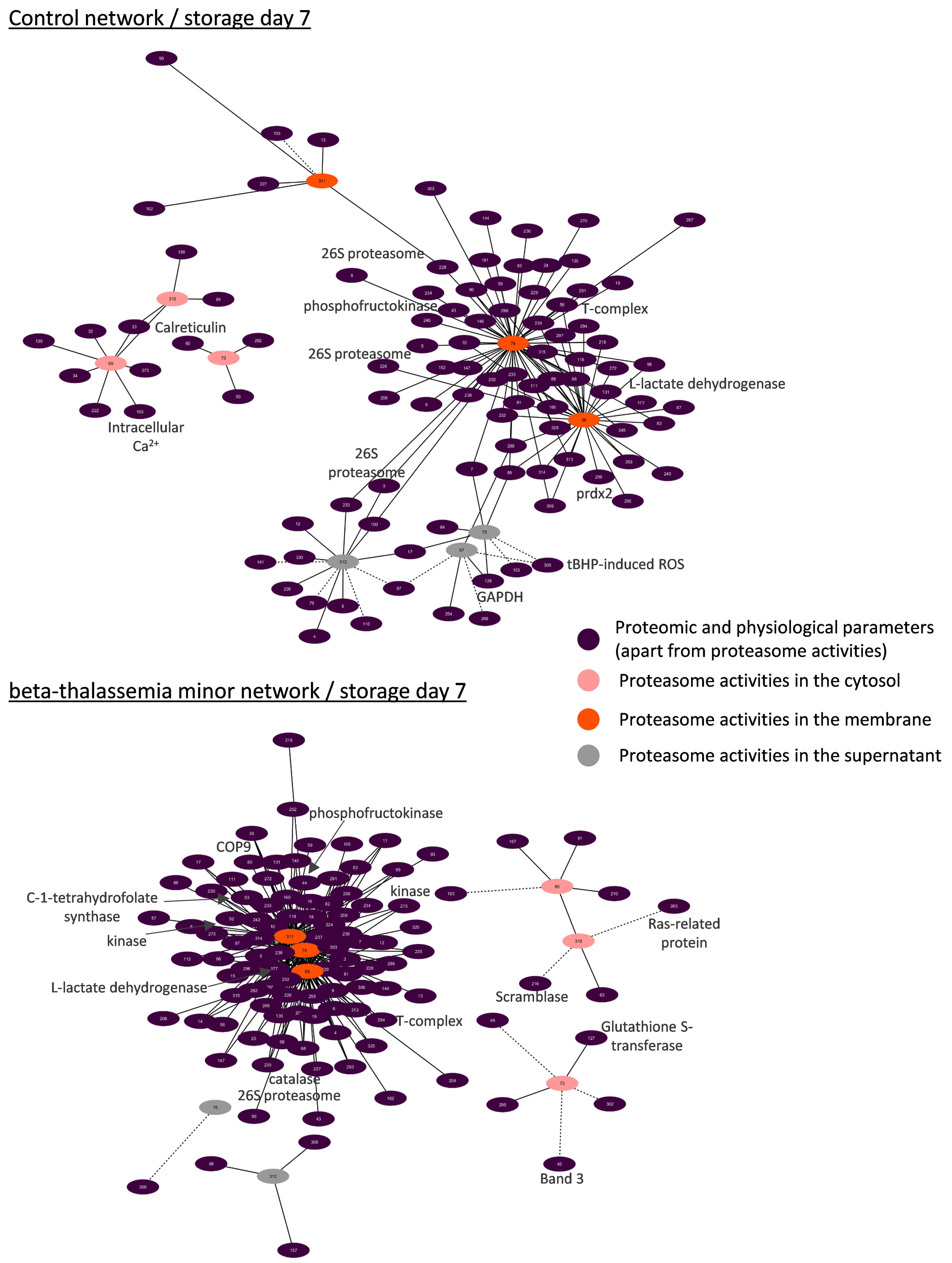
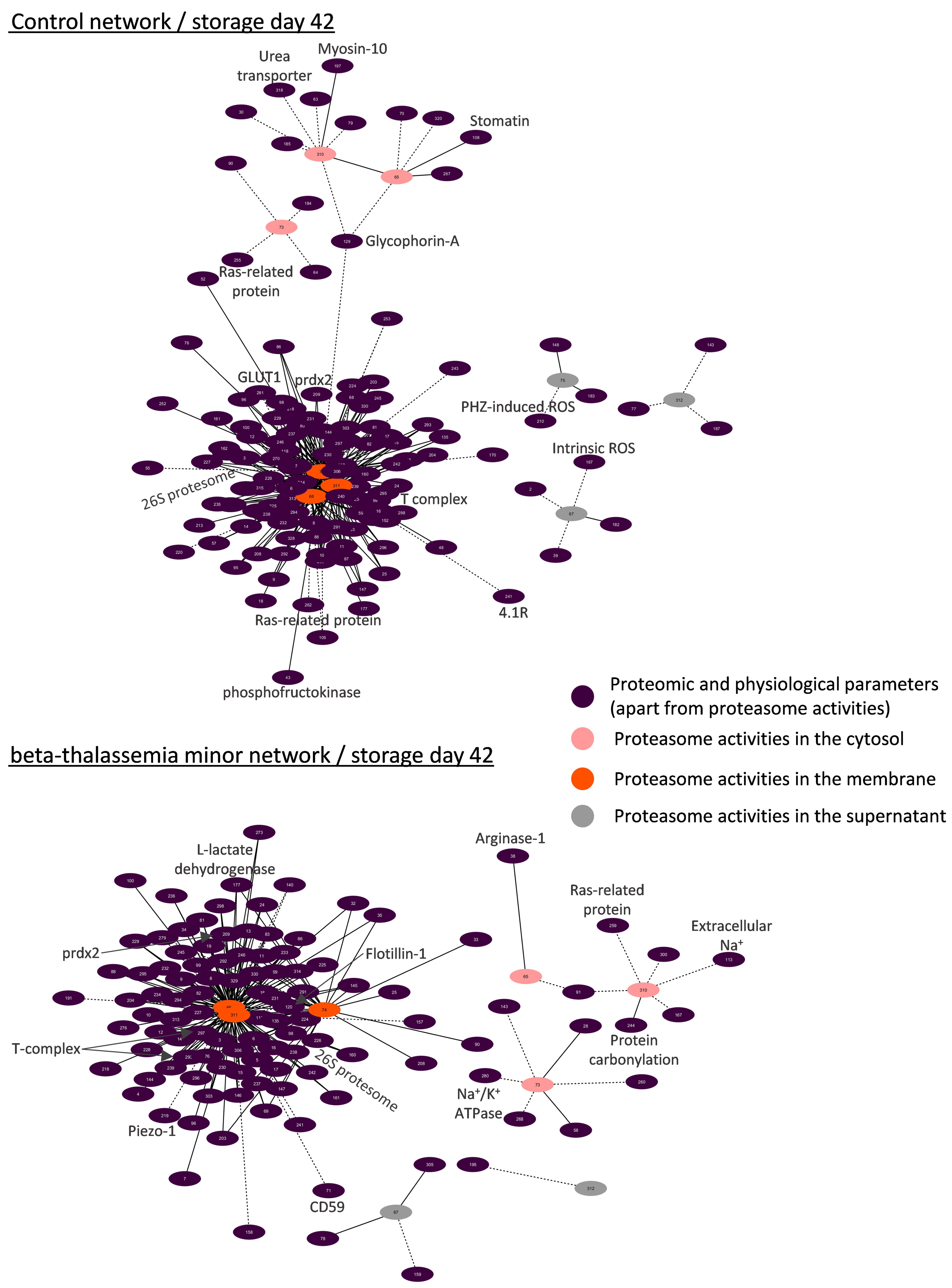
3.4.1. Biological Networking of Proteomic and Physiological Parameters of RBCs
3.4.2. Biological Networking of 20S Proteasome
3.4.3. Biological Networks of Proteasome Activities
4. Discussion
4.1. The Discrete Spatiotemporal Proteasome Activity Profile in Stored RBCs
4.2. Time-Dependent Cross-Talk between Proteasome Activities and ROS Levels
4.3. Storage Induces Higher Levels of Proteasome Activity in the Membrane of βThal+ RBCs
4.4. Biological Networking of Proteasome and Proteasome Activities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Livneh, I.; Cohen-Kaplan, V.; Cohen-Rosenzweig, C.; Avni, N.; Ciechanover, A. The life cycle of the 26S proteasome: From birth, through regulation and function, and onto its death. Cell Res. 2016, 26, 869–885. [Google Scholar] [CrossRef]
- Rousseau, A.; Bertolotti, A. Regulation of proteasome assembly and activity in health and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Reuven, N.; Shaul, Y. 20S proteasomes and protein degradation “by default”. Bioessays 2006, 28, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nissan, G.; Sharon, M. Regulating the 20S proteasome ubiquitin-independent degradation pathway. Biomolecules 2014, 4, 862–884. [Google Scholar] [CrossRef] [PubMed]
- Olshina, M.A.; Ben-Nissan, G.; Sharon, M. Functional regulation of proteins by 20S proteasome proteolytic processing. Cell Cycle 2018, 17, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Dixon, J.E. The 26S proteasome: A cell cycle regulator regulated by cell cycle. Cell Cycle 2016, 15, 875–876. [Google Scholar] [CrossRef]
- Pickering, A.M.; Koop, A.L.; Teoh, C.Y.; Ermak, G.; Grune, T.; Davies, K.J. The immunoproteasome, the 20S proteasome and the PA28alphabeta proteasome regulator are oxidative-stress-adaptive proteolytic complexes. Biochem. J. 2010, 432, 585–594. [Google Scholar] [CrossRef]
- Shang, F.; Taylor, A. Ubiquitin-proteasome pathway and cellular responses to oxidative stress. Free Radic. Biol. Med. 2011, 51, 5–16. [Google Scholar] [CrossRef]
- Naujokat, C.; Hoffmann, S. Role and function of the 26S proteasome in proliferation and apoptosis. Lab. Investig. 2002, 82, 965–980. [Google Scholar] [CrossRef]
- Goodman, S.R.; Kurdia, A.; Ammann, L.; Kakhniashvili, D.; Daescu, O. The human red blood cell proteome and interactome. Exp. Biol. Med. 2007, 232, 1391–1408. [Google Scholar] [CrossRef]
- Neelam, S.; Kakhniashvili, D.G.; Wilkens, S.; Levene, S.D.; Goodman, S.R. Functional 20S proteasomes in mature human red blood cells. Exp. Biol. Med. 2011, 236, 580–591. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.J. Degradation of oxidized proteins by the 20S proteasome. Biochimie 2001, 83, 301–310. [Google Scholar] [CrossRef]
- Reinheckel, T.; Sitte, N.; Ullrich, O.; Kuckelkorn, U.; Davies, K.J.; Grune, T. Comparative resistance of the 20S and 26S proteasome to oxidative stress. Biochem. J. 1998, 335 Pt 3, 637–642. [Google Scholar] [CrossRef]
- Pesciotta, E.N.; Lam, H.S.; Kossenkov, A.; Ge, J.; Showe, L.C.; Mason, P.J.; Bessler, M.; Speicher, D.W. In-Depth, Label-Free Analysis of the Erythrocyte Cytoplasmic Proteome in Diamond Blackfan Anemia Identifies a Unique Inflammatory Signature. PLoS ONE 2015, 10, e0140036. [Google Scholar] [CrossRef] [PubMed]
- von Lohneysen, K.; Scott, T.M.; Soldau, K.; Xu, X.; Friedman, J.S. Assessment of the red cell proteome of young patients with unexplained hemolytic anemia by two-dimensional differential in-gel electrophoresis (DIGE). PLoS ONE 2012, 7, e34237. [Google Scholar] [CrossRef][Green Version]
- Basu, A.; Saha, S.; Karmakar, S.; Chakravarty, S.; Banerjee, D.; Dash, B.P.; Chakrabarti, A. 2D DIGE based proteomics study of erythrocyte cytosol in sickle cell disease: Altered proteostasis and oxidative stress. Proteomics 2013, 13, 3233–3242. [Google Scholar] [CrossRef]
- Bosman, G.J.; Lasonder, E.; Luten, M.; Roerdinkholder-Stoelwinder, B.; Novotny, V.M.; Bos, H.; De Grip, W.J. The proteome of red cell membranes and vesicles during storage in blood bank conditions. Transfusion 2008, 48, 827–835. [Google Scholar] [CrossRef]
- Tzounakas, V.L.; Dzieciatkowska, M.; Anastasiadi, A.T.; Karadimas, D.G.; Vergaki, A.; Siourounis, P.; Stamoulis, K.; Papassideri, I.S.; Kriebardis, A.G.; D’Alessandro, A.; et al. Red cell proteasome modulation by storage, redox metabolism and transfusion. Blood Transfus. 2020. [Google Scholar] [CrossRef]
- Geng, Q.; Romero, J.; Saini, V.; Patel, M.B.; Majetschak, M. Extracellular 20S proteasomes accumulate in packed red blood cell units. Vox Sang. 2009, 97, 273–274. [Google Scholar] [CrossRef]
- Tzounakas, V.L.; Anastasiadi, A.T.; Drossos, P.V.; Karadimas, D.G.; Valsami, S.E.; Stamoulis, K.E.; Papassideri, I.S.; Politou, M.; Antonelou, M.H.; Kriebardis, A.G. Sex-related aspects of the red blood cell storage lesion. Blood Transfus. 2021, 19, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Teran, M.M.; Monaco, M.E.; Lazarte, S.S.; Haro, C.; Ledesma Achem, E.; Asensio, N.A.; Isse, B.A. Genetic Regulation of Redox Balance in beta-Thalassemia Trait. Hemoglobin 2020, 44, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Anastasiadi, A.T.; Stefanoni, D.; Cendali, F.; Bertolone, L.; Gamboni, F.; Dzieciatkowska, M.; Rousakis, P.; Vergaki, A.; Soulakis, V.; et al. beta-thalassemia minor is a beneficial determinant of red blood cell storage lesion. Haematologica 2021. [Google Scholar] [CrossRef] [PubMed]
- Tzounakas, V.L.; Anastasiadi, A.T.; Dzieciatkowska, M.; Karadimas, D.G.; Stamoulis, K.; Papassideri, I.S.; Hansen, K.C.; D’Alessandro, A.; Kriebardis, A.G.; Antonelou, M.H. Proteome of Stored RBC Membrane and Vesicles from Heterozygous Beta Thalassemia Donors. Int. J. Mol. Sci. 2021, 22, 3369. [Google Scholar] [CrossRef] [PubMed]
- Kriebardis, A.G.; Antonelou, M.H.; Stamoulis, K.E.; Economou-Petersen, E.; Margaritis, L.H.; Papassideri, I.S. RBC-derived vesicles during storage: Ultrastructure, protein composition, oxidation, and signaling components. Transfusion 2008, 48, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Homma, T.; Kurahashi, T.; Lee, J.; Kang, E.S.; Fujii, J. SOD1 deficiency decreases proteasomal function, leading to the accumulation of ubiquitinated proteins in erythrocytes. Arch. Biochem. Biophys. 2015, 583, 65–72. [Google Scholar] [CrossRef]
- Antonelou, M.H.; Tzounakas, V.L.; Velentzas, A.D.; Stamoulis, K.E.; Kriebardis, A.G.; Papassideri, I.S. Effects of pre-storage leukoreduction on stored red blood cells signaling: A time-course evaluation from shape to proteome. J. Proteom. 2012, 76, 220–238. [Google Scholar] [CrossRef]
- Bardyn, M.; Tissot, J.D.; Prudent, M. Oxidative stress and antioxidant defenses during blood processing and storage of erythrocyte concentrates. Transfus. Clin. Biol. 2018, 25, 96–100. [Google Scholar] [CrossRef]
- Reisz, J.A.; Wither, M.J.; Dzieciatkowska, M.; Nemkov, T.; Issaian, A.; Yoshida, T.; Dunham, A.J.; Hill, R.C.; Hansen, K.C.; D’Alessandro, A. Oxidative modifications of glyceraldehyde 3-phosphate dehydrogenase regulate metabolic reprogramming of stored red blood cells. Blood 2016, 128, e32–e42. [Google Scholar] [CrossRef]
- Fujii, J.; Homma, T.; Kobayashi, S.; Warang, P.; Madkaikar, M.; Mukherjee, M.B. Erythrocytes as a preferential target of oxidative stress in blood. Free Radic. Res. 2021, 1–19. [Google Scholar] [CrossRef]
- Delobel, J.; Prudent, M.; Tissot, J.D.; Lion, N. Proteomics of the red blood cell carbonylome during blood banking of erythrocyte concentrates. Proteom. Clin. Appl. 2016, 10, 257–266. [Google Scholar] [CrossRef]
- VerPlank, J.J.S.; Goldberg, A.L. Regulating protein breakdown through proteasome phosphorylation. Biochem. J. 2017, 474, 3355–3371. [Google Scholar] [CrossRef]
- Conconi, M.; Petropoulos, I.; Emod, I.; Turlin, E.; Biville, F.; Friguet, B. Protection from oxidative inactivation of the 20S proteasome by heat-shock protein 90. Biochem. J. 1998, 333 Pt 2, 407–415. [Google Scholar] [CrossRef]
- Biondani, A.; Turrini, F.; Carta, F.; Matte, A.; Filippini, A.; Siciliano, A.; Beuzard, Y.; De Franceschi, L. Heat-shock protein-27, -70 and peroxiredoxin-II show molecular chaperone function in sickle red cells: Evidence from transgenic sickle cell mouse model. Proteom. Clin. Appl. 2008, 2, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Strader, M.B.; Jana, S.; Meng, F.; Heaven, M.R.; Shet, A.S.; Thein, S.L.; Alayash, A.I. Post-translational modification as a response to cellular stress induced by hemoglobin oxidation in sickle cell disease. Sci. Rep. 2020, 10, 14218. [Google Scholar] [CrossRef] [PubMed]
- Warang, P.; Homma, T.; Pandya, R.; Sawant, A.; Shinde, N.; Pandey, D.; Fujii, J.; Madkaikar, M.; Mukherjee, M.B. Potential involvement of ubiquitin-proteasome system dysfunction associated with oxidative stress in the pathogenesis of sickle cell disease. Br. J. Haematol. 2018, 182, 559–566. [Google Scholar] [CrossRef]
- Khandros, E.; Thom, C.S.; D’Souza, J.; Weiss, M.J. Integrated protein quality-control pathways regulate free alpha-globin in murine beta-thalassemia. Blood 2012, 119, 5265–5275. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, A.; Righetti, P.G.; Zolla, L. The red blood cell proteome and interactome: An update. J. Proteome Res. 2010, 9, 144–163. [Google Scholar] [CrossRef]
- Seelig, A.; Boes, B.; Kloetzel, P.M. Characterization of mouse proteasome subunit MC3 and identification of proteasome subtypes with different cleavage characteristics. Proteasome subunits, proteasome subpopulations. Enzyme Protein 1993, 47, 330–342. [Google Scholar] [CrossRef]
- Shashova, E.E.; Kolegova, E.S.; Zav’yalov, A.A.; Slonimskaya, E.M.; Kondakova, I.V. Changes in the Activity of Proteasomes and Calpains in Metastases of Human Lung Cancer and Breast Cancer. Bull. Exp. Biol. Med. 2017, 163, 486–489. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.; Shu, X.; Chen, Q.; Wei, T.; Wang, H.; Wang, X.; Wu, Q.; Zhang, X.; Liu, X.; et al. Proteasome regulation by reversible tyrosine phosphorylation at the membrane. Oncogene 2021, 40, 1942–1956. [Google Scholar] [CrossRef] [PubMed]
- Coates, H.W.; Capell-Hattam, I.M.; Brown, A.J. The mammalian cholesterol synthesis enzyme squalene monooxygenase is proteasomally truncated to a constitutively active form. J. Biol. Chem. 2021, 296, 100731. [Google Scholar] [CrossRef]
- Oliva, J.; French, S.W.; Li, J.; Bardag-Gorce, F. Proteasome inhibitor treatment reduced fatty acid, triacylglycerol and cholesterol synthesis. Exp. Mol. Pathol. 2012, 93, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Dhungana, S.; Merrick, B.A.; Tomer, K.B.; Fessler, M.B. Quantitative proteomics analysis of macrophage rafts reveals compartmentalized activation of the proteasome and of proteasome-mediated ERK activation in response to lipopolysaccharide. Mol. Cell Proteom. 2009, 8, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Abriel, H.; Staub, O. Ubiquitylation of ion channels. Physiology 2005, 20, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Miroshnikova, Y.A.; Manet, S.; Li, X.; Wickstrom, S.A.; Faurobert, E.; Albiges-Rizo, C. Calcium signaling mediates a biphasic mechanoadaptive response of endothelial cells to cyclic mechanical stretch. Mol. Biol. Cell 2021, 32, mbcE21030106. [Google Scholar] [CrossRef]
- Wei, Q.; Xia, Y. Proteasome inhibition down-regulates endothelial nitric-oxide synthase phosphorylation and function. J. Biol. Chem. 2006, 281, 21652–21659. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Fan, G.; Shan, P.; Qiu, X.; Dong, S.; Liao, L.; Yu, C.; Wang, T.; Gu, X.; Li, Q.; et al. Regulation of energy homeostasis by the ubiquitin-independent REGgamma proteasome. Nat. Commun. 2016, 7, 12497. [Google Scholar] [CrossRef]
- Marcoux, G.; Laroche, A.; Hasse, S.; Bellio, M.; Mbarik, M.; Tamagne, M.; Allaeys, I.; Zufferey, A.; Levesque, T.; Rebetz, J.; et al. Platelet EVs contain an active proteasome involved in protein processing for antigen presentation via MHC-I molecules. Blood 2021. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Storage | ||||||
|---|---|---|---|---|---|---|
| Activities | 7 | 14 | 21 | 28 | 42 | |
| Cytosol | CH-like | 34 ± 8 | 29 ± 7 * | 24 ± 5 * | 24 ± 6 *,§ | 21 ± 5 *,§ |
| CASP-like | 21 ± 5 | 18 ± 5 * | 15 ± 5 *,§ | 14 ± 4 *,§ | 14 ± 4 *,§ | |
| TR-like | 44 ± 12 | 40 ± 10 | 31 ± 10 *,§ | 29 ± 8 *,§ | 30 ± 11 *,§ | |
| Membrane | CH-like | 42 ± 19 | 48 ± 21 | 55 ± 21 | 46 ± 21 | 39 ± 20 # |
| CASP-like | 40 ± 13 | 52 ± 16 | 81 ± 35 *,§ | 81 ± 29 *,§ | 63 ± 34 # | |
| TR-like | 29 ± 11 | 34 ± 9 | 50 ± 22 *,§ | 36 ± 18 # | 29 ± 14 # | |
| Supernatant | CH-like | 0.3 ± 0.1 | 0.3 ± 0.2 | 0.5 ± 0.3 | 0.5 ± 0.3 | 0.7 ± 0.3 *,# |
| CASP-like | 1.3 ± 0.5 | 1.7 ± 0.7 | 1.8 ± 0.5 * | 1.7 ± 0.7 * | 1.9 ± 0.6 * | |
| TR-like | 3.4 ± 1.0 | 3.6 ± 0.8 | 3.9 ± 1.2 | 4.0 ± 0.9 *,# | 4.5 ± 1.3 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anastasiadi, A.T.; Tzounakas, V.L.; Arvaniti, V.-Z.; Dzieciatkowska, M.; Stamoulis, K.; Lekka, M.E.; Papassideri, I.S.; D’Alessandro, A.; Kriebardis, A.G.; Antonelou, M.H. Red Blood Cell Proteasome in Beta-Thalassemia Trait: Topology of Activity and Networking in Blood Bank Conditions. Membranes 2021, 11, 716. https://doi.org/10.3390/membranes11090716
Anastasiadi AT, Tzounakas VL, Arvaniti V-Z, Dzieciatkowska M, Stamoulis K, Lekka ME, Papassideri IS, D’Alessandro A, Kriebardis AG, Antonelou MH. Red Blood Cell Proteasome in Beta-Thalassemia Trait: Topology of Activity and Networking in Blood Bank Conditions. Membranes. 2021; 11(9):716. https://doi.org/10.3390/membranes11090716
Chicago/Turabian StyleAnastasiadi, Alkmini T., Vassilis L. Tzounakas, Vasiliki-Zoi Arvaniti, Monika Dzieciatkowska, Konstantinos Stamoulis, Marilena E. Lekka, Issidora S. Papassideri, Angelo D’Alessandro, Anastasios G. Kriebardis, and Marianna H. Antonelou. 2021. "Red Blood Cell Proteasome in Beta-Thalassemia Trait: Topology of Activity and Networking in Blood Bank Conditions" Membranes 11, no. 9: 716. https://doi.org/10.3390/membranes11090716
APA StyleAnastasiadi, A. T., Tzounakas, V. L., Arvaniti, V.-Z., Dzieciatkowska, M., Stamoulis, K., Lekka, M. E., Papassideri, I. S., D’Alessandro, A., Kriebardis, A. G., & Antonelou, M. H. (2021). Red Blood Cell Proteasome in Beta-Thalassemia Trait: Topology of Activity and Networking in Blood Bank Conditions. Membranes, 11(9), 716. https://doi.org/10.3390/membranes11090716