
Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthesis of F16-Betulin Conjugate
2.2. Isolation of Rat Thymocytes and Flow Cytometry
2.3. Isolation of Rat Liver Mitochondria
2.4. Optical Microscopy Imaging of Mitochondrial Samples
2.5. Monitoring Optical Density of Mitochondrial Suspension
2.6. Determination of Mitochondrial Respiration and Oxidative Phosphorylation
2.7. Measuring Activity of Complexes of the Mitochondrial Electron Transport Chain (ETC)
2.8. Monitoring of Mitochondrial Membrane Potential
2.9. Production of H2O2 by Rat Liver Mitochondria
2.10. Statistical Analysis
3. Results
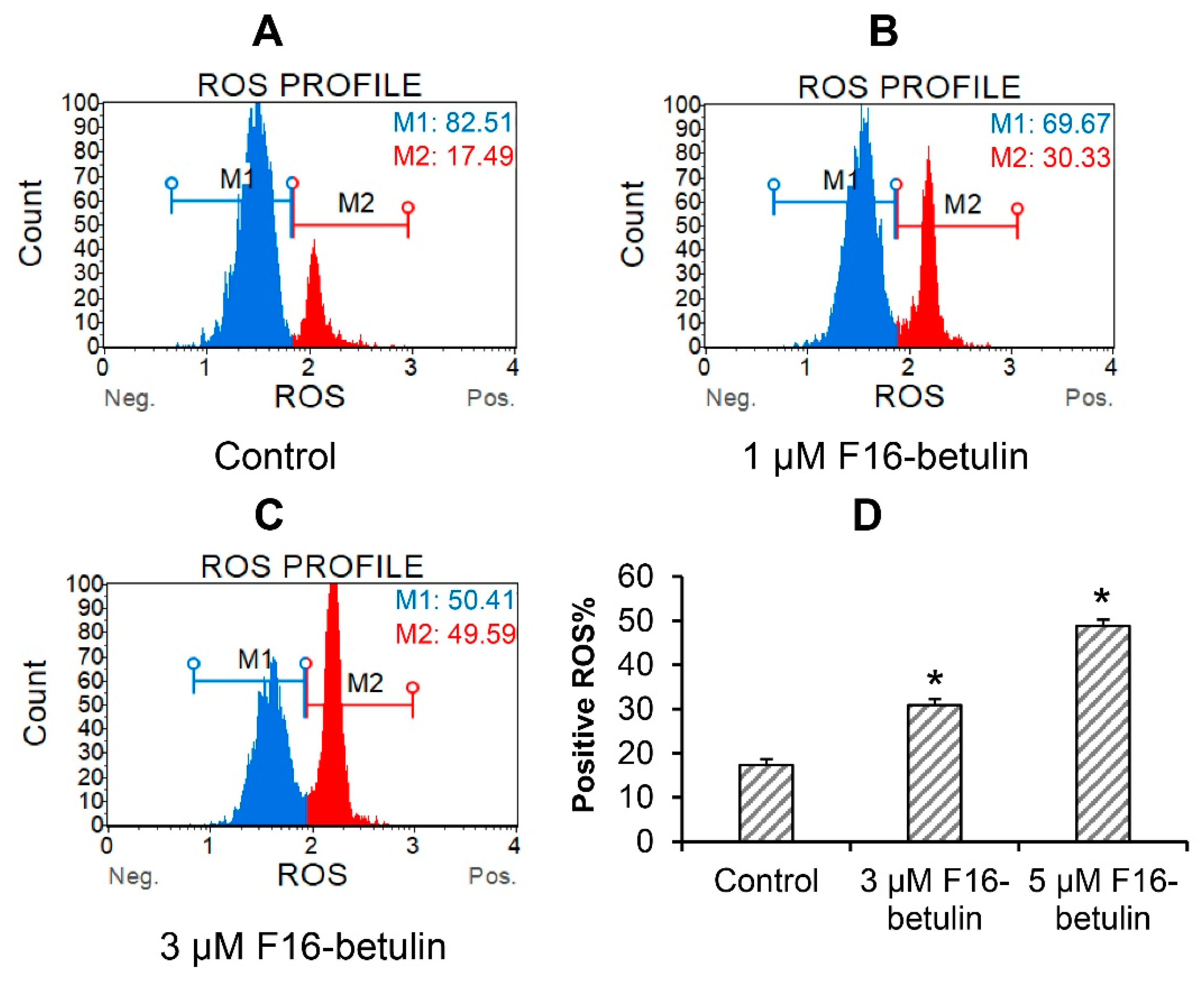
3.1. F16-Betulin Conjugate Induces Death of Rat Thymocytes due to Superoxide Overproduction and Mitochondrial Dysfunction
3.2. F16-Betulin Conjugate Affects the Surface Properties of Mitochondrial Membranes and Induces Their Aggregation
3.3. F16-Betulin Conjugate Induces Mitochondrial Dysfunction
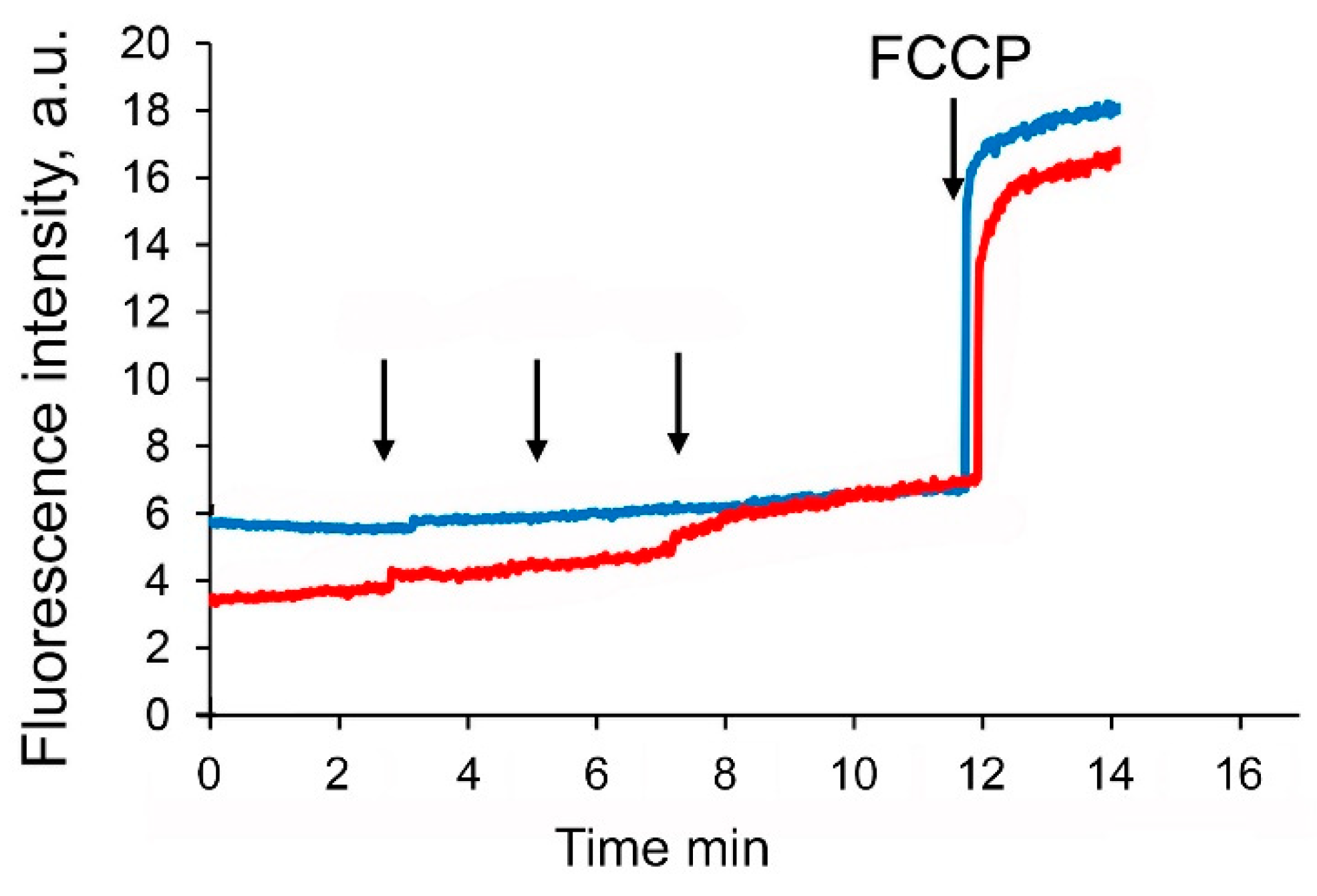
3.4. F16-Betulin Conjugate Induces Overproduction of Hydrogen Peroxide by Mitochondria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Connolly, J.D.; Hill, R.A. Triterpenoids. Nat. Prod. Rep. 2010, 27, 79–132. [Google Scholar] [CrossRef] [PubMed]
- Spivak, A.Y.; Nedopekina, D.A.; Khalitova, R.R.; Gubaidullin, R.R.; Odinokov, V.N.; Bel’skii, Y.P.; Bel’skaya, N.V.; Khazanov, V.A. Triphenylphosphonium cations of betulinic acid derivatives: Synthesis and antitumor activity. Med. Chem. Res. 2017, 26, 518–531. [Google Scholar] [CrossRef]
- Spivak, A.Y.; Nedopekina, D.A.; Shakurova, E.R.; Khalitova, R.R.; Gubaidullin, R.R.; Odinokov, V.N.; Dzhemilev, U.M.; Bel’skii, Y.P.; Bel’skaya, N.V.; Stankevich, S.A.; et al. Synthesis of lupane triterpenoids with triphenylphosphonium substituents and studies of their antitumor activity. Russ. Chem. Bull. 2013, 62, 188–198. [Google Scholar] [CrossRef]
- Nedopekina, D.A.; Gubaidullin, R.R.; Odinokov, V.N.; Maximchik, P.V.; Zhivotovsky, B.; Bel’skii, Y.P.; Khazanov, V.A.; Manuylova, A.V.; Gogvadze, V.; Spivak, A.Y. Mitochondria-targeted betulinic and ursolic acid derivatives: Synthesis and anticancer activity. MedChemComm 2017, 8, 1934–1945. [Google Scholar] [CrossRef]
- Ye, Y.; Zhang, T.; Yuan, H.; Li, D.; Lou, H.; Fan, P. Mitochondria- targeted lupane triterpenoid derivatives and their selective apoptosis-inducing anticancer mechanisms. J. Med. Chem. 2017, 60, 6353–6363. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Penkov, N.V.; Nedopekina, D.A.; Sharapov, V.A.; Khoroshavina, E.I.; Davletshin, E.V.; Belosludtseva, N.V.; Spivak, A.Y.; et al. Mitochondria-targeted prooxidant effects of betulinic acid conjugated with delocalized lipophilic cation F16. Free Radic. Biol. Med. 2021, 168, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Protasoni, M.; Zeviani, M. Mitochondrial structure and bioenergetics in normal and disease conditions. Int. J. Mol. Sci. 2021, 22, 586. [Google Scholar] [CrossRef] [PubMed]
- Belosludtsev, K.N.; Dubinin, M.V.; Belosludtseva, N.V.; Mironova, G.D. Mitochondrial Ca2+ transport: Mechanisms, molecular structures, and role in cells. Biochemistry 2019, 84, 593–597. [Google Scholar] [CrossRef]
- Wang, S.F.; Chen, S.; Tseng, L.M.; Lee, H.C. Role of the mitochondrial stress response in human cancer progression. Exp. Biol. Med. 2020, 245, 861–878. [Google Scholar] [CrossRef]
- Zhang, B.B.; Wang, D.G.; Guo, F.F.; Xuan, C. Mitochondrial membrane potential and reactive oxygen species in cancer stem cells. Fam. Cancer 2015, 14, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Khailova, L.S.; Silachev, D.N.; Rokitskaya, T.I.; Avetisyan, A.V.; Lyamsaev, K.G.; Severina, I.I.; Il’yasova, T.M.; Gulyaev, M.V.; Dedukhova, V.I.; Trendeleva, T.A.; et al. A short-chain alkyl derivative of rhodamine 19 acts as a mild uncoupler of mitochondria and a neuroprotector. Biochim. Biophys. Acta 2014, 1837, 1739–1747. [Google Scholar] [CrossRef]
- Antonenko, Y.N.; Denisov, S.S.; Silachev, D.N.; Khailova, L.S.; Jankauskas, S.S.; Rokitskaya, T.I.; Danilina, T.I.; Kotova, E.A.; Korshunova, G.A.; Plotnikov, E.Y.; et al. A long-linker conjugate of fluorescein and triphenylphosphonium as mitochondria-targeted uncoupler and fluorescent neuro- and nephroprotector. Biochim. Biophys. Acta 2016, 1860, 2463–2473. [Google Scholar] [CrossRef] [PubMed]
- Starenki, D.; Park, J.I. Selective mitochondrial uptake of MKT-077 can suppress medullary thyroid carcinoma cell survival in vitro and in vivo. Endocrinol. Metab. 2015, 30, 593. [Google Scholar] [CrossRef] [PubMed]
- Battogtokh, G.; Choi, Y.S.; Kang, D.S.; Park, S.J.; Shim, M.S.; Huh, K.M.; Cho, Y.Y.; Lee, J.Y.; Lee, H.S.; Kang, H.C. Mitochondria-targeting drug conjugates for cytotoxic, anti-oxidizing and sensing purposes: Current strategies and future perspectives. Acta Pharm. Sin. B 2018, 8, 862–880. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Tenkov, K.S.; Sharapov, V.A.; Kosareva, E.A.; Dubinin, M.V. Effect of dequalinium on respiration and the inner membrane permeability of rat liver mitochondria. Biochem. Moscow Suppl. Ser. A 2018, 12, 121–127. [Google Scholar] [CrossRef]
- Tate, A.D.; Antonelli, P.J.; Hannabass, K.R.; Dirain, C.O. Mitochondria-targeted antioxidant mitoquinone reduces cisplatin-induced ototoxicity in guinea pigs. Otolaryngol. Head Neck Surg. 2017, 156, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Cho, Y.Y.; Shim, M.S.; Lee, J.Y.; Lee, H.S.; Kang, H.C. Mitochondria-targeted drug delivery in cancers. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165808. [Google Scholar] [CrossRef]
- Iaubasarova, I.R.; Khailova, L.S.; Firsov, A.M.; Grivennikova, V.G.; Kirsanov, R.S.; Korshunova, G.A.; Kotova, E.A.; Antonenko, Y.N. The mitochondria-targeted derivative of the classical uncoupler of oxidative phosphorylation carbonyl cyanide m-chlorophenylhydrazone is an effective mitochondrial recoupler. PLoS ONE 2020, 15, e0244499. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, J.; Chen, Y. Betulinic acid and the pharmacological effects of tumor suppression. Mol. Med. Rep. 2016, 14, 4489–4495. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Green, I.R.; Ali, I.; Khan, I.A.; Ali, Z.; Al-Sadi, A.M.; Ahmed, I. Ursolic acid derivatives for pharmaceutical use: A patent review (2012–2016). Expert Opin. Ther. Pat. 2017, 27, 1061–1072. [Google Scholar] [CrossRef]
- Mullauer, F.B.; Kessler, J.H.; Medema, J.P. Betulinic acid, a natural compound with potent anti-cancer effects. Anticancer Drugs 2010, 21, 215–227. [Google Scholar] [CrossRef]
- Fantin, V.R.; Leder, P. F16, a mitochondriotoxic compound, triggers apoptosis or necrosis depending on the genetic background of the target carcinoma cell. Cancer Res. 2004, 64, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Spivak, A.Y.; Nedopekina, D.A.; Gubaidullin, R.R.; Davletshin, E.V.; Tukhbatullin, A.A.; D’yakonov, V.A.; Yunusbaeva, M.M.; Dzhemileva, L.U.; Dzhemilev, U.M. Pentacyclic triterpene acid conjugated with mitochondria-targeting cation F16: Synthesis and evaluation of cytotoxic activities. Med. Chem. Res. 2021, 30, 940–951. [Google Scholar] [CrossRef]
- Tsepaeva, O.V.; Nemtarev, A.V.; Abdullin, T.I.; Grigor’eva, L.R.; Kuznetsova, E.V.; Akhmadishina, R.A.; Ziganshina, L.E.; Cong, H.H.; Mironov, V.F. Design, synthesis, and cancer cell growth inhibitory activity of triphenylphosphonium derivatives of the triterpenoid botulin. J. Nat. Prod. 2017, 80, 2232–2239. [Google Scholar] [CrossRef]
- He, H.; Li, D.W.; Yang, L.Y.; Fu, L.; Zhu, X.J.; Wong, W.K.; Jiang, F.L.; Liu, Y. A novel bifunctional mitochondria-targeted anticancer agent with high selectivity for cancer cells. Sci. Rep. 2015, 5, 13543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Wang, L.Y.; Zhai, G.H.; Wen, Z.Y.; Zhang, Z.X. Microwave-assisted solvent-free synthesis of some dimethine cyanine dyes, spectral properties and TD-DFT/PCM calculations. Bull. Korean Chem. Soc. 2007, 28, 2382–2388. [Google Scholar] [CrossRef]
- Guo, L.; Chan, M.S.; Xu, D.; Tam, D.Y.; Bolze, F.; Lo, P.K.; Wong, M.S. Indole-based cyanine as a nuclear RNA-selective two-photon fluorescent probe for live cell imaging. ACS Chem. Biol. 2015, 10, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Konoshenko, G.I.; Mokhova, E.N. Characterization of thymocyte mitochondria. Biokhimiya 1983, 48, 652–657. [Google Scholar]
- Vedernikov, A.A.; Dubinin, M.V.; Zabiakin, V.A.; Samartsev, V.N. Ca(2+)-dependent nonspecific permeability of the inner membrane of liver mitochondria in the guinea fowl (Numida meleagris). J. Bioenergy Biomembr. 2015, 47, 235–242. [Google Scholar] [CrossRef]
- Belosludtsev, K.N.; Belosludtseva, N.V.; Kosareva, E.A.; Talanov, E.Y.; Gudkov, S.V.; Dubinin, M.V. Itaconic acid impairs the mitochondrial function by the inhibition of complexes II and IV and induction of the permeability transition pore opening in rat liver mitochondria. Biochimie 2020, 176, 150–157. [Google Scholar] [CrossRef]
- Spinazzi, M.; Casarin, A.; Pertegato, V.; Salviati, L.; Angelini, C. Assessment of mitochondrial respiratory chain enzymatic activities on tissues and cultured cells. Nat. Protoc. 2012, 7, 1235–1246. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Svinin, A.O.; Vedernikov, A.A.; Starinets, V.S.; Tenkov, K.S.; Belosludtsev, K.N.; Samartsev, V.N. Effect of hypothermia on the functional activity of liver mitochondria of grass snake (Natrix natrix): Inhibition of succinate-fueled respiration and K+ transport, ROS-induced activation of mitochondrial permeability transition. J. Bioenergy Biomembr. 2019, 51, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Król, S.K.; Kiełbus, M.; Rivero-Müller, A.; Stepulak, A. Comprehensive review on betulin as a potent anticancer agent. Biomed. Res. Int. 2015, 2015, 584189. [Google Scholar] [CrossRef]
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Mikheeva, I.B.; Yashin, V.A.; Penkov, N.V.; Vydrina, V.A.; Ishmuratov, G.Y.; Sharapov, V.A.; Khoroshavina, E.I.; et al. Effect of betulin and betulonic acid on isolated rat liver mitochondria and liposomes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183383. [Google Scholar] [CrossRef]
- Lemeshko, V.V.; Lopez, L.F.; Solano, S.; Torres, R. The natural antioxidant otobaphenol delays the permeability transition of mitochondria and induces their aggregation. Antioxid. Redox Signal. 2003, 5, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, M.V.; Samartsev, V.N.; Stepanova, A.E.; Khoroshavina, E.I.; Penkov, N.V.; Yashin, V.A.; Starinets, V.S.; Mikheeva, I.B.; Gudkov, S.V.; Belosludtsev, K.N. Membranotropic effects of ω-hydroxypalmitic acid and Ca2+ on rat liver mitochondria and lecithin liposomes. Aggregation and membrane permeabilization. J. Bioenergy Biomembr. 2018, 50, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Bosetti, F.; Baracca, A.; Lenaz, G.; Solaini, G. Increased state 4 mitochondrial respiration and swelling in early post-ischemic reperfusion of rat heart. FEBS Lett. 2004, 563, 161–164. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R.; Singer, T.P. Relation of superoxide generation and lipid peroxidation to the inhibition of NADH-Q oxidoreductase by rotenone, piericidin A, and MPP. Biochem. Biophys. Res. Commun. 1992, 189, 47–52. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Additions | State 2 | State 3 | State 4 | State 3UDNP | RC |
|---|---|---|---|---|---|
| nmol O2/min/mg Protein | Relative Units | ||||
| Control | 3.82 ± 0.44 | 23.56 ± 0.84 | 4.37 ± 0.13 | 23.23 ± 2.41 | 5.40 ± 0.08 |
| F16-betulin conjugate | |||||
| 10 μM | 4.49 ± 0.43 | 25.80 ± 1.00 | 4.77 ± 0.15 | 27.72 ± 0.79 | 5.41 ± 0.04 |
| 20 μM + 1 µg/mL oligo | 4.64 ± 0.54 | 26.32 ± 0.58 | 5.34 ± 0.18 * 4.21 ± 0.22 # | 28.09 ± 1.39 | 4.93 ± 0.06 * |
| Additions | State 2 | State 3 | State 4 | State 3UDNP | RC |
|---|---|---|---|---|---|
| nmol O2/min/mg Protein | Relative Units | ||||
| Control | 9.86 ± 0.91 | 44.62 ± 2.30 | 9.14 ± 0.67 | 49.54 ± 2.96 | 4.91 ± 0.15 |
| F16-betulin conjugate | |||||
| 10 μM | 9.56 ± 1.00 | 44.91 ± 0.55 | 9.39 ± 1.27 | 51.16 ± 0.94 | 4.87 ± 0.44 |
| 20 μM | 9.85 ± 0.11 | 40.67 ± 0.83 | 9.25 ± 0.43 | 43.25 ± 0.51 | 4.41 ± 0.12 |
| Complex I | Complex II | Complex III | Complex IV | |
|---|---|---|---|---|
| 10 µM conjugate | 99.76 ± 3.74 | 89.51 ± 1.09 * | 112.97 ± 3.31 * | 103.49 ± 3.02 |
| 20 µM conjugate | 62.27 ± 1.76 * | 85.68 ± 3.04 * | 124.66 ± 1.37 * | 91.83 ± 3.77 |
| Glutamate/Malate | Succinate/Rotenone | |
|---|---|---|
| Control | 146 ± 6 | 164 ± 7 |
| 5 µM conjugate | 185 ± 8 * | 168 ± 2 |
| 10 µM conjugate | 205 ± 15 * | 164 ± 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dubinin, M.V.; Semenova, A.A.; Nedopekina, D.A.; Davletshin, E.V.; Spivak, A.Y.; Belosludtsev, K.N. Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation. Membranes 2021, 11, 352. https://doi.org/10.3390/membranes11050352
Dubinin MV, Semenova AA, Nedopekina DA, Davletshin EV, Spivak AY, Belosludtsev KN. Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation. Membranes. 2021; 11(5):352. https://doi.org/10.3390/membranes11050352
Chicago/Turabian StyleDubinin, Mikhail V., Alena A. Semenova, Darya A. Nedopekina, Eldar V. Davletshin, Anna Yu. Spivak, and Konstantin N. Belosludtsev. 2021. "Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation" Membranes 11, no. 5: 352. https://doi.org/10.3390/membranes11050352
APA StyleDubinin, M. V., Semenova, A. A., Nedopekina, D. A., Davletshin, E. V., Spivak, A. Y., & Belosludtsev, K. N. (2021). Effect of F16-Betulin Conjugate on Mitochondrial Membranes and Its Role in Cell Death Initiation. Membranes, 11(5), 352. https://doi.org/10.3390/membranes11050352