
Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Human PBMC Isolation
2.3. Immortalization of Human Memory B Cells
2.4. SPOT Peptide Synthesis
2.5. Expression and Purification of Recombinant Proteins and Peptides
2.6. Mice Immunization
2.7. ELISA
2.8. Neutralization Test
2.9. Indirect Immunofluorescence Assay
2.10. Statistical Analysis
3. Results
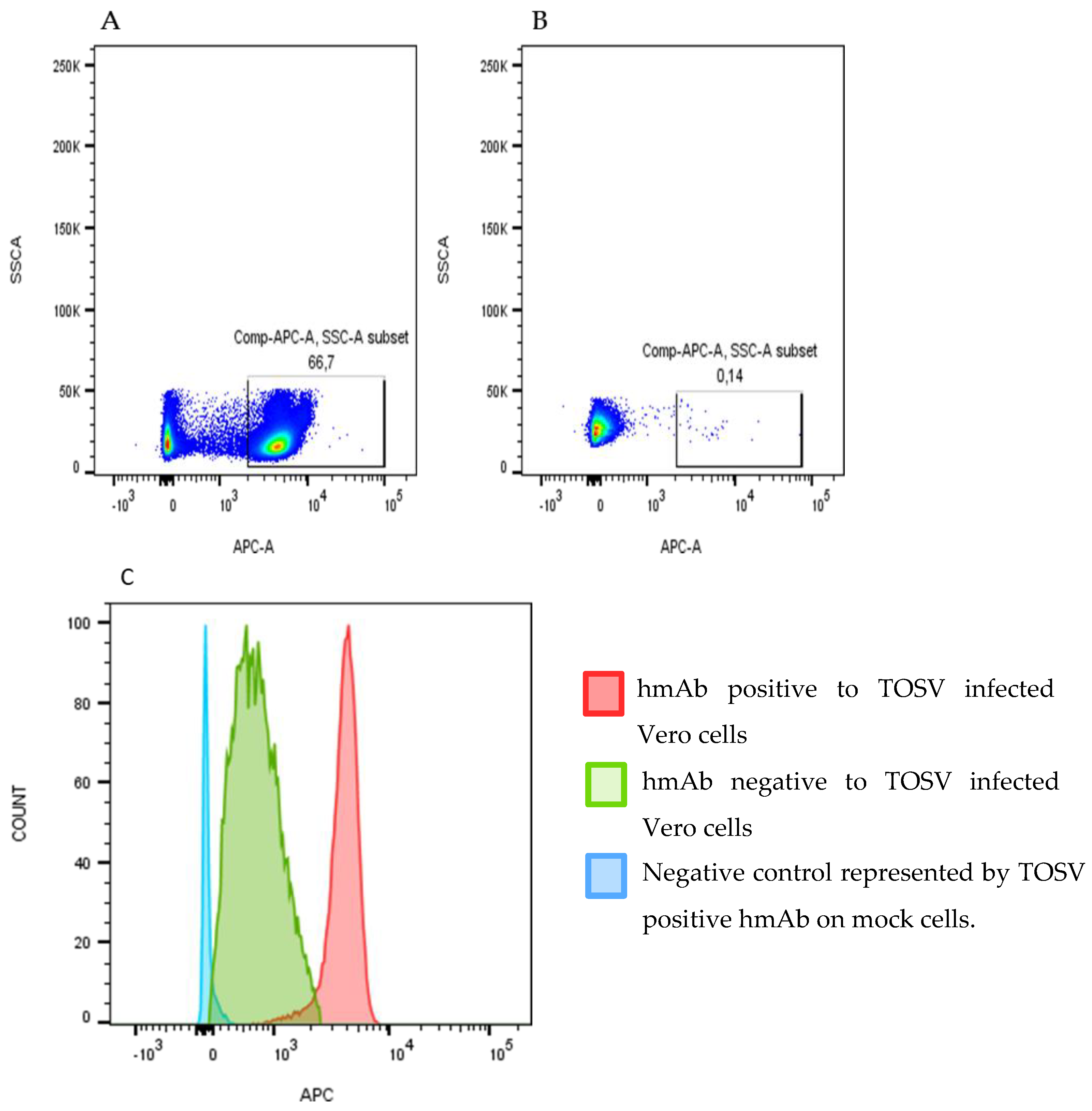
3.1. Isolation and Screening of a Panel of TOSV Human Monoclonal Antibodies
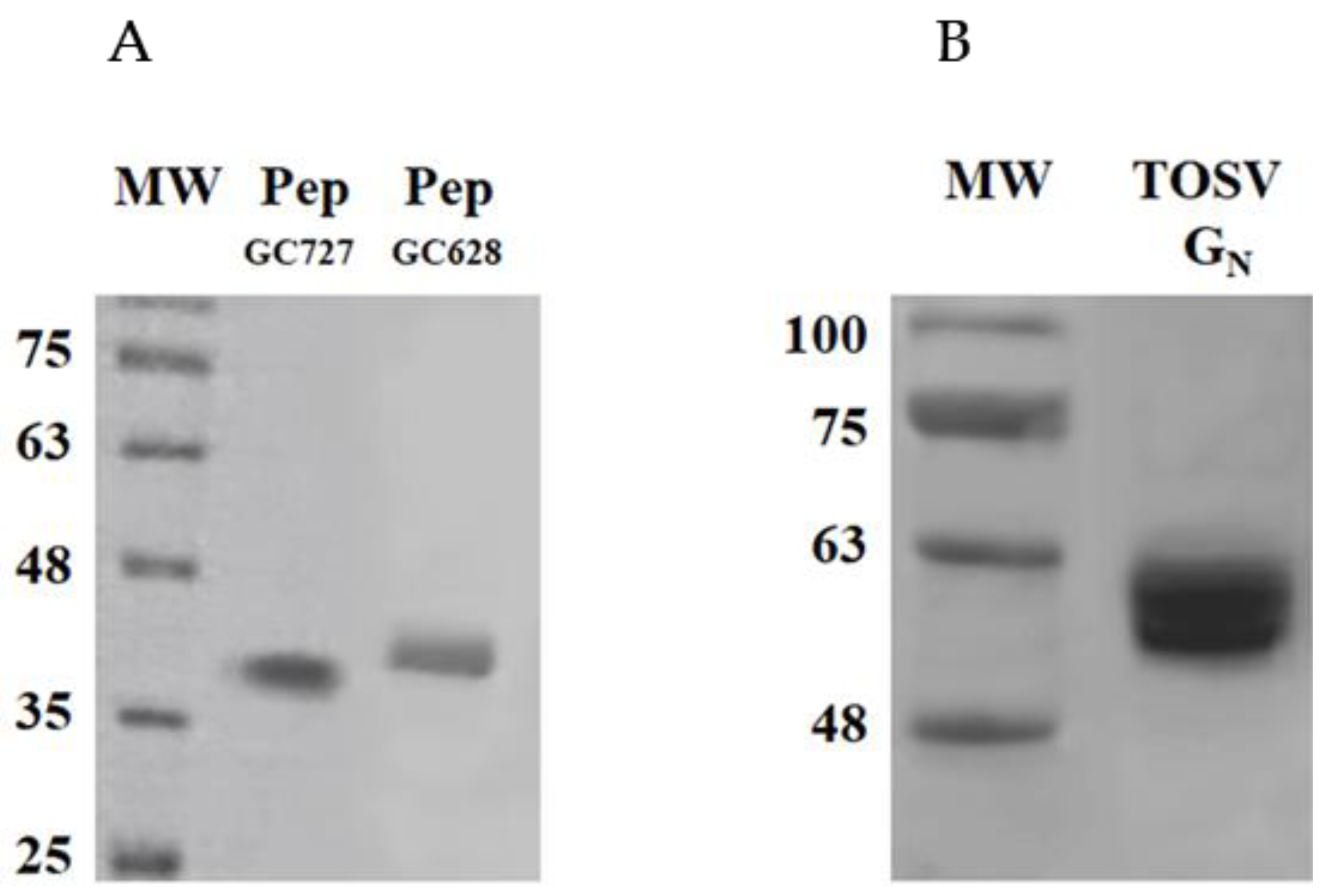
3.2. Identification of Neutralizing TOSV Gn Epitopes by Pepscan Analysis
3.3. Cross-Reactivity of Human mAbs
3.4. Immunization of Mice with Peptides
3.5. Serological Cross-Reactivity of Immunized Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Abudurexiti, A.; Adkins, S.; Alioto, D.; Alkhovsky, S.V.; Avšič-Županc, T.; Ballinger, M.J.; Bente, D.A.; Beer, M.; Bergeron, É.; Blair, C.D.; et al. Taxonomy of the order Bunyavirales: Update 2019. Arch. Virol. 2019, 164, 1949–1965. [Google Scholar] [CrossRef] [Green Version]
- Verani, P.; Ciufolini, M.G.; Nicoletti, L.; Balducci, M.; Sabatinelli, G.; Coluzzi, M.; Paci, P.; Amaducci, L. Ecological and epidemiological studies of Toscana virus, anarbovirus isolated from Phlebotomus. Ann. Ist. Super. Sanita 1982, 18, 397–399. [Google Scholar]
- Verani, P.; Nicoletti, L.; Ciufolini, M.G. Antigenic and biological characterization of Toscana virus, a newPhlebotomus fever group virus isolated in Italy. Acta Virol. 1984, 28, 39–47. [Google Scholar]
- Verani, P.; Ciufolini, M.G.; Caciolli, S.; Renzi, A.; Nicoletti, L.; Sabatinelli, G.; Bartolozzi, D.; Volpi, G.; Amaducci, L.; Coluzzi, M.; et al. Ecology of viruses isolated from sand flies in Italy and characterized of a new Phlebovirus (Arabia virus). Am. J. Trop. Med. Hyg. 1988, 38, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Valassina, M.; Cusi, M.G.; Valensin, P.E. A Mediterranean arbovirus: The Toscana virus. J. NeuroVirology 2003, 9, 577–583. [Google Scholar] [CrossRef]
- Valassina, M.; Valentini, M.; Pugliese, A.; Valensin, P.E.; Cusi, M.G. Serological survey of toscana virus infections in a high-risk population in Italy. Clin. Diagn. Lab. Immunol. 2003, 10, 483–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charrel Rèmi, N.; Pierre, G.; Josè-Maria, N.; Loredana, N.; Anna, P.; Paz, S.; Antonio, T.; de Lambelleire, X.E. Emergence of Toscana Virus in Europe. Emerg. Infect. Diseas. 2005, 11, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu-Gàmez, S.; Pèrez-Ruiz, M.; Palop-Borràs, B.; Navarro-Marì, J.M. Unusual Manifestation of Toscana virus Infection, Spain. Emerg. Infect. Dis. 2009, 15, 347–348. [Google Scholar] [CrossRef] [PubMed]
- Cusi, M.G.; GoriSavellini, G.; Zanelli, G. Toscana virus epidemiology: From Italy to beyond. Open Virol. J. 2010, 4, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Charrel, R.N.; Bichaud, L.; De Lamballerie, X. Emergence of Toscana virus in the Mediterranean area. World J. Virol. 2012, 1, 135–141. [Google Scholar] [CrossRef]
- Accardi, L.; Gro, M.C.; Di Bonito, P.; Giorgi, C. Toscana virus genomic L segment: Molecular cloning, coding strategy and amino acid sequence in comparison with other negative strand RNA viruses. Virus Res. 1993, 27, 119–131. [Google Scholar] [CrossRef]
- Di Bonito, P.; Mochi, S.; Gro, M.C.; Fortini, D.; Giorgi, C. Organization of the M genomic segment of Toscana phlebovirus. J. Gen. Virol. 1997, 76, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Gro, M.C.; Di Bonito, P.; Fortini, D.; Mochi, S.; Giorgi, C. Completion of molecular characterization of Toscana phlebovirus genome: Nucleotide sequence, coding strategy of M genomic segment and its aminoacid sequence comparison to other phleboviruses. Virus Res. 1997, 51, 81–91. [Google Scholar] [CrossRef]
- Giorgi, C.; Accardi, L.; Nicoletti, L.; Gro, M.C.; Takehara, K.; Hilditch, C.; Morikawa, S.; Bishop, D.H. Sequences and coding strategies of the SRNAs of Toscana and Rift Valley fever viruses compared to those of Punta Toro, Sicilian Sandfly fever, and Uukuniemi viruses. Virology 1991, 180, 738–753. [Google Scholar] [CrossRef]
- Saluzzo, J.F.; Anderson, G.W., Jr.; Hodgson, L.A.; Digoutte, J.P.; Smith, J.F. Antigenic and biological properties of Rift Valley fever virus isolated during the 1987 Mauritanian epidemic. Res. Virol. 1989, 140, 155–164. [Google Scholar] [CrossRef]
- Saluzzo, J.F.; Anderson, G.W., Jr.; Smith, J.F.; Fontenille, D.; Coulanges, P. Biological and antigenic relationship between Rift Valley fever virus strains isolated in Egypt and Madagascar. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 701. [Google Scholar] [CrossRef]
- Gonzalez-Scarano, F.; Shope, R.E.; Calisher, C.E.; Nathanson, N. Characterization of monoclonal antibodies against Gn and N proteins of La Crosse and Tahyna, two California serogroup bunyaviruses. Virology 1982, 120, 42–53. [Google Scholar] [CrossRef]
- Grady, L.; Kinch, W. Two monoclonal antibodies against La Crosse virus show host-dependent neutralizing activity. J. Gen. Virol. 1985, 66, 2773–2776. [Google Scholar] [CrossRef] [PubMed]
- Arikawa, J.; Schmaljohn, A.L.; Dalrymple, J.M.; Schmaljohn, C.S. Characterization of Hantaan virus envelope glycoprotein antigenic determinants defined by monoclonal antibodies. J. Gen. Virol. 1989, 70, 615–624. [Google Scholar] [CrossRef]
- Di Bonito, P.; Bosco, S.; Mochi, S.; Accardi, L.; Ciufolini, M.G.; Nicoletti, L.; Giorgi, C. Human antibody response to Toscana virus glycoproteins expressed by recombinant baculovirus. J. Med. Virol. 2002, 68, 615–619. [Google Scholar] [CrossRef]
- Cusi, M.G.; Valentini, M.; Valensin, P.E.; Valassina, M. Immune response to the neurotropic Toscana virus infection: Preliminary data. In Proceedings of the 1st SIV International Workshop on Neurovirology, Alghero, Italy, 23–25 June 2002. [Google Scholar]
- Cusi, M.G.; Valensin, P.E.; Donati, M.; Valassina, M. Neutralization of Toscana virus is partially mediated by antibodies to the nucleocapsid protein. J. Med. Virol. 2001, 63, 72–75. [Google Scholar] [CrossRef]
- Soldateschi, D.; dal Maso, G.M.; Valassina, M.; Santini, L.; Bianchi, S.; Cusi, M.G. Laboratory diagnosis of Toscana virus infection by enzyme immunoassay with recombinant viral nucleoprotein. J. Clin. Microbiol. 1999, 37, 649–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Authorisation, no. 9/2014—General Authorisation to Process Personal Data for Scientific Research Purposes. Available online: https://www.gazzettaufficiale.it/atto/serie_generale/caricaDettaglioAtto/originario?atto.dataPbblicazioneGazzetta=2014-1230&atto.codiceRedazionale=14A09916&elenco30giorni=true (accessed on 30 December 2014).
- Law Decree 22 December 2017, No. 219, published in the Ocial Gazette No. 12 of 16 January 2018. Available online: https://www.gazzettaufficiale.it/eli/gu/2018/01/16/12/sg/pdf (accessed on 16 January 2016).
- Traggiai, E.; Becker, S.; Subbarao, K.; Kolesnikova, L.; Uematsu, Y.; Gismondo, M.R.; Murphy, B.R.; Rappuoli, R.; Lanzavecchia, A. An efficient method to make human monoclonal antibodies from memory B cells: Potent neutralization of SARS coronavirus. Nat. Med. 2004, 10, 871–875. [Google Scholar] [CrossRef]
- Hilpert, K.; Winkler, D.F.; Hancock, R.E. Peptide arrays on cellulose support: SPOT synthesis, a time and cost-efficient method for synthesis of large numbers of peptides in a parallel and addressable fashion. Nat. Protoc. 2007, 2, 1333–1349. [Google Scholar] [CrossRef]
- Winkler, D.F.; Campbell, W.D. The spot technique: Synthesis and screening of peptide macroarrays on cellulose membranes. Methods. Mol. Biol. 2008, 494, 47–70. [Google Scholar] [PubMed]
- Lin, M.; Mcrae, H.; Dan, H.; Tangorra, E.; Laverdiere, A.; Pasick, J. High-resolution epitope mapping for monoclonal antibodies to the structural protein Erns of classical swine fever virus using peptide array and random peptide phage display approaches. J. Gen. Virol. 2010, 138, 2928–2940. [Google Scholar] [CrossRef] [PubMed]
- Di Bonito, P.; Nicoletti, L.; Mochi, S.; Accardi, L.; Marchi, A.; Giorgi, C. Immunological characterization of Toscana virus proteins. Arch. Virol. 1999, 144, 1947–1960. [Google Scholar] [CrossRef] [PubMed]
- Backovic, M.; Johansson, D.X.; Klupp, B.G.; Mettenleiter, T.C.; Persson, M.A.; Rey, F.A. Efficient method for production of high yields of Fab fragments in Drosophila S2 cells. Protein Eng. Des. Sel. 2010, 23, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Hlavacek, W.S.; Posner, R.G.; Perelson, A.S. Steric effects on multivalent ligand-receptor binding: Exclusion of ligand sites by bound cell surface receptors. Biophys. J. 1999, 76, 3031–3043. [Google Scholar] [CrossRef] [Green Version]
- Rizk, R.Z.; Christensen, N.D.; Michael, K.M.; Müller, M.; Sehr, P.; Waterboer, T.; Pawlita, M. Reactivity pattern of 92 monoclonal antibodies with 15 human papillomavirus types. J. Gen. Virol. 2008, 89 Pt 1, 117–129. [Google Scholar] [CrossRef]
- Elliott, R.M.; Brennan, B. Emerging phleboviruses. Curr. Opin. Virol. 2014, 5, 50–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Mice | Antigens Injected for Immunization |
|---|---|
| Group 1 | Peptide 1-KLH |
| Group 2 | Peptide 2-KLH |
| Group 3 | Peptide 3-KLH |
| Group 4 | Peptide 1-KLH and peptide 2-KLH |
| Group 5 | Peptide 1-KLH and peptide 3-KLH |
| Group 6 | Peptide 2-KLH and peptide 3-KLH |
| Group 7 | Peptide 1-KLH, peptide 2-KLH and peptide 3-KLH |
| Group 8 | Pep GC683 |
| Group 9 | Pep GC683 and peptide 3-KLH |
| Group 10 | Pep GC727 |
| Group 11 | rGn |
| Group 12 | PBS |
| mAbs of Acute Phase | |
| mAbs | NT Titer |
| 5 | neg |
| mAbs of Convalescent Phase | |
| mAbs | NT Titer |
| 26 | neg |
| 7 | <1/128 |
| 8 | 1/128–1/512 |
| 12 | >1/512 |
| Antibody ID | NT Titer | |||
|---|---|---|---|---|
| TOSV | SFNV | PUNV | SFSV | |
| TVB 27 | 1/161 | 1/25 | - | - |
| TVB 73 | >1/512 | - | - | - |
| TVB 147 | 1/645 | - | - | - |
| TVB 161 | 1/645 | - | - | - |
| TVB 164 | 1/6 | - | - | - |
| GROUPS | CROSS-REACTIVITY | |||
|---|---|---|---|---|
| SFNV | SFSV | CYPR | PUNV | |
| Group 1 (Pep 1-KLH) | + | − | − | − |
| Group 2 (Pep 2-KLH) | + | − | − | − |
| Group 3 (Pep 3-KLH) | + | − | − | + |
| Group 4 (Pep 1-KLH + Pep2-KLH) | + | + | + | − |
| Group 5 (Pep 1-KLH + Pep 3-KLH) | + | − | − | + |
| Group 6(Pep2-KLH + Pep3-KLH) | + | − | − | + |
| Group7(Pep1-KLH+Pep2-KLH + Pep3-KLH) | + | − | − | + |
| Group 8 (PepGc 683) | + | − | − | − |
| Group 9 (PepGc 683 + Pep3-KLH) | + | − | − | + |
| Group 10 (Pep Gc 727) | + | − | − | + |
| Group 11 (rGn) | − | − | − | − |
| Group 12 (PBS) | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandolfo, C.; Prathyumn, S.; Terrosi, C.; Anichini, G.; Gori Savellini, G.; Corti, D.; Bracci, L.; Lanzavecchia, A.; Roman-Sosa, G.; Cusi, M.G. Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein. Vaccines 2021, 9, 924. https://doi.org/10.3390/vaccines9080924
Gandolfo C, Prathyumn S, Terrosi C, Anichini G, Gori Savellini G, Corti D, Bracci L, Lanzavecchia A, Roman-Sosa G, Cusi MG. Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein. Vaccines. 2021; 9(8):924. https://doi.org/10.3390/vaccines9080924
Chicago/Turabian StyleGandolfo, Claudia, Shibily Prathyumn, Chiara Terrosi, Gabriele Anichini, Gianni Gori Savellini, Davide Corti, Luisa Bracci, Antonio Lanzavecchia, Gleyder Roman-Sosa, and Maria Grazia Cusi. 2021. "Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein" Vaccines 9, no. 8: 924. https://doi.org/10.3390/vaccines9080924
APA StyleGandolfo, C., Prathyumn, S., Terrosi, C., Anichini, G., Gori Savellini, G., Corti, D., Bracci, L., Lanzavecchia, A., Roman-Sosa, G., & Cusi, M. G. (2021). Identification of a Neutralizing Epitope on TOSV Gn Glycoprotein. Vaccines, 9(8), 924. https://doi.org/10.3390/vaccines9080924