
The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthetic Liposome Preparation
2.2. Virus Growth and Purification
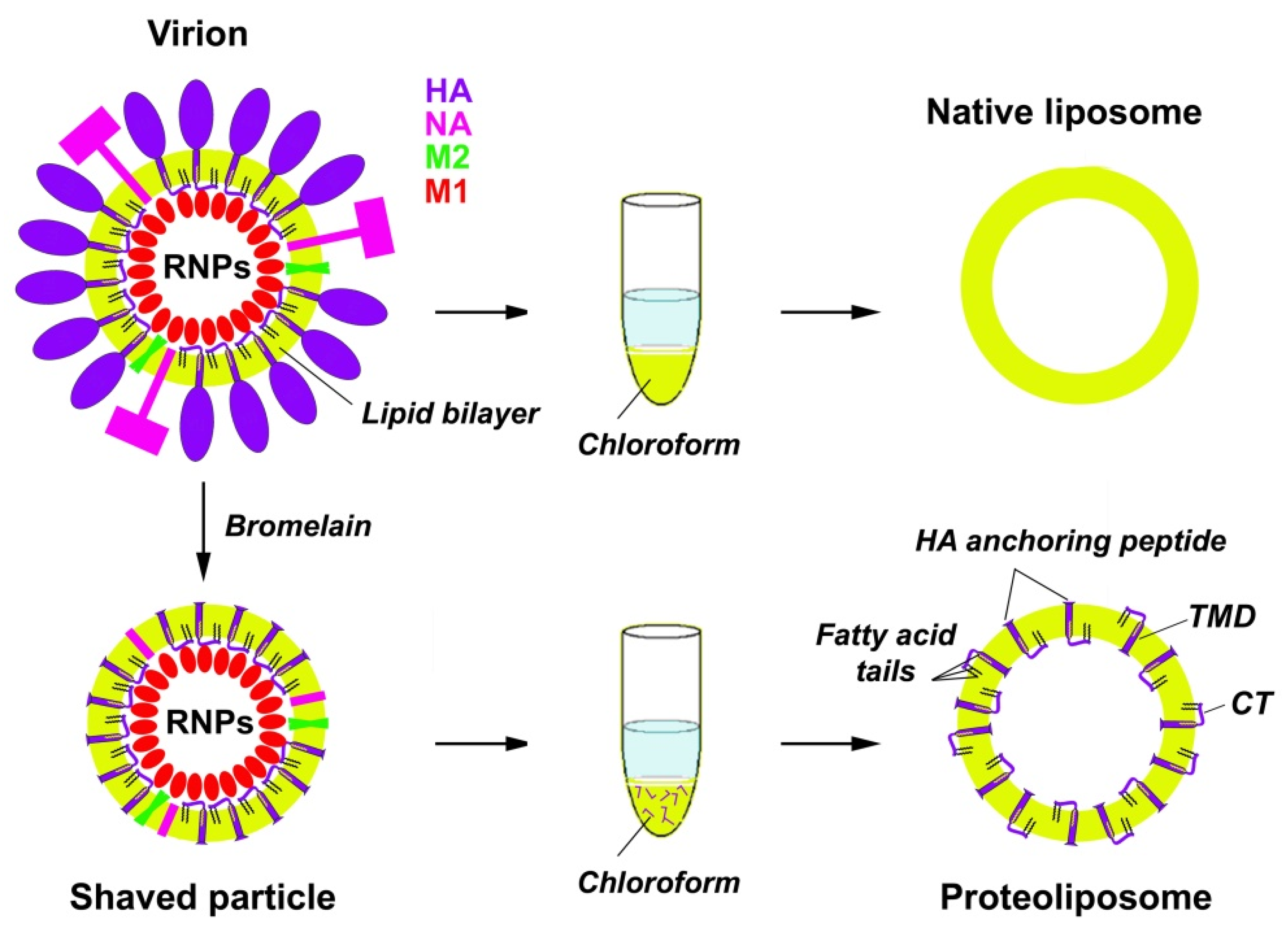
2.3. Preparation of the Subviral Particles
2.4. Preparation of Native Viral Liposomes and Proteoliposomes with HA LI45 Peptides
2.5. Isolation of M1 Protein
2.6. Dynamic Light Scattering (DLS) Analysis
2.7. Negative-Stain Transmission Electron Microscopy (EM)
2.8. Small-Angle X-ray Scattering
2.9. Modeling the Electron Density Distribution of the Phospholipid Bilayer
3. Results and Discussion
3.1. Synthetic Liposomes
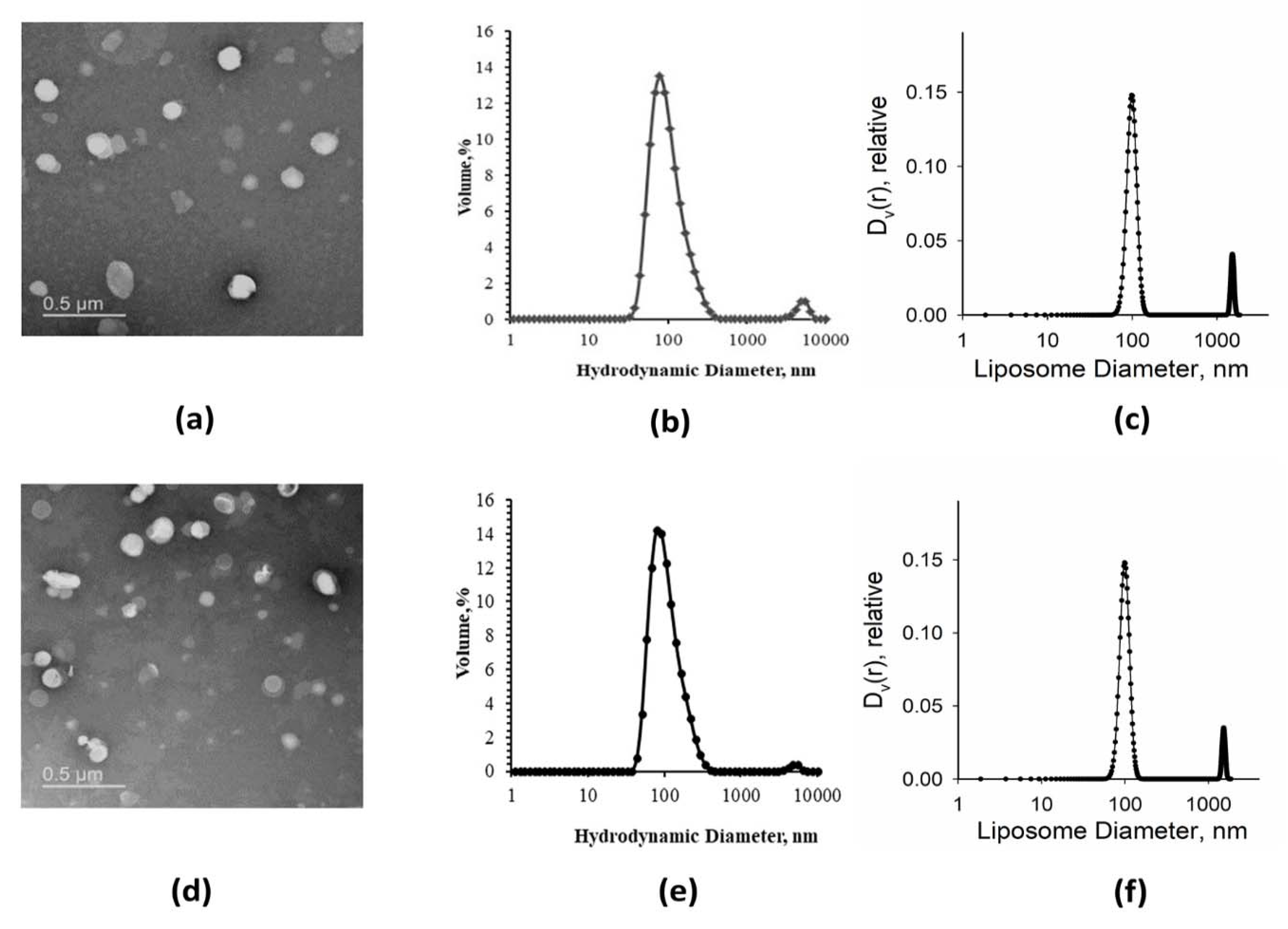
3.1.1. Electron Microscopy (EM), Dynamic Light Scattering (DLS), and Small-Angle X-ray Scattering (SAXS) Analysis of Size Distributions of Synthetic Liposomes
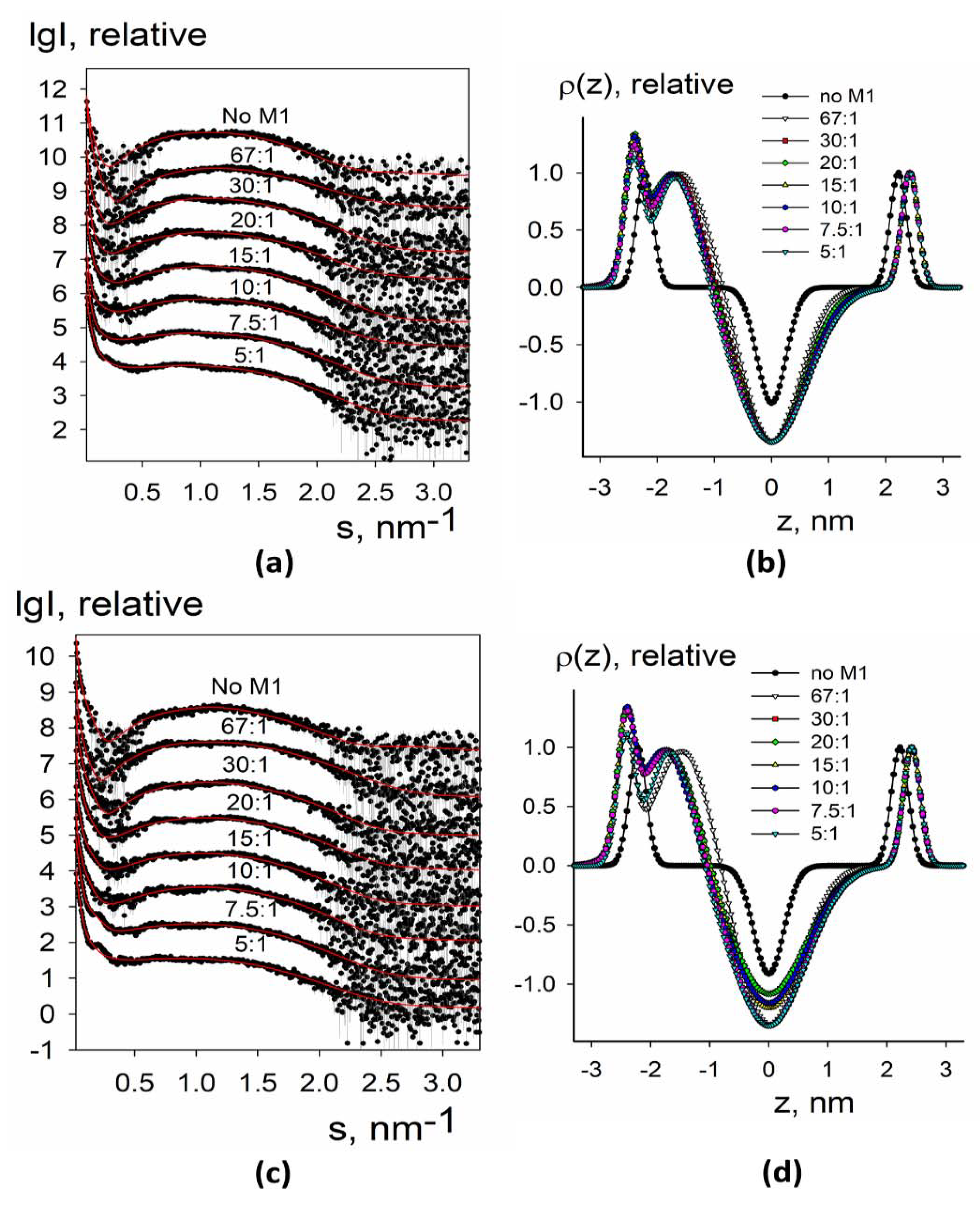
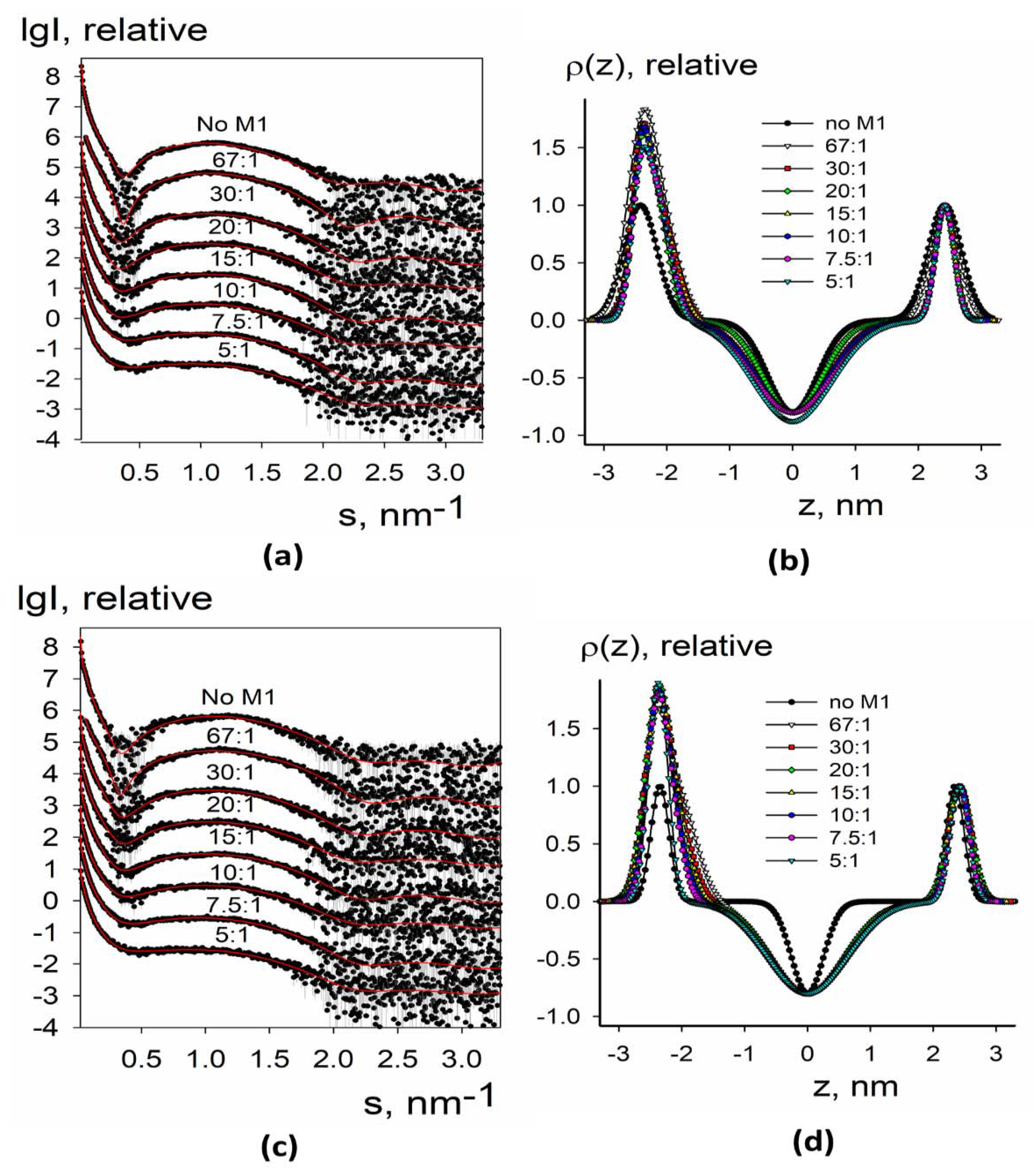
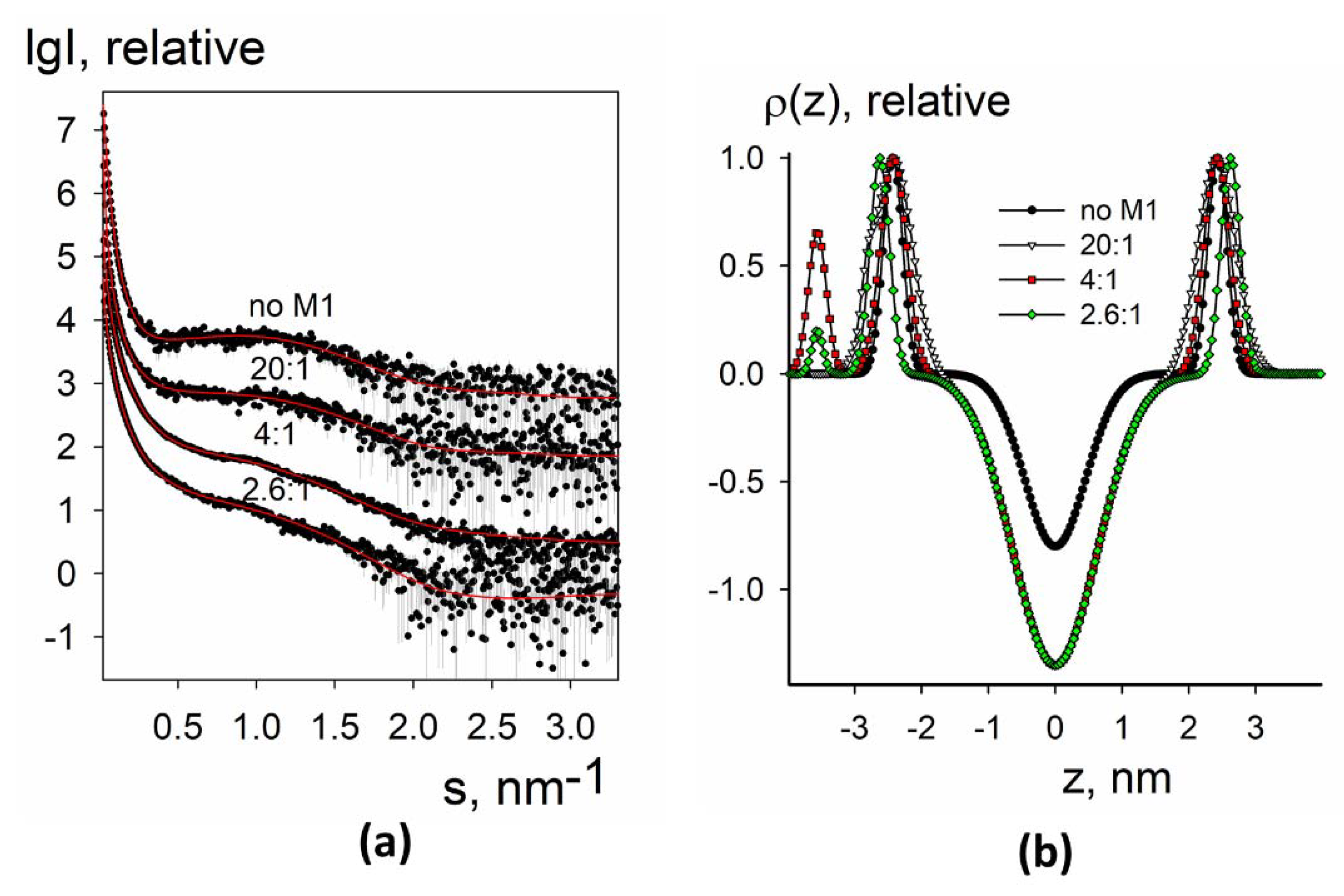
3.1.2. SAXS Analysis of the Electron Density Profiles of Synthetic Liposomes
3.2. Native Liposomes and Proteoliposomes
3.2.1. EM and DLS Analysis of Native Liposomes and Proteoliposomes
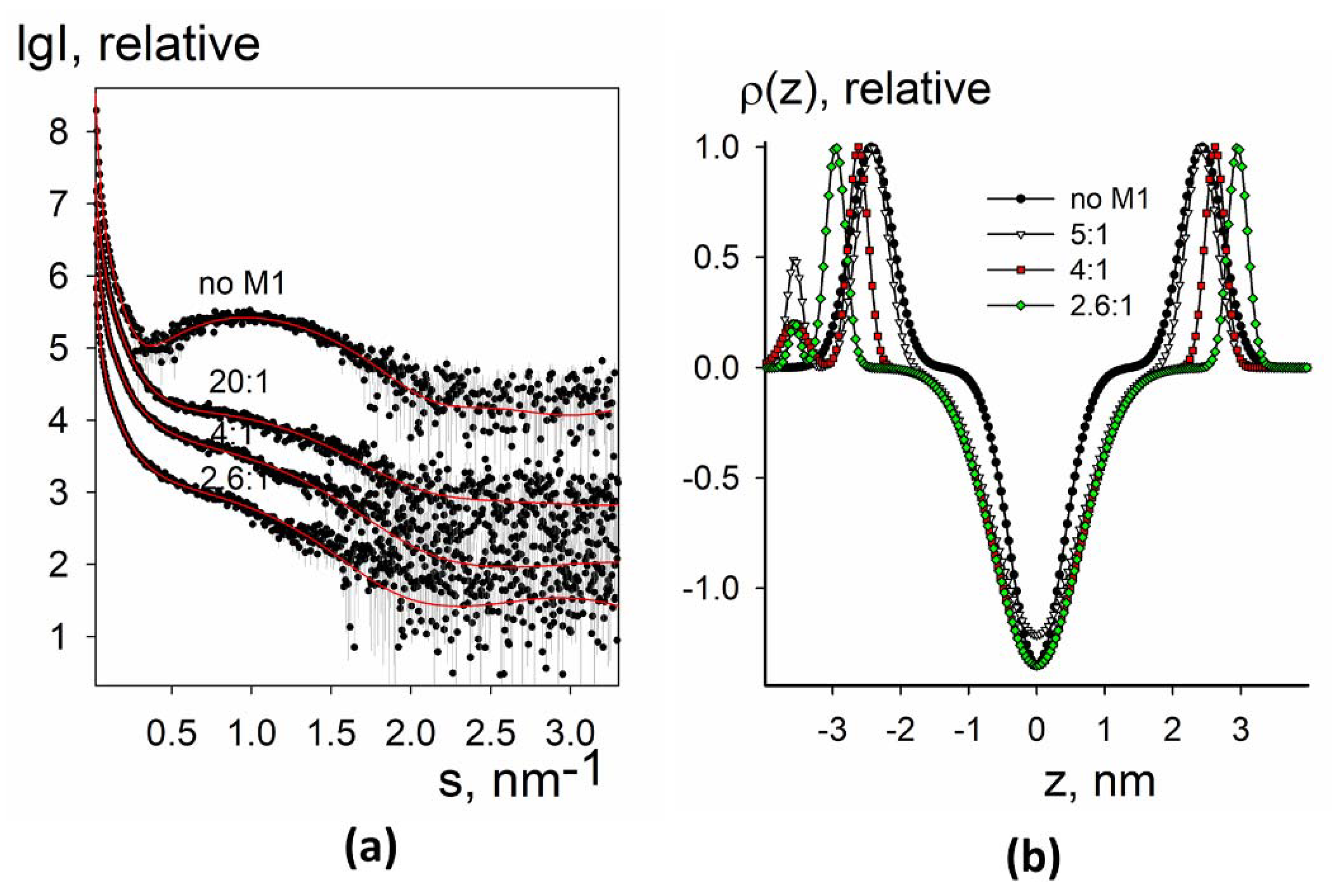
3.2.2. SAXS Analysis of Electron Density Profiles of Native Liposomes and Proteoliposomes
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rossman, J.S.; Lamb, R.A. Influenza Virus Assembly and Budding. Virology 2011, 411, 229–236. [Google Scholar] [CrossRef]
- Harris, A.; Forouhar, F.; Qiu, S.; Sha, B.; Luo, M. The Crystal Structure of the Influenza Matrix Protein M1 at Neutral PH: M1-M1 Protein Interfaces Can Rotate in the Oligomeric Structures of M1. Virology 2001, 289, 34–44. [Google Scholar] [CrossRef]
- Shtykova, E.V.; Dadinova, L.A.; Fedorova, N.V.; Golanikov, A.E.; Bogacheva, E.N.; Ksenofontov, A.L.; Baratova, L.A.; Shilova, L.A.; Tashkin, V.Y.; Galimzyanov, T.R.; et al. Influenza Virus Matrix Protein M1 Preserves Its Conformation with PH, Changing Multimerization State at the Priming Stage Due to Electrostatics. Sci. Rep. 2017, 7, 16793. [Google Scholar] [CrossRef]
- Batishchev, O.V.; Shilova, L.A.; Kachala, M.V.; Tashkin, V.Y.; Sokolov, V.S.; Fedorova, N.V.; Baratova, L.A.; Knyazev, D.G.; Zimmerberg, J.; Chizmadzhev, Y.A. PH-Dependent Formation and Disintegration of the Influenza A Virus Protein Scaffold To Provide Tension for Membrane Fusion. J. Virol. 2016, 90, 575–585. [Google Scholar] [CrossRef]
- McCauley, J.W.; Mahy, B.W. Structure and Function of the Influenza Virus Genome. Biochem. J. 1983, 211, 281–294. [Google Scholar] [CrossRef]
- Sha, B.; Luo, M. Structure of a Bifunctional Membrane-RNA Binding Protein, Influenza Virus Matrix Protein M1. Nat. Struct. Biol. 1997, 4, 239–244. [Google Scholar] [CrossRef]
- Arzt, S.; Baudin, F.; Barge, A.; Timmins, P.; Burmeister, W.P.; Ruigrok, R.W. Combined Results from Solution Studies on Intact Influenza Virus M1 Protein and from a New Crystal Form of Its N-Terminal Domain Show That M1 Is an Elongated Monomer. Virology 2001, 279, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Safo, M.K.; Musayev, F.N.; Mosier, P.D.; Zhou, Q.; Xie, H.; Desai, U.R. Crystal Structures of Influenza A Virus Matrix Protein M1: Variations on a Theme. PLoS ONE 2014, 9, e109510. [Google Scholar] [CrossRef] [PubMed]
- Shtykova, E.V.; Baratova, L.A.; Fedorova, N.V.; Radyukhin, V.A.; Ksenofontov, A.L.; Volkov, V.V.; Shishkov, A.V.; Dolgov, A.A.; Shilova, L.A.; Batishchev, O.V.; et al. Structural Analysis of Influenza A Virus Matrix Protein M1 and Its Self-Assemblies at Low PH. PLoS ONE 2013, 8, e82431. [Google Scholar] [CrossRef] [PubMed]
- Gregoriades, A.; Frangione, B. Insertion of Influenza M Protein into the Viral Lipid Bilayer and Localization of Site of Insertion. J. Virol. 1981, 40, 323–328. [Google Scholar] [CrossRef]
- Ruigrok, R.W.H.; Barge, A.; Durrer, P.; Brunner, J.; Ma, K.; Whittaker, G.R. Membrane Interaction of Influenza Virus M1 Protein. Virology 2000, 267, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Bobone, S.; Hilsch, M.; Storm, J.; Dunsing, V.; Herrmann, A.; Chiantia, S. Phosphatidylserine Lateral Organization Influences the Interaction of Influenza Virus Matrix Protein 1 with Lipid Membranes. J. Virol. 2017, 91, e00267-17. [Google Scholar] [CrossRef] [PubMed]
- Hilsch, M.; Goldenbogen, B.; Sieben, C.; Höfer, C.T.; Rabe, J.P.; Klipp, E.; Herrmann, A.; Chiantia, S. Influenza A Matrix Protein M1 Multimerizes upon Binding to Lipid Membranes. Biophys. J. 2014, 107, 912–923. [Google Scholar] [CrossRef]
- Peukes, J.; Xiong, X.; Erlendsson, S.; Qu, K.; Wan, W.; Calder, L.J.; Schraidt, O.; Kummer, S.; Freund, S.M.V.; Kräusslich, H.-G.; et al. The Native Structure of the Assembled Matrix Protein 1 of Influenza A Virus. Nature 2020, 587, 495–498. [Google Scholar] [CrossRef]
- Leser, G.P.; Lamb, R.A. Lateral Organization of Influenza Virus Proteins in the Budozone Region of the Plasma Membrane. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Ali, A.; Avalos, R.T.; Ponimaskin, E.; Nayak, D.P. Influenza Virus Assembly: Effect of Influenza Virus Glycoproteins on the Membrane Association of M1 Protein. J. Virol. 2000, 74, 8709–8719. [Google Scholar] [CrossRef]
- Zhang, J.; Leser, G.P.; Pekosz, A.; Lamb, R.A. The Cytoplasmic Tails of the Influenza Virus Spike Glycoproteins Are Required for Normal Genome Packaging. Virology 2000, 269, 325–334. [Google Scholar] [CrossRef]
- Jin, H.; Leser, G.P.; Zhang, J.; Lamb, R.A. Influenza Virus Hemagglutinin and Neuraminidase Cytoplasmic Tails Control Particle Shape. EMBO J. 1997, 16, 1236–1247. [Google Scholar] [CrossRef]
- Kordyukova, L.V.; Shtykova, E.V.; Baratova, L.A.; Svergun, D.I.; Batishchev, O.V. Matrix Proteins of Enveloped Viruses: A Case Study of Influenza A Virus M1 Protein. J. Biomol. Struct. Dyn. 2019, 37, 671–690. [Google Scholar] [CrossRef] [PubMed]
- Petrich, A.; Dunsing, V.; Bobone, S.; Chiantia, S. Influenza A M2 Recruits M1 to the Plasma Membrane: A Fluorescence Fluctuation Microscopy Study. BioRxiv. 2021. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Li, S.; Eghiaian, F.; Sieben, C.; Herrmann, A.; Schaap, I.A.T. Bending and Puncturing the Influenza Lipid Envelope. Biophys. J. 2011, 100, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Serebryakova, M.V.; Kordyukova, L.V.; Baratova, L.A.; Markushin, S.G. Mass Spectrometric Sequencing and Acylation Character Analysis of C-Terminal Anchoring Segment from Influenza A Hemagglutinin. Eur. J. Mass Spectrom. 2006, 12, 51–62. [Google Scholar] [CrossRef]
- Kordyukova, L.V.; Serebryakova, M.V.; Baratova, L.A.; Veit, M. S Acylation of the Hemagglutinin of Influenza Viruses: Mass Spectrometry Reveals Site-Specific Attachment of Stearic Acid to a Transmembrane Cysteine. J. Virol. 2008, 82, 9288–9292. [Google Scholar] [CrossRef] [PubMed]
- Serebryakova, M.V.; Kordyukova, L.V.; Semashko, T.A.; Ksenofontov, A.L.; Rudneva, I.A.; Kropotkina, E.A.; Filippova, I.Y.; Veit, M.; Baratova, L.A. Influenza Virus Hemagglutinin Spike Neck Architectures and Interaction with Model Enzymes Evaluated by MALDI-TOF Mass Spectrometry and Bioinformatics Tools. Virus Res. 2011, 160, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Peterson, G.L. A Simplification of the Protein Assay Method of Lowry et al. Which Is More Generally Applicable. Anal. Biochem. 1977, 83, 346–356. [Google Scholar] [CrossRef]
- Kordyukova, L.V.; Ksenofontov, A.L.; Serebryakova, M.V.; Ovchinnikova, T.V.; Fedorova, N.V.; Ivanova, V.T.; Baratova, L.A. Influenza A Hemagglutinin C-Terminal Anchoring Peptide: Identification and Mass Spectrometric Study. Protein Pept. Lett. 2004, 11, 385–391. [Google Scholar] [CrossRef]
- Zhirnov, O.P. Isolation of Matrix Protein M1 from Influenza Viruses by Acid-Dependent Extraction with Nonionic Detergent. Virology 1992, 186, 324–330. [Google Scholar] [CrossRef]
- Kordyukova, L.; Serebryakova, M.; Polyakov, V.; Ovchinnikova, T.; Smirnova, Y.; Fedorova, N.; Baratova, L. Influenza A Virus M1 Protein Structure Probed by In Situ Limited Proteolysis with Bromelain. PPL 2008, 15, 922–930. [Google Scholar] [CrossRef]
- Blanchet, C.E.; Spilotros, A.; Schwemmer, F.; Graewert, M.A.; Kikhney, A.; Jeffries, C.M.; Franke, D.; Mark, D.; Zengerle, R.; Cipriani, F.; et al. Versatile Sample Environments and Automation for Biological Solution X-Ray Scattering Experiments at the P12 Beamline (PETRA III, DESY). J. Appl. Crystallogr. 2015, 48, 431–443. [Google Scholar] [CrossRef]
- Konarev, P.V.; Volkov, V.V.; Sokolova, A.V.; Koch, M.H.J.; Svergun, D.I. PRIMUS: A Windows PC-Based System for Small-Angle Scattering Data Analysis. J. Appl. Crystallogr. 2003, 36, 1277–1282. [Google Scholar] [CrossRef]
- Franke, D.; Petoukhov, M.V.; Konarev, P.V.; Panjkovich, A.; Tuukkanen, A.; Mertens, H.D.T.; Kikhney, A.G.; Hajizadeh, N.R.; Franklin, J.M.; Jeffries, C.M.; et al. ATSAS 2.8: A Comprehensive Data Analysis Suite for Small-Angle Scattering from Macromolecular Solutions. J. Appl. Crystallogr. 2017, 50, 1212–1225. [Google Scholar] [CrossRef]
- Konarev, P.V.; Petoukhov, M.V.; Dadinova, L.A.; Fedorova, N.V.; Volynsky, P.E.; Svergun, D.I.; Batishchev, O.V.; Shtykova, E.V. BILMIX: A New Approach to Restore the Size Polydispersity and Electron Density Profiles of Lipid Bilayers from Liposomes Using Small-Angle X-Ray Scattering Data. J. Appl. Crystallogr. 2020, 53, 236–243. [Google Scholar] [CrossRef]
- MacDonald, R.C.; MacDonald, R.I.; Menco, B.P.; Takeshita, K.; Subbarao, N.K.; Hu, L.R. Small-Volume Extrusion Apparatus for Preparation of Large, Unilamellar Vesicles. Biochim. Biophys. Acta 1991, 1061, 297–303. [Google Scholar] [CrossRef]
- Frisken, B.J.; Asman, C.; Patty, P.J. Studies of Vesicle Extrusion. Langmuir 2000, 16, 928–933. [Google Scholar] [CrossRef]
- Konarev, P.V.; Gruzinov, A.Y.; Mertens, H.D.T.; Svergun, D.I. Restoring Structural Parameters of Lipid Mixtures from Small-Angle X-Ray Scattering Data. J. Appl. Crystallogr. 2021, 54, 169–179. [Google Scholar] [CrossRef]
- Kučerka, N.; Nieh, M.-P.; Katsaras, J. Small-Angle Scattering from Homogenous and Heterogeneous Lipid Bilayers. Advances in Planar Lipid Bilayers and Liposomes. 2010, 12, 201–235. [Google Scholar] [CrossRef]
- Kiselev, M.A.; Lesieur, P.; Kisselev, A.M.; Lombardo, D.; Aksenov, V.L. Model of Separated Form Factors for Unilamellar Vesicles. Appl. Phys. A: Mater. Sci. Process. 2002, 74, s1654–s1656. [Google Scholar] [CrossRef]
- Gennis, R.B. Biomembranes: Molecular Structure and Function; Springer: New York, NY, USA, 1989. [Google Scholar] [CrossRef]
- Saletti, D.; Radzimanowski, J.; Effantin, G.; Midtvedt, D.; Mangenot, S.; Weissenhorn, W.; Bassereau, P.; Bally, M. The Matrix Protein M1 from Influenza C Virus Induces Tubular Membrane Invaginations in an in Vitro Cell Membrane Model. Sci. Rep. 2017, 7, 40801. [Google Scholar] [CrossRef]
- Galimzyanov, T.R.; Lyushnyak, A.S.; Aleksandrova, V.V.; Shilova, L.A.; Mikhalyov, I.I.; Molotkovskaya, I.M.; Akimov, S.A.; Batishchev, O.V. Line Activity of Ganglioside GM1 Regulates the Raft Size Distribution in a Cholesterol-Dependent Manner. Langmuir 2017, 33, 3517–3524. [Google Scholar] [CrossRef]
- Hamm, M.; Kozlov, M.M. Elastic Energy of Tilt and Bending of Fluid Membranes. Eur. Phys. J. E 2000, 3, 323–335. [Google Scholar] [CrossRef]
- Khelashvili, G.; Kollmitzer, B.; Heftberger, P.; Pabst, G.; Harries, D. Calculating the Bending Modulus for Multicomponent Lipid Membranes in Different Thermodynamic Phases. J. Chem. Theory Comput. 2013, 9, 3866–3871. [Google Scholar] [CrossRef]
- Siche, S.; Brett, K.; Möller, L.; Kordyukova, L.; Mintaev, R.; Alexeevski, A.; Veit, M. Two Cytoplasmic Acylation Sites and an Adjacent Hydrophobic Residue, but No Other Conserved Amino Acids in the Cytoplasmic Tail of HA from Influenza A Virus Are Crucial for Virus Replication. Viruses 2015, 7, 6458–6475. [Google Scholar] [CrossRef] [PubMed]
- Veit, M.; Thaa, B. Association of Influenza Virus Proteins with Membrane Rafts. Adv. Virol. 2011, 2011, 370606. [Google Scholar] [CrossRef]
- Fontana, J.; Steven, A.C. At Low PH, Influenza Virus Matrix Protein M1 Undergoes a Conformational Change Prior to Dissociating from the Membrane. J. Virol. 2013, 87, 5621–5628. [Google Scholar] [CrossRef] [PubMed]
- Khrustalev, V.V.; Kordyukova, L.V.; Arutyunyan, A.M.; Poboinev, V.V.; Khrustaleva, T.A.; Stojarov, A.N.; Baratova, L.A.; Sapon, A.S.; Lugin, V.G. The Cytoplasmic Tail of Influenza A/H1N1 Virus Hemagglutinin Is β-Structural. J. Biomol. Struct. Dyn. 2020, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kikhney, A.G.; Borges, C.R.; Molodenskiy, D.S.; Jeffries, C.M.; Svergun, D.I. SASBDB: Towards an Automatically Curated and Validated Repository for Biological Scattering Data. Protein Sci. 2020, 29, 66–75. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kordyukova, L.V.; Konarev, P.V.; Fedorova, N.V.; Shtykova, E.V.; Ksenofontov, A.L.; Loshkarev, N.A.; Dadinova, L.A.; Timofeeva, T.A.; Abramchuk, S.S.; Moiseenko, A.V.; et al. The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer. Membranes 2021, 11, 772. https://doi.org/10.3390/membranes11100772
Kordyukova LV, Konarev PV, Fedorova NV, Shtykova EV, Ksenofontov AL, Loshkarev NA, Dadinova LA, Timofeeva TA, Abramchuk SS, Moiseenko AV, et al. The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer. Membranes. 2021; 11(10):772. https://doi.org/10.3390/membranes11100772
Chicago/Turabian StyleKordyukova, Larisa V., Petr V. Konarev, Nataliya V. Fedorova, Eleonora V. Shtykova, Alexander L. Ksenofontov, Nikita A. Loshkarev, Lubov A. Dadinova, Tatyana A. Timofeeva, Sergei S. Abramchuk, Andrei V. Moiseenko, and et al. 2021. "The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer" Membranes 11, no. 10: 772. https://doi.org/10.3390/membranes11100772
APA StyleKordyukova, L. V., Konarev, P. V., Fedorova, N. V., Shtykova, E. V., Ksenofontov, A. L., Loshkarev, N. A., Dadinova, L. A., Timofeeva, T. A., Abramchuk, S. S., Moiseenko, A. V., Baratova, L. A., Svergun, D. I., & Batishchev, O. V. (2021). The Cytoplasmic Tail of Influenza A Virus Hemagglutinin and Membrane Lipid Composition Change the Mode of M1 Protein Association with the Lipid Bilayer. Membranes, 11(10), 772. https://doi.org/10.3390/membranes11100772