
Neutrophils and Influenza: A Thin Line between Helpful and Harmful

 and
and 
{kind=link}
Abstract
1. Introduction
2. Murine vs. Human Neutrophils
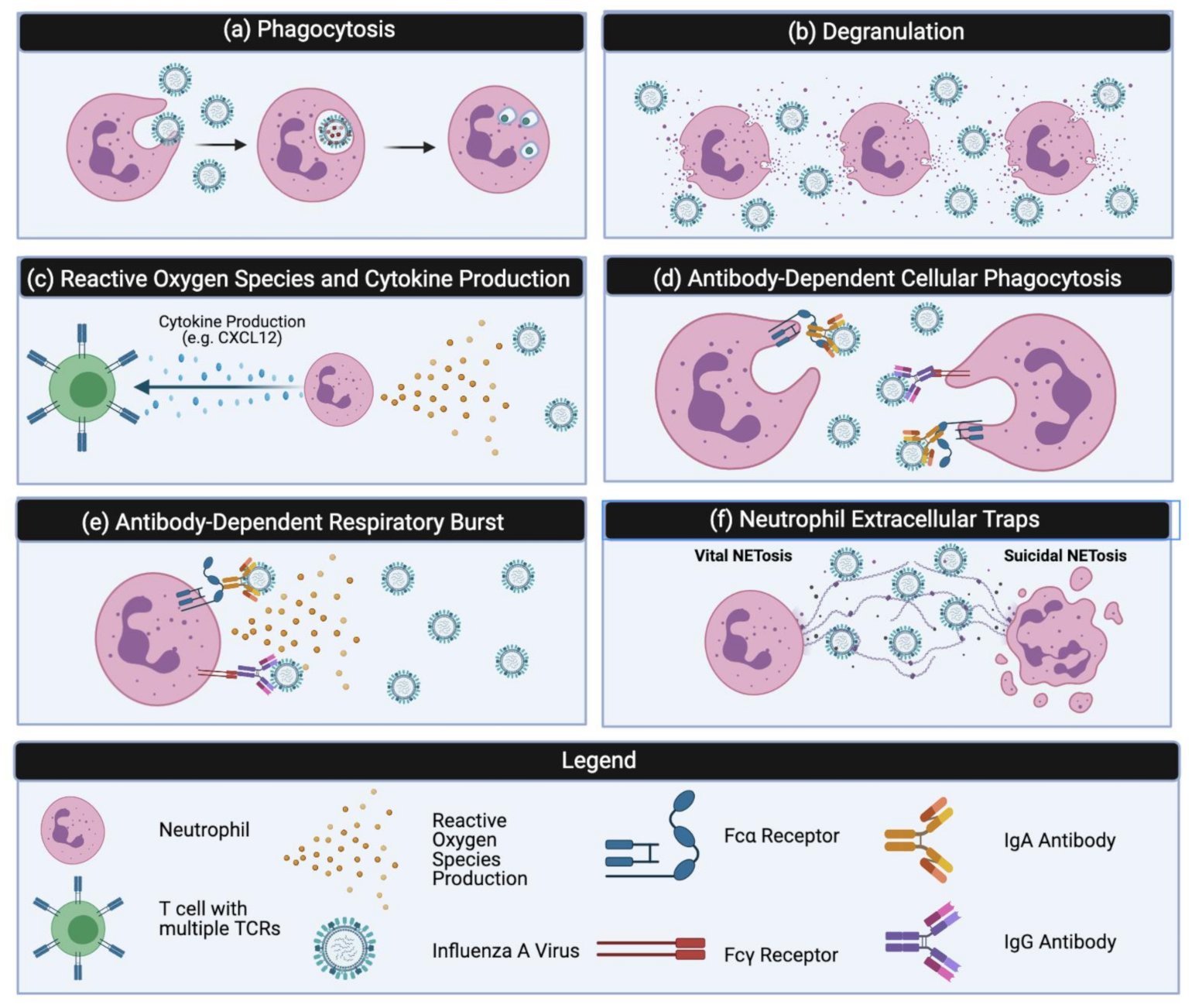
3. Classical Neutrophil Effector Functions
3.1. Phagocytosis
3.2. Degranulation
3.3. Reactive Oxygen Species (ROS)
3.4. Cytokine Production
4. Antibody Mediated Effector Functions
4.1. Antibody Dependent Cellular Phagocytosis
4.2. Antibody-Dependent Respiratory Burst
4.3. Neutrophil Extracellular Traps (NETs)
5. Influenza Viruses and Vaccines
6. Neutrophils and Influenza Virus Infection: A Complex Relationship
6.1. The Beneficial Roles of Neutrophils during Influenza Virus Infection
6.2. The Negative Roles of Neutrophils during Influenza Virus Infection
6.3. A Negligible Role for Neutrophils in Influenza Virus Infection
7. Implications and Considerations in HA-Targeted Universal Vaccine Design
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Minns, D.; Smith, K.J.; Alessandrini, V.; Hardisty, G.; Melrose, L.; Jackson-Jones, L.; MacDonald, A.S.; Davidson, D.J.; Gwyer Findlay, E. The Neutrophil Antimicrobial Peptide Cathelicidin Promotes Th17 Differentiation. Nat. Commun. 2021, 12, 1285. [Google Scholar] [CrossRef] [PubMed]
- Liew, P.X.; Kubes, P. The Neutrophil’s Role during Health and Disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Mackey, J.B.G.; Coffelt, S.B.; Carlin, L.M. Neutrophil Maturity in Cancer. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Cowland, J.B.; Borregaard, N. Granulopoiesis and Granules of Human Neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef]
- Devi, S.; Wang, Y.; Chew, W.K.; Lima, R.; A.-González, N.; Mattar, C.N.Z.; Chong, S.Z.; Schlitzer, A.; Bakocevic, N.; Chew, S.; et al. Neutrophil Mobilization via Plerixafor-Mediated CXCR4 Inhibition Arises from Lung Demargination and Blockade of Neutrophil Homing to the Bone Marrow. J. Exp. Med. 2013, 210, 2321–2336. [Google Scholar] [CrossRef]
- Köhler, A.; De Filippo, K.; Hasenberg, M.; van den Brandt, C.; Nye, E.; Hosking, M.P.; Lane, T.E.; Männ, L.; Ransohoff, R.M.; Hauser, A.E.; et al. G-CSF–Mediated Thrombopoietin Release Triggers Neutrophil Motility and Mobilization from Bone Marrow via Induction of Cxcr2 Ligands. Blood 2011, 117, 4349–4357. [Google Scholar] [CrossRef]
- Dancey, J.T.; Deubelbeiss, K.A.; Harker, L.A.; Finch, C.A. Neutrophil Kinetics in Man. J. Clin. Investig. 1976, 58, 705–715. [Google Scholar] [CrossRef]
- Lawrence, S.M.; Corriden, R.; Nizet, V. How Neutrophils Meet Their End. Trends Immunol. 2020, 41, 531–544. [Google Scholar] [CrossRef]
- Cartwright, G.E.; Athens, J.W.; Wintrobe, M.M. Analytical Review: The Kinetics of Granulopoiesis in Normal Man. Blood 1964, 24, 780–803. [Google Scholar] [CrossRef]
- Mauer, A.M.; Athens, J.W.; Ashenbrucker, H.; Cartwright, G.E.; Wintrobe, M.M. Leukokinetic Studies. II. A Method for Labeling Granulocytes in vitro with radioactive diisopropylfluorophosphate (DFP32)*. J. Clin. Investig. 1960, 39, 1481–1486. [Google Scholar] [CrossRef]
- Pillay, J.; den Braber, I.; Vrisekoop, N.; Kwast, L.M.; de Boer, R.J.; Borghans, J.A.M.; Tesselaar, K.; Koenderman, L. In Vivo Labeling with 2H2O Reveals a Human Neutrophil Lifespan of 5.4 Days. Blood 2010, 116, 625–627. [Google Scholar] [CrossRef]
- Martin, C.; Burdon, P.C.E.; Bridger, G.; Gutierrez-Ramos, J.-C.; Williams, T.J.; Rankin, S.M. Chemokines Acting via CXCR2 and CXCR4 Control the Release of Neutrophils from the Bone Marrow and Their Return Following Senescence. Immunity 2003, 19, 583–593. [Google Scholar] [CrossRef]
- Eash, K.J.; Greenbaum, A.M.; Gopalan, P.K.; Link, D.C. CXCR2 and CXCR4 Antagonistically Regulate Neutrophil Trafficking from Murine Bone Marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef]
- Schnoor, M.; Vadillo, E.; Guerrero-Fonseca, I.M. The Extravasation Cascade Revisited from a Neutrophil Perspective. Curr. Opin. Physiol. 2021, 19, 119–128. [Google Scholar] [CrossRef]
- Manley, H.R.; Keightley, M.C.; Lieschke, G.J. The Neutrophil Nucleus: An Important Influence on Neutrophil Migration and Function. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Alon, R.; Sportiello, M.; Kozlovski, S.; Kumar, A.; Reilly, E.C.; Zarbock, A.; Garbi, N.; Topham, D.J. Leukocyte Trafficking to the Lungs and beyond: Lessons from Influenza for COVID-19. Nat. Rev. Immunol. 2021, 21, 49–64. [Google Scholar] [CrossRef]
- Rosales, C. Neutrophils at the Crossroads of Innate and Adaptive Immunity. J. Leukoc. Biol. 2020, 108, 377–396. [Google Scholar] [CrossRef]
- Boxio, R.; Bossenmeyer, P.C.; Steinckwich, N.; Dournon, C.; Nüße, O. Mouse Bone Marrow Contains Large Numbers of Functionally Competent Neutrophils. J. Leukoc. Biol. 2004, 75, 604–611. [Google Scholar] [CrossRef]
- Southgate, E.L.; He, R.L.; Gao, J.-L.; Murphy, P.M.; Nanamori, M.; Ye, R.D. Identification of Formyl Peptides from Listeria Monocytogenes and Staphylococcus Aureus as Potent Chemoattractants for Mouse Neutrophils. J. Immunol. 2008, 181, 1429–1437. [Google Scholar] [CrossRef]
- Hasenberg, M.; Köhler, A.; Bonifatius, S.; Borucki, K.; Riek-Burchardt, M.; Achilles, J.; Männ, L.; Baumgart, K.; Schraven, B.; Gunzer, M. Rapid Immunomagnetic Negative Enrichment of Neutrophil Granulocytes from Murine Bone Marrow for Functional Studies In Vitro and In Vivo. PLoS ONE 2011, 6, e17314. [Google Scholar] [CrossRef]
- Risso, A. Leukocyte Antimicrobial Peptides: Multifunctional Effector Molecules of Innate Immunity. J. Leukoc. Biol. 2000, 68, 785–792. [Google Scholar] [CrossRef]
- Salvatore, M.; García-Sastre, A.; Ruchala, P.; Lehrer, R.I.; Chang, T.; Klotman, M.E. β-Defensin Inhibits Influenza Virus Replication by Cell-Mediated Mechanism(s). J. Infect. Dis. 2007, 196, 835–843. [Google Scholar] [CrossRef]
- Bruhns, P.; Jönsson, F. Mouse and Human FcR Effector Functions. Immunol. Rev. 2015, 268, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Hamdan, T.A.; Lang, P.A.; Lang, K.S. The Diverse Functions of the Ubiquitous Fcγ Receptors and Their Unique Constituent, FcRγ Subunit. Pathogens 2020, 9, 140. [Google Scholar] [CrossRef]
- Koernig, S.; Campbell, I.K.; Mackenzie-Kludas, C.; Schaub, A.; Loetscher, M.; Ng, W.C.; Zehnder, R.; Pelczar, P.; Sanli, I.; Alhamdoosh, M.; et al. Topical Application of Human-Derived Ig Isotypes for the Control of Acute Respiratory Infection Evaluated in a Human CD89-Expressing Mouse Model. Mucosal Immunol. 2019, 12, 1013–1024. [Google Scholar] [CrossRef]
- Launay, P.; Grossetête, B.; Arcos-Fajardo, M.; Gaudin, E.; Torres, S.P.; Beaudoin, L.; Patey-Mariaud de Serre, N.; Lehuen, A.; Monteiro, R.C. Fcalpha Receptor (CD89) Mediates the Development of Immunoglobulin A (IgA) Nephropathy (Berger’s Disease). Evidence for Pathogenic Soluble Receptor-Iga Complexes in Patients and CD89 Transgenic Mice. J. Exp. Med. 2000, 191, 1999–2009. [Google Scholar] [CrossRef]
- van Egmond, M.; van Vuuren, A.J.; Morton, H.C.; van Spriel, A.B.; Shen, L.; Hofhuis, F.M.; Saito, T.; Mayadas, T.N.; Verbeek, J.S.; van de Winkel, J.G. Human Immunoglobulin A Receptor (FcalphaRI, CD89) Function in Transgenic Mice Requires Both FcR Gamma Chain and CR3 (CD11b/CD18). Blood 1999, 93, 4387–4394. [Google Scholar] [CrossRef] [PubMed]
- Heijnen, I.A.; van Vugt, M.J.; Fanger, N.A.; Graziano, R.F.; de Wit, T.P.; Hofhuis, F.M.; Guyre, P.M.; Capel, P.J.; Verbeek, J.S.; van de Winkel, J.G. Antigen Targeting to Myeloid-Specific Human Fc Gamma RI/CD64 Triggers Enhanced Antibody Responses in Transgenic Mice. J. Clin. Investig. 1996, 97, 331–338. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, S.E.; Taylor, S.M.; Malladi, P.; Yuhan, H.; Cassel, D.L.; Chien, P.; Schwartz, E.; Schreiber, A.D.; Surrey, S.; Reilly, M.P. The Role of the Human Fc Receptor Fc Gamma RIIA in the Immune Clearance of Platelets: A Transgenic Mouse Model. J. Immunol. 1999, 162, 4311–4318. [Google Scholar] [PubMed]
- Li, F.; Ravetch, J.V. Inhibitory Fcγ Receptor Engagement Drives Adjuvant and Anti-Tumor Activities of Agonistic CD40 Antibodies. Science 2011, 333, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wirthmueller, U.; Ravetch, J.V. Reconstitution of Human Fc Gamma RIII Cell Type Specificity in Transgenic Mice. J. Exp. Med. 1996, 183, 1259–1263. [Google Scholar] [CrossRef]
- Mancardi, D.A.; Albanesi, M.; Jönsson, F.; Iannascoli, B.; Van Rooijen, N.; Kang, X.; England, P.; Daëron, M.; Bruhns, P. The High-Affinity Human IgG Receptor FcγRI (CD64) Promotes IgG-Mediated Inflammation, Anaphylaxis, and Antitumor Immunotherapy. Blood 2013, 121, 1563–1573. [Google Scholar] [CrossRef]
- Herant, M.; Heinrich, V.; Dembo, M. Mechanics of Neutrophil Phagocytosis: Experiments and Quantitative Models. J. Cell Sci. 2006, 119, 1903–1913. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Rieu, P.; Descamps-Latscha, B.; Lesavre, P.; Halbwachs-Mecarelli, L. Neutrophils: Molecules, Functions and Pathophysiological Aspects. Lab. Investig. 2000, 80, 617–653. [Google Scholar] [CrossRef]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The Multifaceted Functions of Neutrophils. Annu. Rev. Pathol 2014, 9, 181–218. [Google Scholar] [CrossRef]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil Function: From Mechanisms to Disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Dahlgren, C.; Karlsson, A.; Bylund, J. Intracellular Neutrophil Oxidants: From Laboratory Curiosity to Clinical Reality. J. Immunol. 2019, 202, 3127–3134. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Mitra, S.; Abraham, E. Participation of Superoxide in Neutrophil Activation and Cytokine Production. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2006, 1762, 732–741. [Google Scholar] [CrossRef]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine Production by Human Neutrophils: Revisiting the “Dark Side of the Moon”. Eur. J. Clin. Investig. 2018, 48, e12952. [Google Scholar] [CrossRef]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-Derived Cytokines: Facts beyond Expression. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Wang, Y.; Jönsson, F. Expression, Role, and Regulation of Neutrophil Fcγ Receptors. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Tay, M.Z.; Wiehe, K.; Pollara, J. Antibody-Dependent Cellular Phagocytosis in Antiviral Immune Responses. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Mullarkey, C.E.; Bailey, M.J.; Golubeva, D.A.; Tan, G.S.; Nachbagauer, R.; He, W.; Novakowski, K.E.; Bowdish, D.M.; Miller, M.S.; Palese, P. Broadly Neutralizing Hemagglutinin Stalk-Specific Antibodies Induce Potent Phagocytosis of Immune Complexes by Neutrophils in an Fc-Dependent Manner. mBio 2016, 7. [Google Scholar] [CrossRef]
- DiLillo, D.J.; Tan, G.S.; Palese, P.; Ravetch, J.V. Broadly Neutralizing Hemagglutinin Stalk–Specific Antibodies Require FcγR Interactions for Protection against Influenza Virus in Vivo. Nat. Med. 2014, 20, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Ana-Sosa-Batiz, F.; Vanderven, H.; Jegaskanda, S.; Johnston, A.; Rockman, S.; Laurie, K.; Barr, I.; Reading, P.; Lichtfuss, M.; Kent, S.J. Influenza-Specific Antibody-Dependent Phagocytosis. PLoS ONE 2016, 11, e0154461. [Google Scholar] [CrossRef]
- Joos, C.; Marrama, L.; Polson, H.E.J.; Corre, S.; Diatta, A.-M.; Diouf, B.; Trape, J.-F.; Tall, A.; Longacre, S.; Perraut, R. Clinical Protection from Falciparum Malaria Correlates with Neutrophil Respiratory Bursts Induced by Merozoites Opsonized with Human Serum Antibodies. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Sheehy, S.H.; Douglas, A.D.; Draper, S.J. Challenges of Assessing the Clinical Efficacy of Asexual Blood-Stage Plasmodium Falciparum Malaria Vaccines. Hum. Vaccin. Immunother. 2013, 9, 1831–1840. [Google Scholar] [CrossRef]
- Kapelski, S.; Klockenbring, T.; Fischer, R.; Barth, S.; Fendel, R. Assessment of the Neutrophilic Antibody-Dependent Respiratory Burst (ADRB) Response to Plasmodium Falciparum. J. Leukoc. Biol. 2014, 96, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, D.; de Cassan, S.C.; Williams, A.R.; Douglas, A.D.; Forbes, E.K.; Adame-Gallegos, J.R.; Shi, J.; Pleass, R.J.; Draper, S.J. Assessment of Antibody-Dependent Respiratory Burst Activity from Mouse Neutrophils on Plasmodium Yoelii Malaria Challenge Outcome. J. Leukoc. Biol. 2014, 95, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochem. Mosc. 2020, 85, 1178–1190. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: How Vital Is It? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel Cell Death Program Leads to Neutrophil Extracellular Traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil Elastase and Myeloperoxidase Regulate the Formation of Neutrophil Extracellular Traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef]
- Björnsdottir, H.; Welin, A.; Michaëlsson, E.; Osla, V.; Berg, S.; Christenson, K.; Sundqvist, M.; Dahlgren, C.; Karlsson, A.; Bylund, J. Neutrophil NET Formation Is Regulated from the inside by Myeloperoxidase-Processed Reactive Oxygen Species. Free Radic. Biol. Med. 2015, 89, 1024–1035. [Google Scholar] [CrossRef]
- Alemán, O.R.; Mora, N.; Cortes-Vieyra, R.; Uribe-Querol, E.; Rosales, C. Differential Use of Human Neutrophil Fcγ Receptors for Inducing Neutrophil Extracellular Trap Formation. J. Immunol. Res. 2016, 2016, e2908034. [Google Scholar] [CrossRef]
- Behnen, M.; Leschczyk, C.; Möller, S.; Batel, T.; Klinger, M.; Solbach, W.; Laskay, T. Immobilized Immune Complexes Induce Neutrophil Extracellular Trap Release by Human Neutrophil Granulocytes via FcγRIIIB and Mac-1. J. Immunol. 2014, 193, 1954–1965. [Google Scholar] [CrossRef]
- Aleyd, E.; van Hout, M.W.M.; Ganzevles, S.H.; Hoeben, K.A.; Everts, V.; Bakema, J.E.; van Egmond, M. IgA Enhances NETosis and Release of Neutrophil Extracellular Traps by Polymorphonuclear Cells via Fcα Receptor I. J. Immunol. 2014, 192, 2374–2383. [Google Scholar] [CrossRef]
- Stacey, H.D.; Golubeva, D.; Posca, A.; Ang, J.C.; Novakowski, K.E.; Zahoor, M.A.; Kaushic, C.; Cairns, E.; Bowdish, D.M.E.; Mullarkey, C.E.; et al. IgA Potentiates NETosis in Response to Viral Infection. bioRxiv 2021. [Google Scholar] [CrossRef]
- Jenne, C.N.; Wong, C.H.Y.; Zemp, F.J.; McDonald, B.; Rahman, M.M.; Forsyth, P.A.; McFadden, G.; Kubes, P. Neutrophils Recruited to Sites of Infection Protect from Virus Challenge by Releasing Neutrophil Extracellular Traps. Cell Host Microbe 2013, 13, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil Extracellular Traps Mediate a Host Defense Response to Human Immunodeficiency Virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef]
- Muraro, S.P.; De Souza, G.F.; Gallo, S.W.; Da Silva, B.K.; De Oliveira, S.D.; Vinolo, M.A.R.; Saraiva, E.M.; Porto, B.N. Respiratory Syncytial Virus Induces the Classical ROS-Dependent NETosis through PAD-4 and Necroptosis Pathways Activation. Sci. Rep. 2018, 8, 14166. [Google Scholar] [CrossRef] [PubMed]
- Khandpur, R.; Carmona-Rivera, C.; Vivekanandan-Giri, A.; Gizinski, A.; Yalavarthi, S.; Knight, J.S.; Friday, S.; Li, S.; Patel, R.M.; Subramanian, V.; et al. NETs Are a Source of Citrullinated Autoantigens and Stimulate Inflammatory Responses in Rheumatoid Arthritis. Sci. Transl. Med. 2013, 5, 178ra40. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yang, P.; Gao, M.; Yu, T.; Shi, Y.; Zhang, M.; Yao, M.; Liu, Y.; Zhang, X. NLRP3 Activation Induced by Neutrophil Extracellular Traps Sustains Inflammatory Response in the Diabetic Wound. Clin. Sci. 2019, 133, 565–582. [Google Scholar] [CrossRef]
- Bruschi, M.; Bonanni, A.; Petretto, A.; Vaglio, A.; Pratesi, F.; Santucci, L.; Migliorini, P.; Bertelli, R.; Galetti, M.; Belletti, S.; et al. Neutrophil Extracellular Traps Profiles in Patients with Incident Systemic Lupus Erythematosus and Lupus Nephritis. J. Rheumatol. 2020, 47, 377–386. [Google Scholar] [CrossRef]
- Jenne, C.N.; Kubes, P. Virus-Induced NETs—Critical Component of Host Defense or Pathogenic Mediator? PLoS Pathog. 2015, 11. [Google Scholar] [CrossRef]
- Tang, B.M.; Shojaei, M.; Teoh, S.; Meyers, A.; Ho, J.; Ball, T.B.; Keynan, Y.; Pisipati, A.; Kumar, A.; Eisen, D.P.; et al. Neutrophils-Related Host Factors Associated with Severe Disease and Fatality in Patients with Influenza Infection. Nat. Commun. 2019, 10, 3422. [Google Scholar] [CrossRef]
- Peteranderl, C.; Herold, S.; Schmoldt, C. Human Influenza Virus Infections. Semin. Respir. Crit. Care Med. 2016, 37, 487–500. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. Influenza: The Once and Future Pandemic. Public Health Rep. 2010, 125, 16–26. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. The Pathology of Influenza Virus Infections. Annu. Rev. Pathol. 2008, 3, 499–522. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Kash, J.C. Influenza Virus Evolution, Host Adaptation and Pandemic Formation. Cell Host Microbe 2010, 7, 440–451. [Google Scholar] [CrossRef]
- Taubenberger, J.K.; Morens, D.M. Influenza Viruses: Breaking All the Rules. mBio 2013, 4. [Google Scholar] [CrossRef]
- Krammer, F.; Smith, G.J.D.; Fouchier, R.A.M.; Peiris, M.; Kedzierska, K.; Doherty, P.C.; Palese, P.; Shaw, M.L.; Treanor, J.; Webster, R.G.; et al. Influenza. Nat. Rev. Dis. Prim. 2018, 4, 1–21. [Google Scholar] [CrossRef]
- Paules, C.; Subbarao, K. Influenza. Lancet 2017, 390, 697–708. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26, D49–D53. [Google Scholar] [CrossRef]
- Karakus, U.; Thamamongood, T.; Ciminski, K.; Ran, W.; Günther, S.C.; Pohl, M.O.; Eletto, D.; Jeney, C.; Hoffmann, D.; Reiche, S.; et al. MHC Class II Proteins Mediate Cross-Species Entry of Bat Influenza Viruses. Nature 2019, 567, 109–112. [Google Scholar] [CrossRef]
- Giotis, E.S.; Carnell, G.; Young, E.F.; Ghanny, S.; Soteropoulos, P.; Wang, L.-F.; Barclay, W.S.; Skinner, M.A.; Temperton, N. Entry of the Bat Influenza H17N10 Virus into Mammalian Cells Is Enabled by the MHC Class II HLA-DR Receptor. Nat. Microbiol. 2019, 4, 2035–2038. [Google Scholar] [CrossRef]
- Banerjee, A.; Mossman, K.L.; Miller, M.S. Bat Influenza Viruses: Making a Double Agent of MHC Class II. Trends Microbiol. 2020, 28, 703–706. [Google Scholar] [CrossRef]
- Zhu, X.; Yang, H.; Guo, Z.; Yu, W.; Carney, P.J.; Li, Y.; Chen, L.-M.; Paulson, J.C.; Donis, R.O.; Tong, S.; et al. Crystal Structures of Two Subtype N10 Neuraminidase-like Proteins from Bat Influenza A Viruses Reveal a Diverged Putative Active Site. Proc. Natl. Acad. Sci. USA 2012, 109, 18903–18908. [Google Scholar] [CrossRef]
- Li, Q.; Sun, X.; Li, Z.; Liu, Y.; Vavricka, C.J.; Qi, J.; Gao, G.F. Structural and Functional Characterization of Neuraminidase-like Molecule N10 Derived from Bat Influenza A Virus. Proc. Natl. Acad. Sci. USA 2012, 109, 18897–18902. [Google Scholar] [CrossRef] [PubMed]
- Houser, K.; Subbarao, K. Influenza Vaccines: Challenges and Solutions. Cell Host Microbe 2015, 17, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H. Seasonal Influenza and Vaccine Herd Effect. Clin. Exp. Vaccine Res. 2014, 3, 128–132. [Google Scholar] [CrossRef]
- Plans-Rubió, P. The Vaccination Coverage Required to Establish Herd Immunity against Influenza Viruses. Prev. Med. 2012, 55, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Soema, P.C.; Kompier, R.; Amorij, J.-P.; Kersten, G.F.A. Current and next Generation Influenza Vaccines: Formulation and Production Strategies. Eur. J. Pharm. Biopharm. 2015, 94, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Liu, Y.-M.; Tseng, Y.-C.; Ma, C. Better Influenza Vaccines: An Industry Perspective. J. Biomed. Sci. 2020, 27, 33. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Bijker, E.M. A Guide to Vaccinology: From Basic Principles to New Developments. Nat. Rev. Immunol. 2021, 21, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Heaton, N.S.; Sachs, D.; Chen, C.-J.; Hai, R.; Palese, P. Genome-Wide Mutagenesis of Influenza Virus Reveals Unique Plasticity of the Hemagglutinin and NS1 Proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20248–20253. [Google Scholar] [CrossRef]
- DiLillo, D.J.; Palese, P.; Wilson, P.C.; Ravetch, J.V. Broadly Neutralizing Anti-Influenza Antibodies Require Fc Receptor Engagement for in Vivo Protection. J. Clin. Investig. 2016, 126, 605–610. [Google Scholar] [CrossRef]
- Corti, D.; Voss, J.; Gamblin, S.J.; Codoni, G.; Macagno, A.; Jarrossay, D.; Vachieri, S.G.; Pinna, D.; Minola, A.; Vanzetta, F.; et al. A Neutralizing Antibody Selected from Plasma Cells That Binds to Group 1 and Group 2 Influenza A Hemagglutinins. Science 2011, 333, 850–856. [Google Scholar] [CrossRef]
- He, W.; Tan, G.S.; Mullarkey, C.E.; Lee, A.J.; Lam, M.M.W.; Krammer, F.; Henry, C.; Wilson, P.C.; Ashkar, A.A.; Palese, P.; et al. Epitope Specificity Plays a Critical Role in Regulating Antibody-Dependent Cell-Mediated Cytotoxicity against Influenza A Virus. Proc. Natl. Acad. Sci. USA 2016, 113, 11931–11936. [Google Scholar] [CrossRef]
- He, W.; Chen, C.-J.; Mullarkey, C.E.; Hamilton, J.R.; Wong, C.K.; Leon, P.E.; Uccellini, M.B.; Chromikova, V.; Henry, C.; Hoffman, K.W.; et al. Alveolar Macrophages Are Critical for Broadly-Reactive Antibody-Mediated Protection against Influenza A Virus in Mice. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Leon, P.E.; He, W.; Mullarkey, C.E.; Bailey, M.J.; Miller, M.S.; Krammer, F.; Palese, P.; Tan, G.S. Optimal Activation of Fc-Mediated Effector Functions by Influenza Virus Hemagglutinin Antibodies Requires Two Points of Contact. Proc. Natl. Acad. Sci. USA 2016, 113, E5944–E5951. [Google Scholar] [CrossRef]
- Perrone, L.A.; Plowden, J.K.; García-Sastre, A.; Katz, J.M.; Tumpey, T.M. H5N1 and 1918 Pandemic Influenza Virus Infection Results in Early and Excessive Infiltration of Macrophages and Neutrophils in the Lungs of Mice. PLoS Pathog. 2008, 4, e1000115. [Google Scholar] [CrossRef]
- Wang, J.P.; Bowen, G.N.; Padden, C.; Cerny, A.; Finberg, R.W.; Newburger, P.E.; Kurt-Jones, E.A. Toll-like Receptor–Mediated Activation of Neutrophils by Influenza A Virus. Blood 2008, 112, 2028–2034. [Google Scholar] [CrossRef]
- de Jong, M.D.; Simmons, C.P.; Thanh, T.T.; Hien, V.M.; Smith, G.J.D.; Chau, T.N.B.; Hoang, D.M.; Van Vinh Chau, N.; Khanh, T.H.; Dong, V.C.; et al. Fatal Outcome of Human Influenza A (H5N1) Is Associated with High Viral Load and Hypercytokinemia. Nat. Med. 2006, 12, 1203–1207. [Google Scholar] [CrossRef]
- Weiland, J.E.; Davis, W.B.; Holter, J.F.; Mohammed, J.R.; Dorinsky, P.M.; Gadek, J.E. Lung Neutrophils in the Adult Respiratory Distress Syndrome. Clinical and Pathophysiologic Significance. Am. Rev. Respir. Dis. 1986, 133, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Rebetz, J.; Semple, J.W.; Kapur, R. The Pathogenic Involvement of Neutrophils in Acute Respiratory Distress Syndrome and Transfusion-Related Acute Lung Injury. Transfus. Med. Hemother. 2018, 45, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Bendib, I.; de Chaisemartin, L.; Granger, V.; Schlemmer, F.; Maitre, B.; Hüe, S.; Surenaud, M.; Beldi-Ferchiou, A.; Carteaux, G.; Razazi, K.; et al. Neutrophil Extracellular Traps Are Elevated in Patients with Pneumonia-Related Acute Respiratory Distress Syndrome. Anesthesiology 2019, 130, 581–591. [Google Scholar] [CrossRef]
- Stackowicz, J.; Jönsson, F.; Reber, L.L. Mouse Models and Tools for the in Vivo Study of Neutrophils. Front. Immunol. 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Daley, J.M.; Thomay, A.A.; Connolly, M.D.; Reichner, J.S.; Albina, J.E. Use of Ly6G-Specific Monoclonal Antibody to Deplete Neutrophils in Mice. J. Leukoc. Biol. 2008, 83, 64–70. [Google Scholar] [CrossRef]
- Fujisawa, H. Neutrophils Play an Essential Role in Cooperation with Antibody in Both Protection against and Recovery from Pulmonary Infection with Influenza Virus in Mice. J. Virol. 2008, 82, 2772–2783. [Google Scholar] [CrossRef]
- Tate, M.D.; Deng, Y.-M.; Jones, J.E.; Anderson, G.P.; Brooks, A.G.; Reading, P.C. Neutrophils Ameliorate Lung Injury and the Development of Severe Disease during Influenza Infection. J. Immunol. 2009, 183, 7441–7450. [Google Scholar] [CrossRef]
- Tate, M.D.; Brooks, A.G.; Reading, P.C.; Mintern, J.D. Neutrophils Sustain Effective CD8(+) T-Cell Responses in the Respiratory Tract Following Influenza Infection. Immunol. Cell Biol. 2012, 90, 197–205. [Google Scholar] [CrossRef]
- Pillay, J.; Kamp, V.M.; van Hoffen, E.; Visser, T.; Tak, T.; Lammers, J.-W.; Ulfman, L.H.; Leenen, L.P.; Pickkers, P.; Koenderman, L. A Subset of Neutrophils in Human Systemic Inflammation Inhibits T Cell Responses through Mac-1. J. Clin. Investig. 2012, 122, 327–336. [Google Scholar] [CrossRef]
- Tak, T.; Rygiel, T.P.; Karnam, G.; Bastian, O.W.; Boon, L.; Viveen, M.; Coenjaerts, F.E.; Meyaard, L.; Koenderman, L.; Pillay, J. Neutrophil-Mediated Suppression of Influenza-Induced Pathology Requires CD11b/CD18 (MAC-1). Am. J. Respir. Cell Mol. Biol. 2017, 58, 492–499. [Google Scholar] [CrossRef]
- Lim, K.; Hyun, Y.-M.; Lambert-Emo, K.; Capece, T.; Bae, S.; Miller, R.; Topham, D.J.; Kim, M. Neutrophil Trails Guide Influenza-Specific CD8+ T Cells in the Airways. Science 2015, 349. [Google Scholar] [CrossRef]
- Pang, I.K.; Ichinohe, T.; Iwasaki, A. IL-1R Signaling in Dendritic Cells Replaces Pattern-Recognition Receptors in Promoting CD8+ T Cell Responses to Influenza A Virus. Nat. Immunol. 2013, 14, 246–253. [Google Scholar] [CrossRef]
- Neyt, K.; GeurtsvanKessel, C.H.; Deswarte, K.; Hammad, H.; Lambrecht, B.N. Early IL-1 Signaling Promotes IBALT Induction after Influenza Virus Infection. Front. Immunol. 2016, 7, 312. [Google Scholar] [CrossRef]
- Schmitz, N.; Kurrer, M.; Bachmann, M.F.; Kopf, M. Interleukin-1 Is Responsible for Acute Lung Immunopathology but Increases Survival of Respiratory Influenza Virus Infection. J. Virol. 2005, 79, 6441–6448. [Google Scholar] [CrossRef]
- Wong, C.K.; Smith, C.A.; Sakamoto, K.; Kaminski, N.; Koff, J.L.; Goldstein, D.R. Aging Impairs Alveolar Macrophage Phagocytosis and Increases Influenza-Induced Mortality in Mice. J. Immunol. 2017, 199, 1060–1068. [Google Scholar] [CrossRef] [PubMed]
- Peiró, T.; Patel, D.F.; Akthar, S.; Gregory, L.G.; Pyle, C.J.; Harker, J.A.; Birrell, M.A.; Lloyd, C.M.; Snelgrove, R.J. Neutrophils Drive Alveolar Macrophage IL-1β Release during Respiratory Viral Infection. Thorax 2018, 73, 546–556. [Google Scholar] [CrossRef]
- Allen, I.C.; Scull, M.A.; Moore, C.B.; Holl, E.K.; McElvania-TeKippe, E.; Taxman, D.J.; Guthrie, E.H.; Pickles, R.J.; Ting, J.P.-Y. The NLRP3 Inflammasome Mediates in Vivo Innate Immunity to Influenza A Virus through Recognition of Viral RNA. Immunity 2009, 30, 556–565. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.G.; Dash, P.; Aldridge, J.R.; Ellebedy, A.H.; Reynolds, C.; Funk, A.J.; Martin, W.J.; Lamkanfi, M.; Webby, R.J.; Boyd, K.L.; et al. NLRP3 (NALP3/CIAS1/Cryopyrin) Mediates Key Innate and Healing Responses to Influenza A Virus via the Regulation of Caspase-1. Immunity 2009, 30, 566–575. [Google Scholar] [CrossRef]
- Niu, J.; Wu, S.; Chen, M.; Xu, K.; Guo, Q.; Lu, A.; Zhao, L.; Sun, B.; Meng, G. Hyperactivation of the NLRP3 Inflammasome Protects Mice against Influenza A Virus Infection via IL-1β Mediated Neutrophil Recruitment. Cytokine 2019, 120, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Short, K.R.; Kroeze, E.J.B.V.; Fouchier, R.A.M.; Kuiken, T. Pathogenesis of Influenza-Induced Acute Respiratory Distress Syndrome. Lancet Infect. Dis. 2014, 14, 57–69. [Google Scholar] [CrossRef]
- Sugamata, R.; Dobashi, H.; Nagao, T.; Yamamoto, K.; Nakajima, N.; Sato, Y.; Aratani, Y.; Oshima, M.; Sata, T.; Kobayashi, K.; et al. Contribution of Neutrophil-Derived Myeloperoxidase in the Early Phase of Fulminant Acute Respiratory Distress Syndrome Induced by Influenza Virus Infection. Microbiol. Immunol. 2012, 56, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, L.; Zhang, Y.; Pu, L.; Liu, J.; Li, X.; Chen, Z.; Hao, Y.; Wang, B.; Han, J.; et al. High Level of Neutrophil Extracellular Traps Correlates With Poor Prognosis of Severe Influenza A Infection. J. Infect. Dis. 2018, 217, 428–437. [Google Scholar] [CrossRef]
- Shieh, W.-J.; Blau, D.M.; Denison, A.M.; DeLeon-Carnes, M.; Adem, P.; Bhatnagar, J.; Sumner, J.; Liu, L.; Patel, M.; Batten, B.; et al. 2009 Pandemic Influenza A (H1N1). Am. J. Pathol. 2010, 177, 166–175. [Google Scholar] [CrossRef]
- Grunwell, J.R.; Giacalone, V.D.; Stephenson, S.; Margaroli, C.; Dobosh, B.S.; Brown, M.R.; Fitzpatrick, A.M.; Tirouvanziam, R. Neutrophil Dysfunction in the Airways of Children with Acute Respiratory Failure Due to Lower Respiratory Tract Viral and Bacterial Coinfections. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Deng, G.; Bi, J.; Kong, F.; Li, X.; Xu, Q.; Dong, J.; Zhang, M.; Zhao, L.; Luan, Z.; Lv, N.; et al. Acute Respiratory Distress Syndrome Induced by H9N2 Virus in Mice. Arch. Virol. 2010, 155, 187–195. [Google Scholar] [CrossRef]
- Xu, T.; Qiao, J.; Zhao, L.; Wang, G.; He, G.; Li, K.; Tian, Y.; Gao, M.; Wang, J.; Wang, H.; et al. Acute Respiratory Distress Syndrome Induced by Avian Influenza A (H5N1) Virus in Mice. Am. J. Respir. Crit. Care Med. 2006, 174, 1011–1017. [Google Scholar] [CrossRef]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.-A.; Phoon, M.C.; van Rooijen, N.; Chow, V.T. Excessive Neutrophils and Neutrophil Extracellular Traps Contribute to Acute Lung Injury of Influenza Pneumonitis. Am. J. Pathol. 2011, 179, 199–210. [Google Scholar] [CrossRef]
- Steinberg, K.P.; Milberg, J.A.; Martin, T.R.; Maunder, R.J.; Cockrill, B.A.; Hudson, L.D. Evolution of Bronchoalveolar Cell Populations in the Adult Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 1994, 150, 113–122. [Google Scholar] [CrossRef]
- Fry, A.M.; Shay, D.K.; Holman, R.C.; Curns, A.T.; Anderson, L.J. Trends in Hospitalizations for Pneumonia among Persons Aged 65 Years or Older in the United States, 1988–2002. JAMA 2005, 294, 2712–2719. [Google Scholar] [CrossRef]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality Associated with Influenza and Respiratory Syncytial Virus in the United States. JAMA 2003, 289, 179–186. [Google Scholar] [CrossRef]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Brammer, L.; Bridges, C.B.; Cox, N.J.; Fukuda, K. Influenza-Associated Hospitalizations in the United States. JAMA 2004, 292, 1333–1340. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, J.; Legge, K.; Perlman, S. Age-Related Increases in PGD2 Expression Impair Respiratory DC Migration, Resulting in Diminished T Cell Responses upon Respiratory Virus Infection in Mice. J. Clin. Investig. 2011, 121, 4921–4930. [Google Scholar] [CrossRef]
- Stout-Delgado, H.W.; Vaughan, S.E.; Shirali, A.C.; Jaramillo, R.J.; Harrod, K.S. Impaired NLRP3 Inflammasome Function in Elderly Mice during Influenza Infection Is Rescued by Treatment with Nigericin. J. Immunol. 2012, 188, 2815–2824. [Google Scholar] [CrossRef]
- Kulkarni, U.; Zemans, R.L.; Smith, C.A.; Wood, S.C.; Deng, J.C.; Goldstein, D.R. Excessive Neutrophil Levels in the Lung Underlie the Age-Associated Increase in Influenza Mortality. Mucosal Immunol. 2019, 12, 545–554. [Google Scholar] [CrossRef]
- Grégoire, M.; Uhel, F.; Lesouhaitier, M.; Gacouin, A.; Guirriec, M.; Mourcin, F.; Dumontet, E.; Chalin, A.; Samson, M.; Berthelot, L.-L.; et al. Impaired Efferocytosis and Neutrophil Extracellular Trap Clearance by Macrophages in ARDS. Eur. Respir. J. 2018, 52. [Google Scholar] [CrossRef]
- Scott, B.N.V.; Kubes, P. Death to the Neutrophil! A Resolution for Acute Respiratory Distress Syndrome? Eur. Respir. J. 2018, 52. [Google Scholar] [CrossRef]
- Menk, M.; Estenssoro, E.; Sahetya, S.K.; Neto, A.S.; Sinha, P.; Slutsky, A.S.; Summers, C.; Yoshida, T.; Bein, T.; Ferguson, N.D. Current and Evolving Standards of Care for Patients with ARDS. Intensiv. Care Med. 2020, 46, 2157–2167. [Google Scholar] [CrossRef]
- Carlet, J.; Payen, D.; Opal, S.M. Steroids for Sepsis and ARDS: This Eternal Controversy Remains with COVID-19. Lancet 2020, 396, e61–e62. [Google Scholar] [CrossRef]
- Ackermann, M.; Anders, H.-J.; Bilyy, R.; Bowlin, G.L.; Daniel, C.; De Lorenzo, R.; Egeblad, M.; Henneck, T.; Hidalgo, A.; Hoffmann, M.; et al. Patients with COVID-19: In the Dark-NETs of Neutrophils. Cell Death Differ. 2021, 1–15. [Google Scholar] [CrossRef]
- Bianchi, M.; Hakkim, A.; Brinkmann, V.; Siler, U.; Seger, R.A.; Zychlinsky, A.; Reichenbach, J. Restoration of NET Formation by Gene Therapy in CGD Controls Aspergillosis. Blood 2009, 114, 2619–2622. [Google Scholar] [CrossRef]
- Anjani, G.; Vignesh, P.; Joshi, V.; Shandilya, J.K.; Bhattarai, D.; Sharma, J.; Rawat, A. Recent Advances in Chronic Granulomatous Disease. Genes Dis. 2019, 7, 84–92. [Google Scholar] [CrossRef]
- Tate, M.D.; Ioannidis, L.J.; Croker, B.; Brown, L.E.; Brooks, A.G.; Reading, P.C. The Role of Neutrophils during Mild and Severe Influenza Virus Infections of Mice. PLoS ONE 2011, 6, e17618. [Google Scholar] [CrossRef] [PubMed]
- Wareing, M.D.; Shea, A.L.; Inglis, C.A.; Dias, P.B.; Sarawar, S.R. CXCR2 Is Required for Neutrophil Recruitment to the Lung during Influenza Virus Infection, but Is Not Essential for Viral Clearance. Viral Immunol. 2007, 20, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Nachbagauer, R.; Margine, I.; Tan, G.S.; Hirsh, A.; Krammer, F. Vaccination with Soluble Headless Hemagglutinin Protects Mice from Challenge with Divergent Influenza Viruses. Vaccine 2015, 33, 3314–3321. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Miller, M.S.; Hai, R.; Ryder, A.B.; Rose, J.K.; Palese, P.; García-Sastre, A.; Krammer, F.; Albrecht, R.A. Hemagglutinin Stalk Immunity Reduces Influenza Virus Replication and Transmission in Ferrets. J. Virol. 2016, 90, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Hai, R.; Yondola, M.; Tan, G.S.; Leyva-Grado, V.H.; Ryder, A.B.; Miller, M.S.; Rose, J.K.; Palese, P.; García-Sastre, A.; et al. Assessment of Influenza Virus Hemagglutinin Stalk-Based Immunity in Ferrets. J. Virol. 2014, 88, 3432–3442. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Pica, N.; Hai, R.; Margine, I.; Palese, P. Chimeric Hemagglutinin Influenza Virus Vaccine Constructs Elicit Broadly Protective Stalk-Specific Antibodies. J. Virol. 2013, 87, 6542–6550. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Margine, I.; Hai, R.; Flood, A.; Hirsh, A.; Tsvetnitsky, V.; Chen, D.; Palese, P. H3 Stalk-Based Chimeric Hemagglutinin Influenza Virus Constructs Protect Mice from H7N9 Challenge. J. Virol. 2014, 88, 2340–2343. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, S.T.; Lai, J.; Ma, J.; Stacey, H.D.; Miller, M.S.; Mullarkey, C.E. Neutrophils and Influenza: A Thin Line between Helpful and Harmful. Vaccines 2021, 9, 597. https://doi.org/10.3390/vaccines9060597
George ST, Lai J, Ma J, Stacey HD, Miller MS, Mullarkey CE. Neutrophils and Influenza: A Thin Line between Helpful and Harmful. Vaccines. 2021; 9(6):597. https://doi.org/10.3390/vaccines9060597
Chicago/Turabian StyleGeorge, Sneha T., Jonathan Lai, Julia Ma, Hannah D. Stacey, Matthew S. Miller, and Caitlin E. Mullarkey. 2021. "Neutrophils and Influenza: A Thin Line between Helpful and Harmful" Vaccines 9, no. 6: 597. https://doi.org/10.3390/vaccines9060597
APA StyleGeorge, S. T., Lai, J., Ma, J., Stacey, H. D., Miller, M. S., & Mullarkey, C. E. (2021). Neutrophils and Influenza: A Thin Line between Helpful and Harmful. Vaccines, 9(6), 597. https://doi.org/10.3390/vaccines9060597