
In Vitro Priming of Human T Cells by Dendritic Cells Provides a Screening Tool for Candidate Vaccines for Burkholderia pseudomallei

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Bacterial Preparation
2.2. Healthy Donors
2.3. Dendritic Cell Enrichment
2.4. Bacterial Stimulation
2.5. Cell-Surface Antibody Labelling
2.6. Intracellular Cytokine Analysis
2.7. Enzyme Linked Immunosorbent Assay (ELISA)
2.8. DC: T Cell Co-Cultures
2.9. Flow Cytometry Analysis
2.10. Statistical Analysis
3. Results
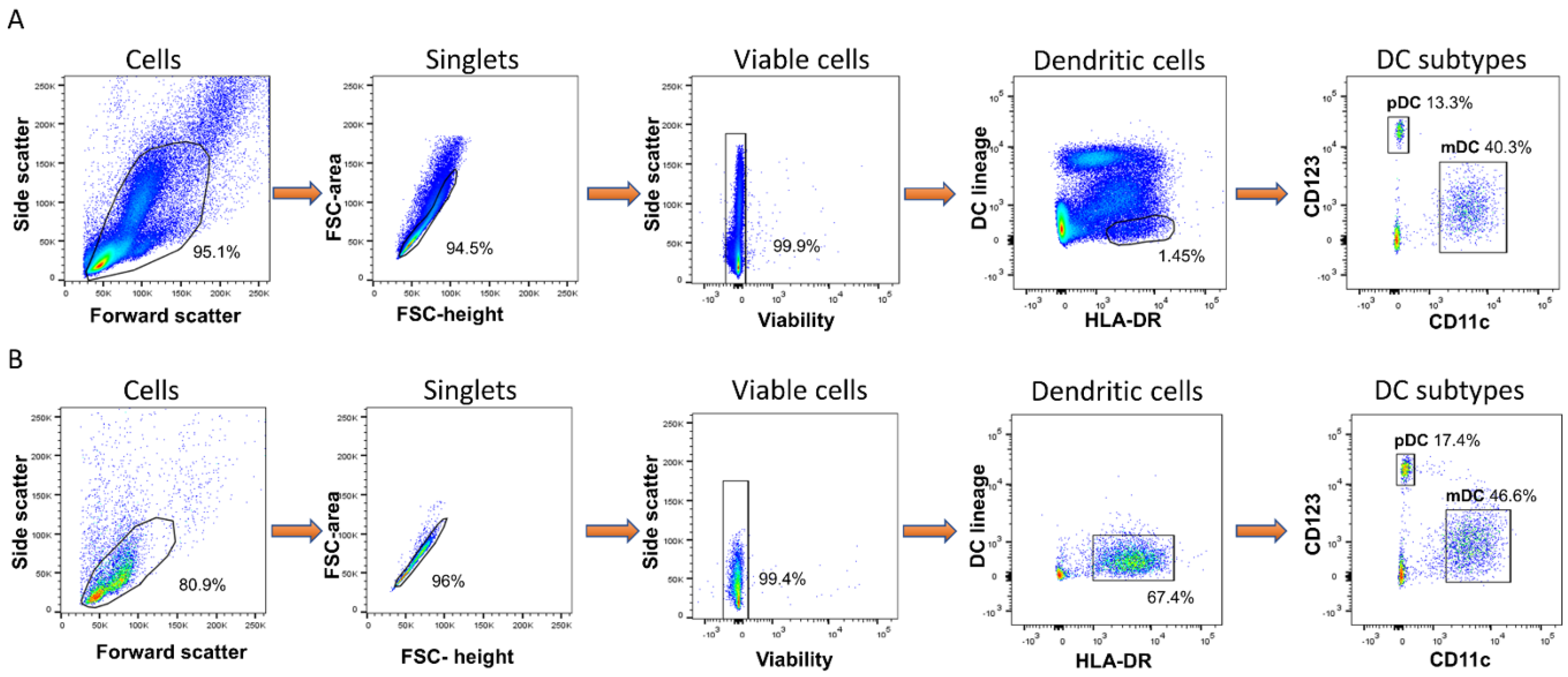
3.1. B. pseudomallei Activates Both mDCs and pDCs Enriched from Fresh Blood
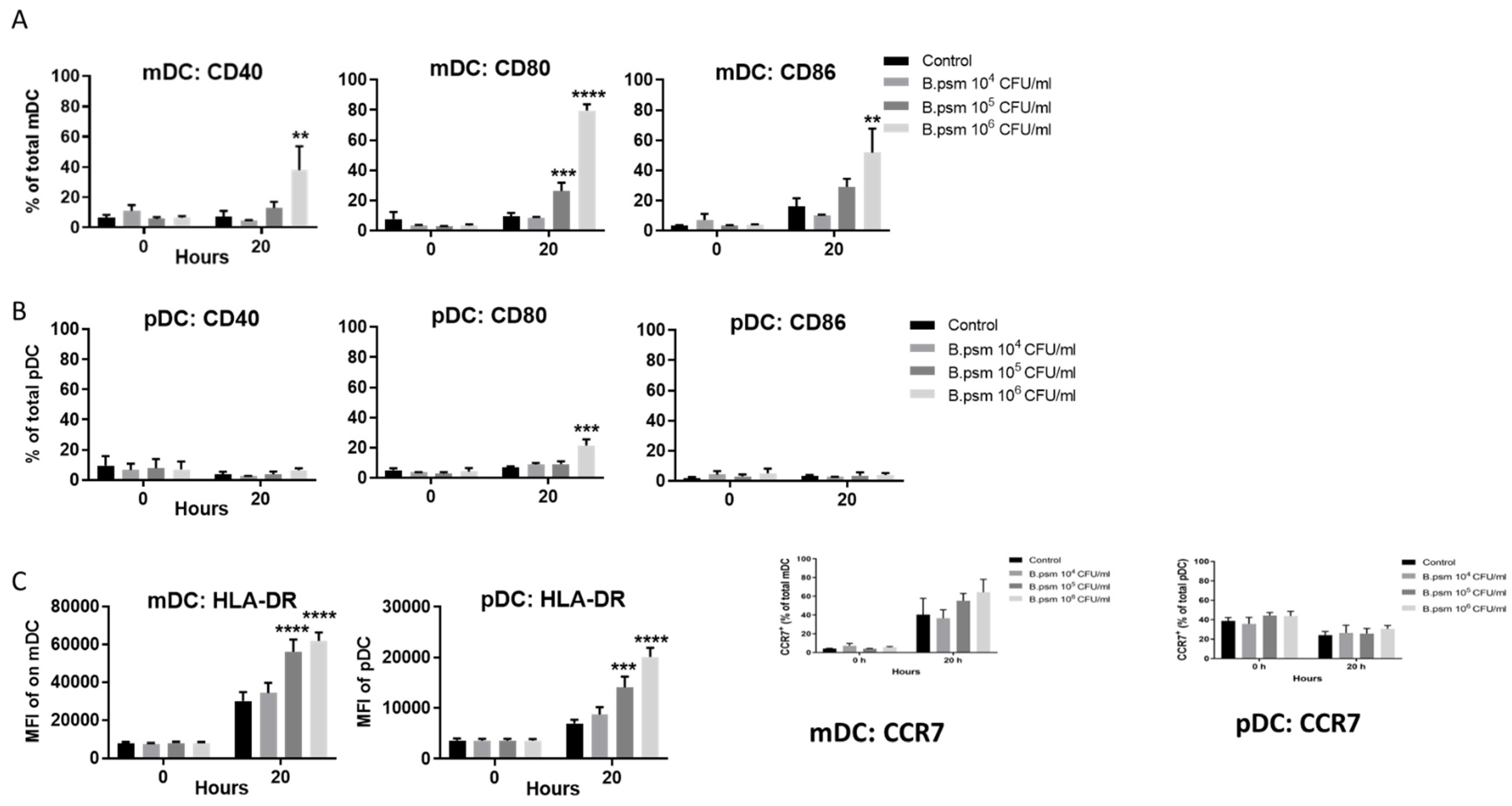
3.2. B.psm Stimulates Proinflammatory DC Cytokine Production
3.3. DCs Conditioned with B.psm Stimulate CD4+ T-Cell Proliferation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Author Summary
Content Includes Material Subject to © Crown Copyright (2021), Dstl
References
- Cheng, A.; Currie, B. Melioidosis: Epidemiology, pathophysiology, and management. Clin. Microbiol. Rev. 2005, 18, 383–416. [Google Scholar] [CrossRef] [PubMed]
- Willcocks, S.J.; Denman, C.C.; Atkins, H.S.; Wren, B.W. Intracellular replication of the well-armed pathogen Burkholderia pseudomallei. Curr. Opin. Microbiol. 2016, 29, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Merad, M.; Sathe, P.; Helft, J.; Miller, J.; Mortha, A. The Dendritic Cell Lineage: Ontogeny and Function of Dendritic Cells and Their Subsets in the Steady State and the Inflamed Setting. Annu. Rev. Immunol. 2013, 31, 563–604. [Google Scholar] [CrossRef] [PubMed]
- Parcina, M.; Wendt, C.; Goetz, F.; Zawatzky, R.; Zähringer, U.; Heeg, K.; Bekeredjian-Ding, I. Staphylococcus aureus—Induced Plasmacytoid Dendritic Cell Activation Is Based on an IgG-Mediated Memory Response. J. Immunol. 2008, 181, 3823–3833. [Google Scholar] [CrossRef] [PubMed]
- Veckman, V.; Julkunen, I. Streptococcus pyogenes activates human plasmacytoid and myeloid dendritic cells. J. Leukoc. Biol. 2008, 83, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Piccioli, D.; Sammicheli, C.; Tavarini, S.; Nuti, S.; Frigimelica, E.; Manetti, A.G.; Nuccitelli, A.; Aprea, S.; Valentini, S.; Borgogni, E.; et al. Human plasmacytoid dendritic cells are unresponsive to bacterial stimulation and require a novel type of cooperation with myeloid dendritic cells for maturation. Blood 2009, 113, 4232–4239. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.L.; Morris, J.L.; Rush, C.M.; Ketheesan, N. Plasmacytoid dendritic cell bactericidal activity against Burkholderia pseudomallei. Microbes Infect. 2015, 17, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Elvin, S.J.; Healey, G.D.; Westwood, A.; Knight, S.C.; Eyles, J.E.; Williamson, E.D. Protection against Heterologous Burkholderia pseudomallei Strains by Dendritic Cell Immunization. Infect. Immun. 2006, 74, 1706–1711. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sarkar-Tyson, M.; Smither, S.J.; Harding, S.V.; Atkins, T.P.; Titball, R.W. Protective efficacy of heat-inactivated B. thailandensis, B. mallei or B. pseudomallei against experimental melioidosis and glanders. Vaccine 2009, 27, 4447–4451. [Google Scholar] [CrossRef] [PubMed]
- Westera, L.; Drylewicz, J.; den Braber, I.; Mugwagwa, T.; van der Maas, I.; Kwast, L.; Volman, T.; van de Weg-Schrijver, E.H.; Bartha, I.; Spierenburg, G.; et al. Closing the gap between T-cell life span estimates from stable isotope-labeling studies in mice and humans. Blood 2013, 122, 2205–2212. [Google Scholar] [CrossRef]
- Tippayawat, P.; Saenwongsa, W.; Mahawantung, J.; Suwannasaen, D.; Chetchotisakd, P.; Limmathurotsakul, D.; Peacock, S.J.; Felgner, P.L.; Atkins, H.S.; Titball, R.W.; et al. Phenotypic and Functional Characterization of Human Memory T Cell Responses to Burkholderia pseudomallei. PLoS Negl. Trop. Dis. 2009, 3, e407. [Google Scholar] [CrossRef]
- Palucka, K.; Banchereau, J. Human dendritic cell subsets in vaccination. Curr. Opin. Immunol. 2013, 25, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Tel, J.; Aarntzen, E.H.; Baba, T.; Schreibelt, G.; Schulte, B.M.; Benitez-Ribas, D.; Boerman, O.C.; Croockewit, S.; Oyen, W.J.; van Rossum, M.; et al. Natural Human Plasmacytoid Dendritic Cells Induce Antigen-Specific T-Cell Responses in Melanoma Patients. Cancer Res. 2013, 73, 1063–1075. [Google Scholar] [CrossRef]
- Tacken, P.J.; de Vries, I.J.M.; Torensma, R.; Figdor, C.G. Dendritic-cell immunotherapy: From ex vivo loading to in vivo targeting. Nat. Rev. Immunol. 2007, 7, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.L.; Morris, J.L.; Rush, C.M.; Ketheesan, N. Migration of Dendritic Cells Facilitates Systemic Dissemination of Burkholderia pseudomallei. Infect. Immun. 2014, 82, 4233–4240. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reddi, D.; Durant, L.; Bernardo, D.; Noble, A.; English, N.R.; Hendy, P.; Clark, G.C.; Prior, J.L.; Williamson, E.D.; Knight, S.C. In Vitro Priming of Human T Cells by Dendritic Cells Provides a Screening Tool for Candidate Vaccines for Burkholderia pseudomallei. Vaccines 2021, 9, 929. https://doi.org/10.3390/vaccines9080929
Reddi D, Durant L, Bernardo D, Noble A, English NR, Hendy P, Clark GC, Prior JL, Williamson ED, Knight SC. In Vitro Priming of Human T Cells by Dendritic Cells Provides a Screening Tool for Candidate Vaccines for Burkholderia pseudomallei. Vaccines. 2021; 9(8):929. https://doi.org/10.3390/vaccines9080929
Chicago/Turabian StyleReddi, Durga, Lydia Durant, David Bernardo, Alistair Noble, Nicholas R. English, Philip Hendy, Graeme C. Clark, Joann L. Prior, Ethel Diane Williamson, and Stella C. Knight. 2021. "In Vitro Priming of Human T Cells by Dendritic Cells Provides a Screening Tool for Candidate Vaccines for Burkholderia pseudomallei" Vaccines 9, no. 8: 929. https://doi.org/10.3390/vaccines9080929
APA StyleReddi, D., Durant, L., Bernardo, D., Noble, A., English, N. R., Hendy, P., Clark, G. C., Prior, J. L., Williamson, E. D., & Knight, S. C. (2021). In Vitro Priming of Human T Cells by Dendritic Cells Provides a Screening Tool for Candidate Vaccines for Burkholderia pseudomallei. Vaccines, 9(8), 929. https://doi.org/10.3390/vaccines9080929