Abstract
Acinetobacter baumannii is a leading cause of nosocomial infections that severely threaten public health. The formidable adaptability and resistance of this opportunistic pathogen have hampered the development of antimicrobial therapies which consequently leads to very limited treatment options. We mapped the global prevalence of multidrug-resistant A. baumannii and showed that carbapenem-resistant A. baumannii is widespread throughout Asia and the Americas. Moreover, when antimicrobial resistance rates of Acinetobacter spp. exceed a threshold level, the proportion of A. baumannii isolates from clinical samples surges. Therefore, vaccines represent a realistic alternative strategy to tackle this pathogen. Research into anti-A. baumannii vaccines have enhanced in the past decade and multiple antigens have been investigated preclinically with varying results. This review summarises the current knowledge of virulence factors relating to A. baumannii–host interactions and its implication in vaccine design, with a view to understanding the current state of A. baumannii vaccine development and the direction of future efforts.
1. Introduction
Acinetobacter baumannii is an opportunistic gram-negative coccobacillus that demonstrates exquisite survival under various environmental conditions and intrinsic resistance to routinely prescribed antibiotics [1]. Multidrug-resistant (MDR) A. baumannii strains are now prevalent worldwide, with the highest percentage of carbapenem resistance of over 90% being reported in the Mediterranean region, imposing serious burdens on healthcare systems. The vast majority of A. baumannii isolates arise in medical institutions and are closely associated with nosocomial infections, particularly in patients receiving intensive care and in immunocompromised individuals. Ventilator-associated pneumonia (VAP), central line-associated bloodstream infections (BSI), urinal tract infections (UTI), and meningitis are the most common clinical manifestations [2]. Unfortunately, the lack of efficacious treatments has led to high crude mortality ranging from 40% to 80% for infections occurring in sterile sites [3,4,5,6]. In 2017, the World Health Organization (WHO) listed carbapenem-resistant A. baumannii as a critical priority, for which, new antibiotics are urgently needed [7]. In addition, once nosocomial outbreaks of A. baumannii occur, it is difficult to thoroughly eradicate from the environment due to its remarkable resistance to disinfectants and its capacity to rapidly develop tolerance to these antibacterial agents, contributing to prolonged colonisation and transmission [8]. Therefore, vaccination of susceptible individuals is likely to be a more effective intervention for the prevention of A. baumannii infections. Compared with other nosocomial pathogens such as Pseudomonas aeruginosa, Staphylococcus aureus, and Clostridium difficile, the search for vaccines against A. baumannii is in its infancy [9]. Over the last decade, increased research efforts have concentrated on identifying potential virulence factors, clarifying host–pathogen interactions, and developing vaccines against this exquisitely resilient pathogen. In this review, we describe these advances and also provide a comprehensive appraisal of the relevant microbiology, epidemiology, and antibiotic resistance of A. baumannii.
The genus Acinetobacter currently comprises more than 60 validly named species [10]. It has been classified in the family Moraxellaceae, within the order Pseudomonadales, and the class Gammaproteobacteria since 1991 [11]. The whole genome of A. baumannii (strain ATCC 17978) was first sequenced in 2007 by Smith et al. [12]. However, its unambiguous identification in routine diagnostic laboratories has long been hindered as A. baumannii is phenotypically similar to, and phylogenetically close to, several other Acinetobacter species (Figure 1). The A. calcoaceticus–A. baumannii (Acb) complex consists of six valid species: A. baumannii, A. calcoaceticus (formerly Acinetobacter genomic species 1), A. nosocomialis (formerly Acinetobacter genomic species 13TU) [13], A. pittii (formerly Acinetobacter genomic species 3) [13], A. seifertii (formerly Acinetobacter genomic species ‘close to 13TU’) [14], and the most recently added A. lactucae (formerly Acinetobacter NB14, synonymous with A. dijkshoorniae) [10,15,16,17]. While A. calcoaceticus is non-pathogenic, the other five members are all clinically relevant and are frequently identified in a broad sense as A. baumannii, or A. baumannii group, in clinical microbial identification. The dominant species among the clinically isolated A. baumannii group vary with region [18,19], but from a clinical perspective, A. baumannii is the most virulent [5,20].
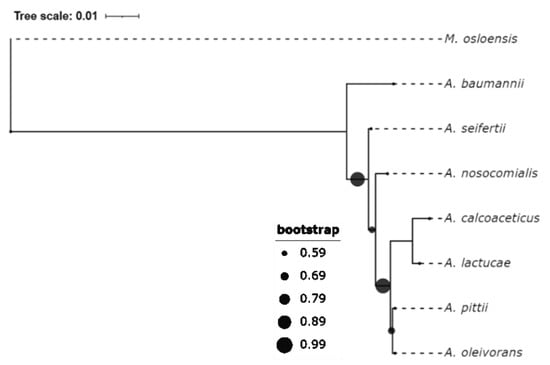
Figure 1.
Phylogenetic analysis of the Acb complex. The phylogeny tree was inferred with MEGA X on a Clustal W alignment of the 16S rRNA in the selected type strains. The maximum likelihood-kimura 2-parameter (ML-K2) model was used. Moraxella osloensis was chosen as an outgroup. Strain names and their corresponding GenBank accession numbers are listed in Table S1, Supplementary Materials. A. oleivorans has also been catalogued into the Acb group in some studies [21,22,23], though its name has not yet been validated.
2. A. baumannii Is Ubiquitous
Acinetobacter spp. are ubiquitous organisms and the first isolate, described as Micrococcus calcoaceticus back in 1911, was from soil [24]. Their widely distributed natural reservoirs occupy nearly all environmental niches including waterbodies, soil, mines, crude oil, sewage, sludge, solid surfaces, raw meat, cheese, milk, unprocessed vegetables, human skin, wild animals, livestock, fish, shrimps, plants, and nectar [25]. The natural habitats of A. baumannii, however, are still poorly defined as it is almost exclusively isolated from hospital environments and communities involving close contact [2]. A. baumannii was rarely detected on human skin (0.5%, n = 186), indicating that this is not the natural habitat of A. baumannii, despite Acinetobacter spp. being associated with human skin flora [26]. The presence of A. baumannii in the human body louse has been reported in many countries worldwide [27,28,29]. For example, the highly antibiotic-susceptible A. baumannii strain SDF which is responsible for community-acquired infections was first isolated from the interior of body lice collected on homeless people in France [30].
Identification of traces of A. baumannii in environmental soil and waterbody samples has frequently been reported [31,32,33,34,35,36,37]. An early study reported that more than a third of the soil samples from Hong Kong contained Acinetobacter spp., with A. baumannii representing 14.7% (n = 34) [38]. More recently, Hrenovic et al. isolated an antibiotic-resistant A. baumannii strain from acid paleosol in Croatia [39]. Most water-recovered A. baumannii strains were from sewage or urban streams, and hence it is challenging to define whether they are native residents or contaminants from other A. baumannii positive fomites [34,35,36,37]. A. baumannii colonisation has also been observed in livestock animals, mostly from cattle and poultry [37,40,41,42,43], with various reports in horses, pigs, donkeys, mules, goats, rabbits, dogs, cats, as well as the food products made from them [37,44], but the frequency varies depending on the geographic location of the farms. Wilharm et al. investigated the relatedness of A. baumannii isolates from livestock avian in Germany to the lineages spread in hospitals worldwide and found there were several strains closely related to human clinical isolates indicating that livestock avian may serve as a powerful vector for the spread of A. baumannii [43]. Considering the complex human–environment–livestock interactions and the elusive transmission chains, these reported cases may be relevant to ‘One Health’ infection control approaches. To date, localised studies have shown considerable variability in the prevalence among birds. For example, an isolation rate of 25% of white stork nestlings in Poland was documented, ranging from 4% to 48% [43]. In contrast, a recent regional study demonstrated A. baumannii barely existed in the choana and rectums of wild songbirds and gulls captured in Germany and Poland (0.19%, n = 1051) [45]. More culture-based screenings of avian populations worldwide, including their living environments are necessary. Genomic approaches may facilitate the linkage between environmental strains and clinical isolates in time excluding misleading strains, that are hospital-derived but contaminating other places.
3. Antimicrobial Resistance
The antimicrobial resistance of A. baumannii is of major concern. It is classified as an ESKAPE pathogen, an acronym referring to Enterococcus faecium, S. aureus, Klebsiella pneumoniae, A. baumannii, P. aeruginosa, and Enterobacter spp., all of which are recognised nosocomial bacteria with high potential to exhibit MDR and virulence [46]. Clinical isolates A. baumannii strains are frequently reported to demonstrate resistance to the most routinely prescribed antibiotics in varying degrees, including carbapenems such as imipenem and meropenem. Carbapenem is one of the β-lactam antibiotics with the broadest spectrum and highest efficiency. Resistance to carbapenem is considered to be a marker for extensively resistant bacteria because it involves a broad range of co-resistance to unrelated antibiotic classes [7]. We have collated an up-to-date profile on the global incidence of carbapenem-resistant A. baumannii strains documented over the past decade (Figure 2). The parameter R% was defined as the percentage of carbapenem-resistant isolates among all clinically isolated A. baumannii strains. Data for each country or region were collected from their latest antimicrobial resistance (AMR) surveillance annual reports, and in the case of those without applicable surveillance data, published figures from the most recent research or reviews were used. It is apparent that carbapenem-resistant A. baumannii is now problematic across Asia and the Americas, except in Japan and Canada. Moreover, Oceania, Western Europe, the Nordic region, and part of central Europe have the lowest R% (<10%), however, in areas surrounding the Mediterranean, including southern Europe, the Middle East, North Africa, up to 90% of A. baumannii clinical isolates are resistant to carbapenems. Details for central Africa and island countries are limited.
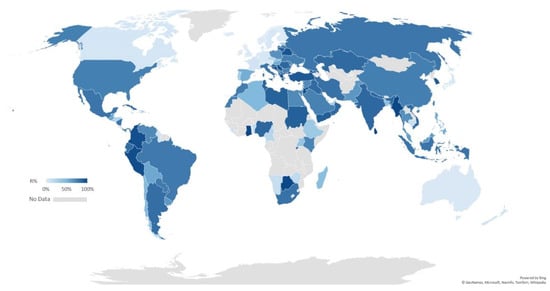
Figure 2.
Global percentage of resistance to carbapenems (R%) of A. baumannii. The shade of blue represents the level of carbapenem resistance worldwide during the last decade (2009–2018). No suitable surveillance or research data is available for areas coloured in grey. A full list of all the statistics and sources is provided in Table S2, Supplementary Materials.
In the treatment of A. baumannii infections, MDR often compels the adoption of last-resort antibiotics such as colistin when essentially no other options are available. However, the increasing quantity of recent cases reporting the emergence of pan-drug-resistant (PDR) strains tolerant to last-resort therapies has dimmed their potential [47,48,49,50,51,52].
4. Clinical Importance
4.1. Susceptible Populations
A. baumannii has been long been recognised as a major opportunistic pathogen causing nosocomial infections, or healthcare-associated infections, and manifests as a broad range of infections, including respiratory infections, BSIs, UTIs, and meningitis. [2]. Patients admitted to intensive care units (ICU), patients with indwelling medical devices, or with immunocompromised medical conditions are particularly predisposed to its infections. Falagas et al. demonstrated that the length of ICU stay was significantly increased in patients with A. baumannii infections [53]. Previous studies demonstrated that the crude mortality rate of A. baumannii infection was over 50% [3], although the attributable mortality varies with infection type. For example, in a multicentre study conducted in eight US metropolitan areas from 2012–2015, the overall recorded death rate of carbapenem-resistant A. baumannii infections was 17.9%, of which infections occurring at normally sterile sites had much higher mortality of 41.3% compared with UTIs (8.3%) [3]. A. baumannii is one of the leading causes of VAP worldwide, especially in Asia, Latin America, and the Middle East [54]. The ICU mortality of MDR A. baumannii caused VAP has been reported to be as high as 84.3% [4]. Crude mortality in patients with A. baumannii bacteraemia ranges from 37% to 52% in the U.S. [5]. Although the incidence of A. baumannii meningitis is comparably low, it is an increasing threat in post-neurosurgical patients, with a mortality rate approaching 70% [6]. A. baumannii skin and soft tissue infections in trauma (wound, burn) victims following natural disasters or wars have also been recorded, such as those during the Iraq conflicts, the Syrian war, the Wenchuan earthquake, the Marmara earthquake, and the Indian Ocean tsunami [47,55,56,57,58]. According to a single site study, after the Wenchuan earthquake, 14% of wound infections in the hospitalised survivors were caused by A. baumannii [58]. Noticeably, during the Coronavirus disease 2019 (COVID-19) pandemic, hospital-acquired co- or secondary infections in COVID-19 in-patients caused by A. baumannii were reported worldwide [59,60,61]. An acute care hospital in New Jersey reported 70% (14/20) of patients with hospital-acquired A. baumannii infections developed VAP and 85% (17/20) of them had co-infections with SARS-CoV-2 [61]. In a serious outbreak in Texas, COVID-19 patients with secondary A. baumannii infections experienced a two-fold higher mortality rate relative to COVID-19 patients without A. baumannii infections. These reports suggested that A. baumannii co-infection may exacerbate the development and prognosis of COVID-19 and hinder the clinical diagnosis and treatment [62].
Community-acquired A. baumannii infections are rarely reported but comparably more often in some tropical or sub-tropical areas in the Asia–Pacific such as northern Australia and Taiwan [63,64,65,66], indicating the potential climate drivers of its prevalence [67]. Unlike healthcare-associated infections, A. baumannii strains involved in community-acquired infection are more susceptible to antibiotics. Farrugia et al. sequenced the whole genome of a community strain D1279779 obtained from Darwin, Australia, and found that the antibiotic resistance island AbaR, associated with nosocomial strains, was absent in that strain [68]. Despite their low occurrence, community-acquired A. baumannii infections are characterised by an acute fulminant course involving septic shock, respiratory failure, severe sepsis, pneumonia, and the mortality can be quite high, i.e., over 60% [67,68].
4.2. Prevalence
Since the 1990s, outbreaks of A. baumannii in clinical sites have often been reported [69], particularly in regions with high levels of carbapenem-resistant rates. In Europe, the clinical isolation rate ranges from less than 1% to over 30% [70,71], and clinical outbreaks are most intensively reported in eastern and southeast Europe. We defined F% as the proportion of Acinetobacter spp. isolates identified among all the clinical samples of monitored AMR species (Escherichia coli, K. pneumoniae, P. aeruginosa, Acinetobacter spp., Streptococcus pneumoniae, S. aureus, and Enterobacter spp.). The parameter F% was compared to R% by region (Figure 2) using the 2018 data from two international surveillance networks, the European Antimicrobial Resistance Surveillance Network (EARS-Net) initiated by the European Centre for Disease Prevention and Control (ECDC) and the Central Asian and European Surveillance of Antimicrobial Resistance Network (CAESAR) from WHO, which together covered the entire European data where both the world’s lowest (<1%) and the highest resistant rates (>95%) can be found (Table S3, Supplementary Materials). There is a strong positive correlation between the two variables with a Spearman’s rank correlation coefficient (ρ) of 0.808 (p-value of 0.000). Curve estimation (Figure 3) shows the data points fit best in the exponential regression model described by the equation of F% = 0.7015 × e0.0304×R% (adjusted R2 = 0.736). This exponential model suggests that when antibiotic resistance rates are low in a region, the clinical isolation rates of Acinetobacter spp. generally remain relatively stable, but once the R% exceeds a certain level, the proportion of A. baumannii isolates from clinical samples surges, a stark warning that there might be considerably more A. baumannii associated infections and a higher risk of nosocomial outbreaks in the future. In this case, with an R% of less than 10%, the detecting frequency F% can be as low as <1%; whereas F% increases by ten-fold if the R% is more than 90%. However, due to the small sample size (n = 41) and high variance, the relationships between the prevalence of A. baumannii outbreaks and its antibiotic resistance should be further validated with more relevant data. No evidence of gender or age difference in incidence has been reported so far.
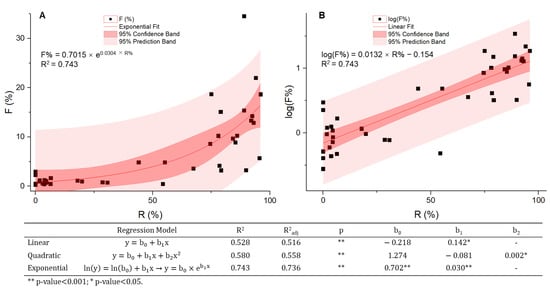
Figure 3.
Correlations between R% and F%. Correlation analysis and curve estimation were performed with SPSS and presented on linear (A) and logarithmic (B) scales. Both data sets follow non-normal distributions, and the correlation coefficient was calculated using Spearman’s rank-order method. R% and F% are in strong positive correlation with the best fit in the exponential model.
Regional differences in the overall proportion of A. baumannii among all clinical isolated, aerobic, and facultative gram-negative pathogens have been reported, ranging from 0.7% in North America to 4.6% in the Middle East [72]. The Middle East is one of the areas hardest hit by A. baumannii. Historically, A. baumannii was also known as ‘Iraqibacter’ because it caused outbreaks of MDR strains in the US military hospitals based in Iraq, Kuwait, and Afghanistan during the Iraq War [55,73]. These strains then spread outside the army to the civilian health care system, most probably via the transfer of colonised casualties [1,55], and have continued to be a persistent issue in these areas. Unfortunately, although serious infections and mortality caused by MDR A. baumannii have been reported [47,51,74], regional epidemiology studies are quite limited. A recent MLST-based study showed that sequence types ST2, ST195, and ST208 had been repeatedly reported from many Middle Eastern countries [75]. Moreover, US service members continue to be colonised with ‘Iraqibacteria’ and, although controversial, they may serve as reservoirs participating in the community transmission [76,77]. For example, a record review performed on the prevalence of MDR pathogens in a burn centre dedicated to military personnel reported that A. baumannii was the most prevalent organism recovered (22%) during the study period of 2003–2008 [78]. Subsequently, from 2009 to 2013, an equivalent level was reported in a study of nationwide data [79]. In contrast, the proportion was approximately 10% before 2000 [80], suggesting that A. baumannii has spread from the US military system to the civilian settings.
5. Virulence Factors
Severe tissue damage with infiltration of large numbers of inflammatory cells is frequently observed at A. baumannii infectious sites. High levels of interleukin 2 (IL-2), IL-4, IL-17, tumour necrosis factor α (TNF-α) and interferon γ (IFN-γ), as well as a lower CD4+/CD8+ ratio, have been reported [81]. The virulence of A. baumannii is strain-dependent and involves complicated mechanisms of pathogenicity which are only beginning to be elucidated. Many efficacious vaccines against other pathogens have been developed based on virulence factors, consequently, A. baumannii virulence factors are worth reviewing.
5.1. Adhesins and Invasins
Studies have demonstrated that the respiratory epithelial cells are more easily invaded by A. baumannii than non-respiratory epithelial cells [82], which likely contributes to the high occurrence and mortality rate of A. baumannii pneumonia. Outer membrane proteins (OMP), including OmpA (also known as Omp38), are the most well-studied A. baumannii adhesins [82,83,84,85] (Table 1). OmpA is a conserved, abundant porin in A. baumannii with a molecular weight of 38 kDa and plays an essential role in adhesion and invasion. Competitive binding studies with recombinant A. baumannii OmpA attenuated both cell binding and invasion [82]. A. baumannii attaches with moderate affinity to epithelial cells, secreting OmpA into the cells, which leads to bacterial uptake via actin rearrangement and membrane reorganisation of host cells, known as a zipper-like mechanism [82]. The internalised bacteria are located in membrane-bound vacuoles [82] and the internalised OmpA is translocated to the nucleus and mitochondria, causing the release of a series of proapoptotic molecules, which promotes cell apoptosis [83,84]. Consequently, OmpA also acts as a toxin contributing to pathogenesis. Notably, we ran BLASTp against the Swiss-Prot database and found that the A. baumannii OmpA (UniProt ID: Q6RYW5) has no significant homology with E. coli (K12) OmpA (96% query cover, 23% identities) but shares 46% and 41% identities with probable lipoprotein YiaD and YfiB, respectively. Considering the query covers are relatively low (31% for both), the results should be interpreted cautiously. Similarly, Omp33–36 (also known as Omp34) is another outer membrane virulence factor involved in host–cell attachment. It is also cytotoxic, inducing apoptosis by activation of caspases and modulation of autophagy, with the consequent accumulation of sequestosome 1 and autophagosome LC3B-II [86]. An isogenic A. baumannii mutant deficient in the omp33 gene resulted in a significant reduction in adherence, invasion, and cytotoxicity of human lung epithelial cells, compared to the wild type [86,87]. Other OMPs functioning as A. baumannii adhesins include the biofilm-associated protein (Bap), which increases the membrane hydrophobicity [88]; the FhaB/FhaC and CdiA/CdiB type Vb secretion system [89]; and the Acinetobacter trimeric autotransporter (Ata). Ata mediates adherence through binding to type IV collagen in the extracellular matrix (including basal lamina) of host cells [90,91,92]. It also contributes to cytotoxicity by inducing the secretion of pro-inflammatory cytokines, such as IL-6 and IL-8, and leading to cellular apoptosis in a caspase-dependent manner [92].
Recently, non-OMP adhesins and invasins have been identified. Both the response regulator, BfmR, and the sensor kinase, BfmS, in the BfmRS two-component system have been reported to play a role in cell adherence [93,94]. Several pili have been found to be relevant with the attachment to both biotic and abiotic surfaces, indicating that the motility may correlate with the adherence ability. Wood et al. showed that A. baumannii is capable of producing different adhesins and pilus assembly systems in response to changes in environmental conditions, such as the BlsA mediated, photoregulated type I pilus assembly system PrpABCD [95]. Eijkelkamp et al. found that type IV pili, associated with twitching motility, may also mediate adhesion [96]. Interestingly, they later found the expression of a homologue of the histone-like nucleoid structuring (H-NS) protein, a known global transcriptional repressor, was disrupted in an A. baumannii strain with enhanced adherence and motility [97]. Thioredoxin-A protein (Trx-A) is a mediator of the type IV pilus system and hence, is also involved in host–cell adhesion [98]. In contrast, the chaperone-usher (Csu) type I pili does not play a role in adherence [99]. Phospholipase C (PLC) and phospholipase D (PLD) both play significant roles in invasion and their inactivation led to impaired epithelial cell invasion and a reduction of the extrapulmonary tissue bacterial burden [100,101,102]. OMPs containing phosphorylcholine (ChoP) are also involved in bacterial adherence and invasion [103]. A. baumannii bound via ChoP to the platelet-activating factor receptor (PAFR) on human lung cells, resulting in activation of signalling involving G protein, clathrin, β-arrestins, proteins involved in the direction of the vacuolar movement, and intracellular calcium, which eventually led to invasion [103]. Host cell receptors are poorly understood. Johansson et al. recently investigated the potential glycoconjugate receptors by binding A. baumannii to various glycosphingolipids on thin-layer chromatograms [104]. Collectively, three predominant glycosphingolipid receptors were isolated and identified, neolactotetraosylceramide, lactotetraosylceramide, and lactotriaosylceramide, indicating that N-acetylglucosamine (GlcNAc) was the basic recognition pattern [104].
5.2. Cytotoxins
The envelope of A. baumannii, particularly lipooligosaccharides (LOS), contributes to its cytotoxicity towards host cells. LOS is the major component on the outer leaflet of the outer membrane [105] and contributes to both immunogenicity and cytotoxicity of A. baumannii. Erridge et al. showed that A. baumannii LOS stimulates TNFs and IL-8 secretion from macrophages [106]. Several studies have demonstrated that the absence or disruption of any of the first three enzymes in the lipid A biosynthetic pathway, acyltransferase (LpxA), deacetylase (LpxC), and N-acyltransferase (LpxD), results in less-lethal infections and reduced serum survival [107,108,109,110], highlighting the immunogenicity of A. baumannii LOS.
As previously mentioned, several envelope proteins including OmpA, Omp33–36, and Ata, also contribute to cytotoxicity. Bacterial γ-glutamyl transferase enzyme (GGT), secreted via the type II secretion system, was also reported to cause cell damage directly by caspase activation and apoptosis, depletion of ATP and subsequent necrosis, and cell-cycle arrest. A high level of serum GGT was associated with oxidative stress and the exacerbation of chronic obstructive pulmonary disease (COPD) [111,112]. Very recently, Elhossieny et al. characterised A. baumannii GGT and showed A. baumannii strains with higher extracellular GGT activity resulted in more severe tissue damage by inducing increased inflammation and oxidation activities with elevated phenoloxidase, lysozyme, lactate dehydrogenase, and lipid peroxidation observed [113].
There are additional virulence factors whose roles and molecular interactions are not fully understood, but their absence has resulted in impaired virulence or lethality. For example, Smith et al. reported a type IV secretion system may contribute to the pathogenesis of A. baumannii based on its identification within pathogenicity islands (PAI) after whole-genome sequencing, but its specific roles have not been studied to date [12]. Further, it has been suggested that A. baumannii may utilise a type VI secretion system to kill competing bacteria [114].
5.3. Persistence
In addition to adhesion, invasion, and cytotoxicity contributing to A. baumannii pathogenesis, serum persistence also affects the infection process indirectly. Capsular exopolysaccharide (CPS) is an envelope component mediating the persistence of many invasive bacteria, enabling organisms to survive better under unfavourable conditions, e.g., desiccation. It also effectively protects pathogens from phagocytosis and humoral immune attack and inhibits the activation of the alternative complement pathway [115]. Many clinical isolates of A. baumannii express the K locus gene cluster, responsible for the biosynthesis and export of CPS [105,116]. However, CPS is not usually, in itself, a pathogenicity determinant as many organisms possessing polysaccharide capsules do not cause human diseases. Russo et al. identified two capsule associated genes, ptk and epsA, that were predicted to encode a putative protein tyrosine kinase (PTK) and a putative polysaccharide export OMP (EpsA), respectively, and the ptk and epsA mutants showed significantly decreased survival in soft tissue infection sites [117]. Similarly, pglC and pglL mutants showed impaired survival in mouse septicaemia models [118,119], suggesting that CPS may act more as a protectin, maintaining serum resistance during the infection process, rather than cytotoxin.
LOS may be important for A. baumannii serum resistance, conferring a competitive advantage in vivo, as mutants lacking LpsB, a highly conserved glycosyltransferase involved in LOS synthesis, showed reduced resistance to human serum [107]. Metal acquisition systems, such as iron (BasD, BauA, NfuA), zinc (ZnuABC, ZigA), and manganese (MumC, MumT) acquisition systems also contribute to in vivo survival [101,120,121,122,123]. Koenigs et al. identified a novel plasminogen binding protein, CipA, the absence of which resulted in the efficient killing of A. baumannii by human serum and impaired its ability to penetrate the endothelial monolayers [124]. Universal stress protein A (UspA) has also been documented to play a role in protecting A. baumannii from H2O2, low pH, and 2,4-DNP, and the uspA mutant formed fewer colonies in sepsis or pneumonia mouse models [125,126,127]. The capacity of A. baumannii to confer resistance to and degrade H2O2 is also largely dependent on the catalases KatE and KatG, which may attenuate the production of reactive oxygen species (ROS) by phagocytic cells of the innate immune system [128]. Finally, OmpA, phospholipase, BfmR/S system, Tuf, and RecA, have also been shown to contribute to persistence [129].

Table 1.
Identified A. baumannii Virulence Factors and Associated Functions.
Table 1.
Identified A. baumannii Virulence Factors and Associated Functions.
| Virulence Factors | Functions Involved | Adhesion | Invasion | Cytotoxicity | Ref |
|---|---|---|---|---|---|
| Ata | type Vc secretion system | + | + | + | [90,91] |
| Bap | type I secretion system | + | [88] | ||
| BasD, BauA | iron acquisition system | + | + | [130] | |
| BfmR/S | two-component regulatory system | + | + | [93,94] | |
| BLP-1, BLP-2 | Bap like protein | + | [131] | ||
| Capsular polysaccharides | outer membrane component | + | [117] | ||
| CdiA/CdiB | type Vb secretion system | + | [132] | ||
| ChoP | phosphorylcholine | + | + | [103] | |
| CipA | plasminogen binding protein | + | [124] | ||
| FhaB/FhaC | type Vb secretion system | + | [89] | ||
| GGT | type II secretion system | + | + | [113] | |
| LOS | outer membrane component | + | [106,107] | ||
| LpsB | LOS production | + | [107] | ||
| LpxA, LpxC, LpxD | LOS production | + | [107,108,109,110] | ||
| NfuA | iron acquisition system | + | + | [101,120,122] | |
| OmpA | porin | + | + | + | [82,83] |
| Omp33-36 | porin | + | + | + | [86,87] |
| paaE | production of toxic epoxide compounds | + | [129,133] | ||
| PLC, PLD | phospholipase | + | [100,101,102] | ||
| TrxA | type 4 pili production | + | [98] | ||
| Type 4 pili | motility apparatus | + | [96] | ||
| T6SS | type VI secretion system | + | + | [114,134] |
6. Anti-A. baumannii Vaccine Development
With the extensive spread of MDR A. baumannii worldwide and increasingly frequent reports of emerging PDR strains, more effective control implications are urgently needed to combat this imminent threat. Thus, novel antibiotics and anti-A. baumannii vaccines are the two main routes being followed, with the latter being especially important in view of the likelihood of the development of resistance to new antibiotics. A. baumannii has an intracellular lifestyle, invading host cells via a zipper-like mechanism [82] and therefore A. baumannii vaccines should induce acquired cellular immunity with long-term memory [135], while those that can elicit both humoral and cellular immune memory are expected to be optimal. Over a decade ago, McConnell et al. pioneered their development [136,137,138], and currently, several laboratories around the world focus on developing A. baumannii vaccines for targeted populations.
6.1. Target Populations
As an opportunistic pathogen, A. baumannii rarely causes diseases in healthy people, nevertheless, protection is needed for vulnerable individuals, particularly those residing in, or frequently travelling to, areas with a high prevalence or historic outbreaks of A. baumannii. These susceptible cohorts are the main target population of A. baumannii vaccines and were nominally identified and characterised into three groups (Table 2) based on reported epidemiology studies.
Table 2.
Target Populations of A. baumannii Vaccines.
6.2. Vaccine Candidates
Scientists have been pursuing an effective vaccine against A. baumannii for the past decade and several antigens and strategies have been examined in preclinical studies.
6.2.1. Whole-Cell Vaccines
Given that a considerable proportion of the individuals susceptible to A. baumannii are those with compromised health conditions, there are perceived potential safety risks with administering live-attenuated vaccines to vulnerable cohorts. Consequently, to our knowledge, only one attenuated whole-cell A. baumannii vaccine candidate has been reported to date which comprises a genetically modified, attenuated MDR clinical strain deficient in the TrxA adhesin (Table 3). Mice immunised via intraperitoneal (i.p.) or subcutaneous (s.c.) routes were protected by 100% and 90% respectively against a ten-fold lethal dose (LD50) challenge for a period of four weeks post-challenge. Moreover, immunisation-induced antisera provided fully effective protection in passive immunisation experiments, highlighting that this attenuated live vaccine induced robust humoral responses despite minimal splenic cell-mediated immunity [139].
Several inactivated whole-cell vaccine candidates formulated either with or without adjuvants were also evaluated [137,140,141,142]. The first reported vaccine candidate against A. baumannii was a formalin-killed ATCC 19606 whole-cell vaccine administered with aluminium phosphate adjuvant. Mice intramuscularly (i.m.) immunised with this vaccine were effectively protected from a lethal sepsis challenge raised by heterologous clinical strains, including one PDR strain, with seven-day survival rates between 85%–100%. Significant increases in the serum levels of immunoglobulin G1 (IgG1), IgG2a, and IgM were detected, suggesting a mixed Th1/Th2-mediated response was stimulated [137]. Interestingly, while sepsis is the most frequently used challenge model, KuoLee et al. first reported an experimental vaccine for protection against respiratory challenge, to test a candidate administered via mucosal route. This intranasal (i.n.) immunisation with formalin-killed cells of a hypervirulent A. baumannii strain LAC-4 successfully elicited both mucosal and humoral immune responses in mice with significant levels of IgA, IgG1, and IgG2a in bronchoalveolar lavage (BAL) fluid being achieved. All immunised mice were completely protected from lethal i.n. or i.p. challenges raised by the LAC-4 strain for up to 10 days [141].
6.2.2. Subunit Vaccines
As novel virulence factors of A. baumannii and their molecular interactions continue to be identified in the past decade, their immunogenicity and their potential as subunit vaccines have been more extensively investigated (Table 3). Outer membrane immunogenic proteins functioning as adhesins or invasins are the main targets for vaccine development because they represent the initial stages of infection, namely bacterial attachment to host cell receptors and colonisation, and blocking this may be the most effective strategy to prevent bacterial infections [143].
Probably the most understood and promising subunit candidate for A. baumannii vaccines is OmpA because, as previously discussed, it is an all-round player involved in almost every critical step of A. baumannii pathogenesis, including adhesion, invasion, cytotoxicity, and also in vivo persistence, and is highly immunogenic in animal models. Elucidation of the native structure of A. baumannii OmpA showed that its amino acid sequence has no homology to the human proteome but shows a low variation (≥89% conserved) across clinical isolates [144,145], which indicates that A. baumannii OmpA would be less likely to cause mutual interference with other OmpA-containing commensal bacteria, minimising the potential side effects as a vaccine. Several studies have presented positive results demonstrating active immunisation with OmpA-based vaccines could confer prophylactic benefits against A. baumannii (Table 3). Intranasally immunised rOmpA conjugated to cholera toxin, showed 15-day survival rates from 40% to 100% following i.p. challenges compared with 100% mortality in two days in the control groups [146]. A study in older (>6 months) diabetic mice demonstrated protection of 50% for up to 28 days following rOmpA vaccination which was equivalent to that in the juvenile group [147] suggesting that OmpA-based vaccine protection may translate across a broad age range and include people with diabetes. The dose of antigen administered may impact the profile of the immune response with low dose (3 μg) rOmpA immunised mice showing a balanced IFN-γ and IL-4 immune responses, while mice that received a higher dose (100 μg) showed a polarised Th2-response as demonstrated by IL-4 secretion [148].
Besides OmpA, several other virulence factors have also been tested as prospective vaccine candidates. Omp33–36 was specifically recognised by IgG, IgM, and IgA from A. baumannii colonised patients and showed no cross-reaction with sera from other infections, [149], highlighting its potential as a vaccine antigen. In addition, this highly conserved adhesin is present in over 1600 strains of A. baumannii with ≥98% identity [150]. Omp33–36 is considered as a suitable immunogen to protect against A. baumannii and its co-administration with OmpA may confer even greater protection; however, data from active immunisation studies are currently insufficient. In silico simulation also predicted peptides constructed by exposed epitopes of Omp33–36 could trigger antibody responses with higher avidity [150]. Alternatively, Bentancor et al. reported the passive administration of anti-Ata rabbit sera protected both immunocompetent and immunocompromised mice from A. baumannii-induced pneumonia by engendering anti-adhesive and robust opsonic activities [151], making Ata a potential vaccine antigen. Although Ata has been identified in many clinical isolates, the variable expression of Ata among clinical isolates suggests that it may not provide adequate coverage [17] as a monovalent vaccine. CPS PNAG and K1 were also suggested as targets for protective immunity against A. baumannii infections [152,153] but their efficacy, when conjugated with carriers, has not been studied yet. Recombinant Bap completely protected immunised mice from sepsis following a lethal challenge, and when mice were challenged at the extreme dose of 106 × LD50 (1013 CFU), a five-day survival rate of 60% was obtained [154]. It should be noted that the use of Freund’s adjuvant (FA) as adjuvant undermines the relevance of this response, given its reactogenicity. Two siderophore receptors in A. baumannii, BauA, and BfnH, proved only moderately effective against lethal sepsis challenges with survival rates of 40% and 44.5%, respectively [155].
Table 3.
Preclinical active immunisation studies of A. baumannii vaccine candidates.
Table 3.
Preclinical active immunisation studies of A. baumannii vaccine candidates.
| Platform and Immunogen | Mice Strain | Immunisation Schedule | In Vivo Challenge Study | Immune Responses | Ref. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Day | Route | Dose | Chall. Strain(s) | Model (Route) | Dose * (CFU) (LD50) | Survival (%) | Survival monitored (days post challenge) | ||||
| Live Attenuated Vaccines | |||||||||||
| Ci79 ΔtrxA | C57BL/6 (4w–6w) | 0, 14 | i.p. / s.c. | 2 × 105 CFU | WT Ci79 | day 28 sepsis (i.p.) | 5 × 106 (10 × LD50) | i.p.—100% s.c.—90% | 28 days | robust humoral responses, minimal cellular immunity | [139] |
| 1 × 106 (2 × LD50) | i.p.—100% s.c.—100% | ||||||||||
| Inactivated Vaccines | |||||||||||
| formalin-killed ATCC 19606 with AlPO4 | C57BL/6 (6w–8w) | 0, 21 | i.m. | 1 × 108 CFU | ATCC 19606 Ab-154 113-16 | day 28 sepsis (i.p.) | 1.6 × 106 (242 × LD50) 3.1 × 106 (6.5 × LD50) 4.1 × 106 (74.5 × LD50) | 100% 100% ~85% | 7 days | significant levels of IgG and IgM, Th2- and Th1-mediated responses | [137] |
| formalin-killed LAC-4 | C57BL/6 BALB/c (8w–12w) | 0, 14, 21 | i.n. | 5 × 107 CFU | LAC-4 | day 42 sepsis (i.p.) pneumonia (i.n.) | 8.4 × 105 (i.p.) 7 × 107 (i.n.) | 100% | 10 days | mucosal immune responses, dominant Th2 responses | [141] |
| formalin-killed IB010 (ATCC 19606 ΔlpxD) with AlPO4 | C57BL/6 (6w–8w) | 0, 14 | i.m. | 1 × 109 CFU | ATCC 19606 Ab-154 | day 21 sepsis (i.p.) | 2.3 × 106 (341 × LD50) 1.1 × 106 (2.2 × LD50) | 100% 100% | 7 days | robust IgG1 and IgG2c responses | [142] |
| Subunit Vaccines | |||||||||||
| rOmpA (ATCC 19606) conjugated with cholera toxin | BALB/c (6w–8w) | 0, 21 | i.n. | 10 μg | strain A strain B strain C strain D strain E | day 28 sepsis (i.p.) | 5 × 108 (2.5 × LD50) 2 × 108 (2 × LD50) 5 × 107 (2 × LD50) 3 × 108 (3 × LD50) 3 × 107 (3 × LD50) | ~50% ~40% ~55% 100% ~70% | 15 days | significant levels of IgG and IgA | [146] |
| rOmpA (ATCC 17978) with Al(OH)3 | diabetic BALB/c (>6 m and 6w–10w) | 0, 21 | s.c. | 3 μg | HUMC 1 | day 35 sepsis (i.v.) | 2 × 107 | ~50% | 28 days | robust humoral responses | [147] |
| rBap (Kh0060) with FA | BALB/c (4w–6w) | 0, 14,28 | not stated | 10 μg | Kh0060 | day 35 sepsis (i.p.) | 1 × 109 (100 × LD50) 1 × 1011 (104 × LD50) 1 × 1013 (106 × LD50) | 100% 80% 60% | 5 days | robust humoral responses | [154] |
| rBauA or rBfnH (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,28 | s.c. | 20 μg | ATCC 19606 | day 49 sepsis (i.p.) | 1.5 × 107 (2 × LD50) | ~40% | 7 days | not specified | [155] |
| rBamA (ATCC 19606) with Al(OH)3 | BALB/c (6w–8w) | 0, 14,28 | i.p. | 20 μg | P-562 | day 45 pneumonia (i.n.) | 1 × 109 | ~80% | 7 days | non-neutralising, opsonic IgG | [156] |
| rBamA (ATCC 19606] with FA | BALB/c (6w–8w] | 0, 14, 28, 42 | s.c. | 20 μg | ATCC 19606 | day unknown, sepsis (i.p.) | 2 × 106 (4 × LD50) 7 × 106 (14 × LD50) | 100% 25% | 4 days | not specified | [157] |
| rTrx-OmpW (ATCC 17978) with alum | ICR (6w–8w) | 0, 14,28 | s.c. | 50 μg | ATCC 17978 | day 49 sepsis (i.p.) | 1 × 106 | 100% | 7 days | opsonic IgG, complements | [158] |
| rTrx-Omp22 (ATCC 17978) | ICR (6w–8w) | 0, 14, 28 | s.c. | 50 μg 20 μg 10 μg 5 μg | Ab1 | day 49 sepsis (i.p.) | 1 × 106 | 100% 33% 33% 0 | 7 days | opsonic IgG, complements | [159] |
| rOmp22 (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 37.5% | 8 days | not specified | [160] |
| rOmpK (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 25% | 8 days | not specified | [160] |
| rOmp22-OmpK (ATCC 19606) with FA | BALB/c (6w–8w) | 0, 14,21 | s.c. | 20 μg | ATCC 19606 | day 42 sepsis (i.p.) | 2 × 108 | 67% | 8 days | not specified | [160] |
| rOmp22-OmpK (ATCC 19606) with MF59 adjuvant | BALB/c (6w–8w) | 0, 14,21 | i.t. | 30 μg | ATCC 19606 | day 42 respiratory (i.t.) | 1 × 108 | 83.3% | 10 days | significant levels of IgG and IgA | [161] |
| Multi-Component Vaccines | |||||||||||
| OMC (ATCC 19606) with AlPO4 | C57BL/6 (6w–8w) | 0, 21 | i.m. | 25 μg | ATCC 19606 Ab-154 113-16 | day 35 sepsis (i.p.) | 1 × 106 (151.3 × LD50) 1 × 107 (20.9 × LD 50) 1.5 × 106 (27.3 × LD50) | 100% 80% 100% | 7 days | robust IgG and IgM responses | [136] |
| OMV (ATCC 19606) with AlPO4 | C57BL/6 (6w–8w) | 0, 14 | i.m. | 10 μg | ATCC 19606 Ab-154 113-16 | day 35 sepsis (i.p.) | 4.5 × 105 (71.3 × LD50) 1.7 × 106 (4.3 × LD50) 2 × 106 (39.9 × LD50) | 100% ~90% 100% | 7 days | robust IgG1, IgG2c and IgM responses | [138] |
| ClyA-Omp22 (ATCC 17978) with E. coli derived OMV | ICR (6w–8w) | 0, 14 | s.c. | 5 μg 10 μg 20 μg 50 μg | Ab1 | day 35 sepsis (i.p.) | 1 × 106 | 30% 40% 63.6% 100% | 7 days | opsonic IgG | [162] |
| Nucleic Acid Vaccines | |||||||||||
| pVAX1-ompA (LAC-4) with CpG-ODN adjuvant | C57BL/6 (6w–8w) | 0, 14, 21 | i.m. | 100 μg | LAC-4 SJZ04 SJZ18 SJZ28 | day 28 respiratory (i.t.) | 1.2 × 107 (2 × LD50) 1 × 108 1 × 108 1 × 108 | ~50% ~40% ~60% ~50% | 7 days | robust humoral responses, mixed Th1/Th2/Th17 cellular responses | [163] |
| pBudCE4.1-ompA | BALB/c (6w–8w) | 0, 7, 14, 28 | i.m. | 25 μg | not stated | day 35 respiratory (i.n.) | 1 × 108 | 60% | 15 days | significant levels of IgG and IgM | [164] |
Abbreviations: i.m., intramuscular; i.n., intranasal; i.t., intratracheal; i.p., intraperitoneal; s.c., subcutaneous. * Dose includes LD50, when reported.
Additionally, some OMPs which are less relevant to A. baumannii pathogenesis and overlooked as virulence factors may also have a potential impact as vaccines. These immunogens are likely to be safer even when a high dose is given [159] and have been shown to be efficacious antigens in other infections [165]. Purified renatured OMP assembly factor BamA protected mice against a lethal pneumonia challenge with a seven-day survival rate of 80% and 60%, respectively following active and passive immunisation. The opsonophagocytic killing assay showed the immunisation elicited a high titre of non-neutralising, opsonising antibodies [156]. The carbapenem resistance OMP (CarO) was also predicted to contain structurally important linear B cell epitopes based on bioinformatic design [166]. OmpW is an A. baumannii porin involved in iron uptake and colistin binding which is conserved across 804 strains with more than 91% identity in protein sequence [167]. We have previously reported that the Burkholderia pseudomallei OmpW homolog afforded extended protection (81 days) against a lethal melioidosis challenge [165]. Immunisation with a recombinant Trx–OmpW fusion protein fully protected mice from sepsis challenge for up to seven days. The antisera demonstrated significant opsonophagocytic activities against homologous strains and clonally distinct clinical isolates in vitro [158]. Later, the same research group identified a novel OMP with a molecular weight of 22 kDa, namely Omp22, highly conserved (95% identity in amino acid sequence) in 851 reported A. baumannii strains and with almost negligible homology to human proteins. Again, a recombinant Trx–Omp22 fusion protein showed 100% protection of immunised mice for seven days after lethal challenge at high doses, with lower doses showing protection of a third of the mice [159]. Although Omp22 has immunogenic and prophylactic properties, it may not be sufficiently potent to invoke adequate protection unless being administered at a high dose. Fusion of Omp22 with another weak antigen, OmpK, improved the protection against A. baumannii to a higher level than those immunised with either of the two proteins individually [160,161] demonstrating the utility of a multivalent vaccine approach.
6.2.3. LOS and Conjugate Vaccines
A rather controversial and neglected antigen for subunit vaccine candidates is LOS (Section 5.2). OMPs extracted from A. baumannii cells or recombinant A. baumannii proteins produced by E. coli customarily require a detoxification step reducing the content of LOS to a minimal level to weaken the endotoxin activity, ensuring vaccine safety, and critically ensuring that the immunological response is mediated by the protein antigen being evaluated. LOS is also not a popular target for vaccine development due to the short-lasting B cell memory it induces; however, mice vaccinated with LOS alone were protected from a lethal challenge [142]. LOS is recognised by TLRs, particularly TLR-4, and safer derivatives may be feasible adjuvants. This has been supported by a study where LOS-free OMPs developed from an A. baumannii mutant deficient in LOS production showed significantly reduced protection (40%) compared with those derived from the wild-type cells, and administration of exogenous LOS restored the survival levels [142]. Noticeably, García-Quintanilla et al. reported that A. baumannii can acquire resistance to colistin and collateral sensitivity to azithromycin, rifampicin, and vancomycin through the complete loss of LOS, suggesting that LOS-based vaccines may deliver weakened protection against some problematic LOS-deficient A. baumannii MDR or PDR strains [142]. Despite this, although it may not be applicable as an immunogen, LOS (or safer derivatives) represent potentially powerful adjuvants boosting protective immune responses, and its usage as well as potential risks should be evaluated more comprehensively.
6.2.4. Multi-Component Vaccines
Unfortunately, despite encouraging results in pre-clinical studies, single subunit antigens are unlikely to elicit comprehensive protection in humans against heterologous A. baumannii strains. To date, single recombinant protein vaccines have performed poorly in providing protection against bacteria that mediate pathogenesis via multiple virulence mechanisms. A. baumannii expresses several virulence factors that allow successful evasion of innate immune defence [168], contributing to the challenge of selecting appropriate antigens. Vaccines containing multiple antigens will display a higher density of epitopes with enriched diversity and are expected to exhibit increased immunogenicity, bringing better protection efficiency. Multi-component vaccines against A. baumannii are insufficiently studied and most of them are outer member complex (OMC) or outer membrane vesicle (OMV) based.
OMPs extracted from A. baumannii reference strain ATCC 19606 effectively elicited both humoral and cellular responses in vaccinated mice and induced 80% to 100% protection against sepsis following challenge with heterologous A. baumannii isolates. Notably, the antisera elicited with this OMC vaccine also cleared established A. baumannii infections (70–100% recovery rate) [136]. However, a drawback of these preparations is that the composition can be ill-defined, leading to major concerns on safety, regulatory compliance, and feasibility for consistent industrial production.
Despite this, the use of naturally secreted outer membrane vesicles (OMV) as vaccine technology platforms has gained renewed interest in recent years particularly since the approval of meningococcal group B vaccine, Bexsero®, an OMV-based product [169]. Crucially they present a wider variety of antigens, including both OMPs and other membrane-associated proteins, and can be further designed to package and deliver antigens. Consequently, with optimal bioengineering design, isolation, LOS detoxification, OMVs could be developed into safe, effective, and cost-effective vaccines against bacterial infections. This topic is covered in two excellent reviews [169,170]. Natural OMVs produced by the A. baumannii reference strain ATCC 19606 proved highly effective in immunising mice, and 90% of the immunised mice were protected against heterologous strains. Subsequent MALDI–TOF MS analysis identified six proteins with high immunogenicity in the OMVs, including previously mentioned, OmpA, OmpW, Omp33–36, and CarO, as well as two other putative proteins [138] which were likely critical to the protective response. In a later study, Huang et al. successfully engineered E. coli-derived OMVs packaging fused proteins of exogenous A. baumannii Omp22 and a pore-forming haemolytic protein ClyA, which protected over 60% and 100% of mice against a sepsis challenge, without any additional adjuvants. It is worth noting that the carrier control groups administered unmodified E. coli OMVs showed 35–63.6% survival indicating that E. coli-derived OMVs elicited cross-immunity to A. baumannii infection [162].
6.2.5. Nucleic Acid Vaccines
Despite nucleic acid vaccines have gained increasing interest as vaccine platforms in recent years, there are barely any studies on nucleic acid vaccines against A. baumannii. Very recently, DNA vaccines delivering A. baumannii OmpA gene were reported to induce a mixed humoral and cellular response protecting lethal bacterial challenges in murine pneumonia models [163,164]. Recently, a pVAX1 vector encoding A. baumannii OmpA and proteoglycan associated lipoprotein (PAL) genes administered intramuscularly resulted in the protection of >80% of immunised mice to lethal pulmonary challenge with four heterologous strains, while immunisation with an OmpA encoding vector elicited 50% survival in response to challenge. A high level of humoral responses and mixed Th1/Th2/Th17 cellular responses, and reduced bacterial loads were reported with reduced inflammatory cytokines and inflammatory cell infiltration in the BAL [163]. Immunisation with an alternative plasmid-encoded-ompA vaccine utilising a eukaryotic expression vector pBudCE4.1 protected 60% of the immunised mice against pulmonary infection for up to 15 days post-challenge, eliciting moderate Il-2, IL-4, Il-12, and IFN-γ responses [164]. These nucleic vaccine approaches are still at an early stage but hold promise as safe cost-effective methods of administering multivalent vaccines.
6.3. Lessons from A. baumannii Vaccines and Future Directions
Despite the aforementioned efforts, the progress of vaccines against A. baumannii has lagged behind that for nosocomial pathogens such as C. difficile, P. aeruginosa, and S. aureus [9], and to date no candidate has entered clinical trials, suggesting the challenges in developing safe and efficacious A. baumannii vaccines.
6.3.1. Modern Vaccine Technologies
Current investigations on anti-A. baumannii vaccines are still in their infancy, with most research efforts looking into conventional approaches and platforms, e.g., subunit and killed vaccines. Novel antigen carriers and advanced delivery systems, such as nucleic acid vaccines, virus vectors, conjugated carriers, and co-delivery of multiple antigens are rarely explored. Nevertheless, as the mRNA and DNA vaccine technologies have been acknowledged throughout the world during the COVID-19 vaccine race, it is foreseeable that burgeoning attention may be paid to nucleic acid vaccines against A. baumannii and other pathogens in the coming future. In addition, proteomic approaches and reverse vaccinology exploit modern bioinformatics, allowing the systematic in silico evaluation and selection of putative immunogens for vaccine candidates, which may have a wider application for future antigen selection and vaccine design against A. baumannii infections.
6.3.2. Long-Term Cellular Immunity
In pre-clinical trials, a promising vaccine candidate against A. baumannii should demonstrate its protective benefits in immunised animals with a significantly improved survival rate, reduced bacterial burdens within organs, and the suppressed accumulation of inflammatory cytokines and chemokines in sera. Furthermore, the resulting anti-sera are expected to contain sufficient antibody titre for effective passive immunisation and even for antibody therapy treating established infections. Therefore, most studies have laid emphasis on evaluating post-immunisation or post-infection humoral immune responses, while cell-mediated immunity remains largely unstudied. However, as an invasive pathogen, it is essential that A. baumannii vaccines elicit dominant cellular immune responses and prime sustained T memory cells for long-term protection. In this context, though most studies reported robust vaccine-induced humoral or Th2-mediated immune responses, their performance in stimulating cellular immunity was not specified. Besides, the intervals between the last immunisation and bacterial challenge were only one week in some studies, making the evaluation of the endurance of the protective response even less reliable. It is hence important for the A. baumannii vaccine research community to standardise the evaluation protocols, such as the animal models, intervals between immunisation and challenges, challenge strains, doses, and routes, so that the comparison in the efficacy of the experimental vaccines between different laboratories is more rigorous [9].
6.3.3. Multivalent Protection
It is encouraging to note that many A. baumannii vaccine studies have evaluated the protection against not only homologous strain challenges but also challenges raised by other clinical isolates to demonstrate the broad protection offered by the vaccine candidates. However, as the protection rates reported in these studies varied considerably with strain, immunisation, and/or challenging dose, their protective effects require more comprehensive evaluations covering a broader range of heterologous strains in diverse challenging scenarios, especially for those involving highly virulent strains and massive exposure dose. Strain-dependent diversity in the selected antigens may also increase the challenges in developing a vaccine providing broad or multivalent protection, as even in a specific bacterial strain, the amino acid sequences of some antigen proteins, e.g., OmpA, may vary among multiple subclasses [145,171]. At a high level, A. baumannii has an exceptional ability to acquire foreign DNA and generate new variants as the whole genome sequencing of ATCC 179878 found a significant fraction of open reading frames (ORF) (17.2%) in 28 putative alien islands [12], and such multiplicity of strains makes repeated infections possible and complicates vaccine development. Therefore, choosing conserved antigens and developing multi-component vaccines will be critical to effective protection.
7. Conclusions
The increasing attention to A. baumannii arose following outbreaks in the US army during the Iraq conflicts, and since then is becoming a global concern. Currently available treatment options for MDR A. baumannii-caused infections are quite limited where colistin is often reported as the only antibiotic delivering therapeutic effects, making the pathogen extremely difficult to treat. Massive research efforts have been devoted to its antibiotic resistance mechanisms and epidemiology but so far, our understanding of its pathology is comparably limited. In this context, vaccines against A. baumannii infections, are urgently needed and will be of great value in the prevention of infection and control of this canny pathogen. The past decade has witnessed the very first efforts in A. baumannii vaccine development, however, none have entered clinical trials, which is clear evidence of the challenges involved and the need for further progress in understanding the complex A. baumannii–host relationships at each stage of the infection process. Most A. baumannii vaccine candidates developed to date explored conventional protein-based technologies, inducing opsonophagocytic antibody-mediated killing and/or antibody-mediated toxin inhibition. However, a polarised Th2 response may not be sufficient for effective and long-term protection, considering that A. baumannii is an invasive pathogen; consequently, a mixed Th1/Th2 orTh1/Th17 response may be more beneficial. Investigations of novel platforms, as well as adjuvant development, could offer the possibility of optimising immune mechanisms and targeting multiple antigens, which will eventually deliver vaccines with broad coverage and clinical efficacy. In addition, the COVID vaccine and associated technology acceleration could change the landscape of future vaccine research and development. With the licensing of the first mRNA vaccines, nucleic acid vaccines against A. baumannii and other invasive pathogens are expected to generate more interest in the near future. There is some urgency, however, in order to tackle the global spread of MDR A. baumannii, and vaccine development needs to accelerate in order to keep up with the pace of spread.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/vaccines9060570/s1, Table S1: Type strains for the phylogenetic analysis of Acb complex, Table S2: Worldwide data for percentage of carbapenem-resistant A. baumannii (R%), Table S3: European data (2018) for R% and F%.
Author Contributions
Conceptualisation, C.M. and S.M.; writing—original draft preparation, C.M.; writing—review and editing, S.M.; visualisation, C.M. Both authors have read and agreed to the published version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
C.M. holds a China Scholarship Council studentship with University College Dublin under grant number CSC202008300014. S.McC. is in receipt of funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No 860325.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study is contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef]
- Antunes, L.C.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Bulens, S.N.; Yi, S.H.; Walters, M.S.; Jacob, J.T.; Bower, C.; Reno, J.; Wilson, L.; Vaeth, E.; Bamberg, W.; Janelle, S.J.; et al. Carbapenem-nonsusceptible Acinetobacter baumannii, 8 US metropolitan areas, 2012–2015. Emerg. Infect. Dis. 2018, 24, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Inchai, J.; Pothirat, C.; Bumroongkit, C.; Limsukon, A.; Khositsakulchai, W.; Liwsrisakun, C. Prognostic factors associated with mortality of drug-resistant Acinetobacter baumannii ventilator-associated pneumonia. J. Intensive Care 2015, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Wisplinghoff, H.; Paulus, T.; Lugenheim, M.; Stefanik, D.; Higgins, P.G.; Edmond, M.B.; Wenzel, R.P.; Seifert, H. Nosocomial bloodstream infections due to Acinetobacter baumannii, Acinetobacter pittii and Acinetobacter nosocomialis in the United States. J. Infect. 2012, 64, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Metan, G.; Alp, E.; Aygen, B.; Sumerkan, B. Acinetobacter baumannii meningitis in post-neurosurgical patients: Clinical outcome and impact of carbapenem resistance. J. Antimicrob. Chemother. 2007, 60, 197–199. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Wendt, C.; Dietze, B.; Dietz, E.; Rüden, H. Survival of Acinetobacter baumannii on dry surfaces. J. Clin. Microbiol. 1997, 35, 1394–1397. [Google Scholar] [CrossRef]
- Chen, W. Current advances and challenges in the development of Acinetobacter vaccines. Hum. Vaccines Immunother. 2015, 11, 2495–2500. [Google Scholar] [CrossRef]
- Weiss, N.; Kracht, M.; Gleim, D. List of Prokaryotic Names with Standing in Nomenclature-Genus Acinetobacter. 2021. Available online: https://lpsn.dsmz.de/genus/acinetobacter (accessed on 24 May 2021).
- Rossau, R.; Van Landschoot, A.; Gillis, M.; De Ley, J. Taxonomy of Moraxellaceae fam. nov., a new bacterial family to accommodate the genera Moraxella, Acinetobacter, and Psychrobacter and related organisms. Int. J. Syst. Bacteriol. 1991, 41, 310–319. [Google Scholar] [CrossRef]
- Smith, M.G.; Gianoulis, T.A.; Pukatzki, S.; Mekalanos, J.J.; Ornston, L.N.; Gerstein, M.; Snyder, M. New insights into Acinetobacter baumannii pathogenesis revealed by high-density pyrosequencing and transposon mutagenesis. Genes Dev. 2007, 21, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus—Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [CrossRef]
- Nemec, A.; Krizova, L.; Maixnerova, M.; Sedo, O.; Brisse, S.; Higgins, P.G. Acinetobacter seifertii sp. nov., a member of the Acinetobacter calcoaceticus–Acinetobacter baumannii complex isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2015, 65, 934–942. [Google Scholar] [CrossRef]
- Cosgaya, C.; Marí-Almirall, A.; Van Assche, A.; Fernández-Orth, D.; Mosqueda, N.; Telli, M.; Huys, G.; Higgins, P.G.; Seifert, H.; Lievens, B.; et al. Acinetobacter dijkshoorniae sp. nov., a new member of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex mainly recovered from clinical samples in different countries. Int. J. Syst. Evol. Microbiol. 2016, 66, 4105–4111. [Google Scholar] [CrossRef]
- Rooney, A.; Dunlap, C.; Flor-Weiler, L.B. Acinetobacter lactucae sp. nov., isolated from iceberg lettuce (Asteraceae: Lactuca sativa). Int. J. Syst. Evol. Microbiol. 2016, 66, 3566–3572. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Genet. 2018, 16, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Boo, T.W.; Walsh, F.; Crowley, B. Molecular characterization of carbapenem-resistant Acinetobacter species in an Irish university hospital: Predominance of Acinetobacter genomic species 3. J. Med. Microbiol. 2009, 58, 209–216. [Google Scholar] [CrossRef]
- Zhang, H.Z.; Zhang, J.S.; Qiao, L. The Acinetobacter baumannii group: A systemic review. World J. Emerg. Med. 2013, 4, 169–174. [Google Scholar] [CrossRef]
- Lee, Y.C.; Huang, Y.T.; Tan, C.K.; Kuo, Y.W.; Liao, C.H.; Lee, P.I.; Hsueh, P.-R. Acinetobacter baumannii and Acinetobacter genospecies 13TU and 3 bacteraemia: Comparison of clinical features, prognostic factors and outcomes. J. Antimicrob. Chemother. 2011, 66, 1839–1846. [Google Scholar] [CrossRef]
- Kang, Y.S.; Jung, J.; Jeon, C.O.; Park, W. Acinetobacter oleivorans sp. nov. is capable of adhering to and growing on diesel-oil. J. Microbiol. 2011, 49, 29–34. [Google Scholar] [CrossRef]
- Hare, J.M.; Bradley, J.A.; Lin, C.L.; Elam, T.J. Diverse responses to UV light exposure in Acinetobacter include the capacity for DNA damage-induced mutagenesis in the opportunistic pathogens Acinetobacter baumannii and Acinetobacter ursingii. Microbiology 2012, 158, 601. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Estrada, V.; Grana-Miraglia, L.; Lopez-Leal, G.; Castillo-Ramirez, S. Phylogenomics reveals clear cases of misclassification and genus-wide phylogenetic markers for Acinetobacter. Genome Biol. Evol. 2019, 11, 2531–2541. [Google Scholar] [CrossRef] [PubMed]
- Beijerinck, M. Pigmenten als oxydatieproducten gevormd door bacterien. Versl K. Akad Wetensch Amst. 1911, 19, 1092–1103. [Google Scholar]
- Al Atrouni, A.; Joly-Guillou, M.-L.; Hamze, M.; Kempf, M. Reservoirs of non-baumannii Acinetobacter Species. Front. Microbiol. 2016, 7, 49. [Google Scholar] [CrossRef]
- Seifert, H.; Dijkshoorn, L.; Gerner-Smidt, P.; Pelzer, N.; Tjernberg, I.; Vaneechoutte, M. Distribution of Acinetobacter species on human skin: Comparison of phenotypic and genotypic identification methods. J. Clin. Microbiol. 1997, 35, 2819–2825. [Google Scholar] [CrossRef]
- La Scola, B.; Raoult, D. Acinetobacter baumannii in human body louse. Emerg. Infect. Dis. 2004, 10, 1671. [Google Scholar] [CrossRef]
- Kempf, M.; Abdissa, A.; Diatta, G.; Trape, J.-F.; Angelakis, E.; Mediannikov, O.; La Scola, B.; Raoult, D. Detection of Acinetobacter baumannii in human head and body lice from Ethiopia and identification of new genotypes. Int. J. Infect. Dis. 2012, 16, e680–e683. [Google Scholar] [CrossRef]
- Bouvresse, S.; Socolovschi, C.; Berdjane, Z.; Durand, R.; Izri, A.; Raoult, D.; Chosidow, O.; Brouqui, P. No evidence of Bartonella quintana but detection of Acinetobacter baumannii in head lice from elementary schoolchildren in Paris. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 475–477. [Google Scholar] [CrossRef]
- Fournier, P.E.; Vallenet, D.; Barbe, V.; Audic, S.; Ogata, H.; Poirel, L.; Richet, H.; Robert, C.; Mangenot, S.; Abergel, C.; et al. Comparative genomics of multidrug resistance in Acinetobacter baumannii. PLoS Genet. 2006, 2, e7. [Google Scholar] [CrossRef]
- Eveillard, M.; Kempf, M.; Belmonte, O.; Pailhories, H.; Joly-Guillou, M.L. Reservoirs of Acinetobacter baumannii out-side the hospital and potential involvement in emerging human community-acquired infections. Int. J. Infect. Dis. 2013, 17, e802–e805. [Google Scholar] [CrossRef]
- Byrne-Bailey, K.G.; Gaze, W.H.; Kay, P.; Boxall, A.B.; Hawkey, P.M.; Wellington, E.M. Prevalence of sulfonamide resistance genes in bacterial isolates from manured agricultural soils and pig slurry in the United Kingdom. Antimicrob. Agents Chemother. 2009, 53, 696–702. [Google Scholar] [CrossRef]
- Furlan, J.; Pitondo-Silva, A.; Stehling, E.G. New STs in multidrug-resistant Acinetobacter baumannii harbouring β -lactamases encoding genes isolated from Brazilian soils. J. Appl. Microbiol. 2018, 125, 506–512. [Google Scholar] [CrossRef]
- Girlich, D.; Poirel, L.; Nordmann, P. First isolation of the blaOXA-23 carbapenemase gene from an environmental Acinetobacter baumannii isolate. Antimicrob. Agents Chemother. 2009, 54, 578–579. [Google Scholar] [CrossRef]
- Higgins, P.G.; Hrenovic, J.; Seifert, H.; Dekic, S. Characterization of Acinetobacter baumannii from water and sludge line of secondary wastewater treatment plant. Water Res. 2018, 140, 261–267. [Google Scholar] [CrossRef]
- Kittinger, C.; Kirschner, A.; Lipp, M.; Baumert, R.; Mascher, F.; Farnleitner, A.H.; Zarfel, G.E. Antibiotic resistance of Acinetobacter spp. isolates from the river Danube: Susceptibility stays high. Int. J. Environ. Res. Public Health 2017, 15, 52. [Google Scholar] [CrossRef]
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.-L.; Dabboussi, F.; Kempf, M. Extrahuman epidemiology of Acinetobacter baumannii in lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef]
- Houang, E.T.; Chu, Y.; Leung, C.; Chu, K.; Berlau, J.; Ng, K.; Cheng, A. Epidemiology and infection control implications of Acinetobacter spp. in Hong Kong. J. Clin. Microbiol. 2001, 39, 228–234. [Google Scholar] [CrossRef]
- Hrenovic, J.; Durn, G.; Goic-Barisic, I.; Kovacic, A. Occurrence of an environmental Acinetobacter baumannii strain similar to a clinical isolate in paleosol from Croatia. Appl. Environ. Microbiol. 2014, 80, 2860–2866. [Google Scholar] [CrossRef] [PubMed]
- Klotz, P.; Higgins, P.G.; Schaubmar, A.R.; Failing, K.; Leidner, U.; Seifert, H.; Scheufen, S.; Semmler, T.; Ewers, C. Seasonal occurrence and carbapenem susceptibility of bovine Acinetobacter baumannii in Germany. Front. Microbiol. 2019, 10, 272. [Google Scholar] [CrossRef]
- Al Bayssari, C.; Dabboussi, F.; Hamze, M.; Rolain, J.-M. Emergence of carbapenemase-producing Pseudomonas aeruginosa and Acinetobacter baumannii in livestock animals in Lebanon. J. Antimicrob. Chemother. 2015, 70, 950–951. [Google Scholar] [CrossRef]
- Poirel, L.; Berçot, B.; Millemann, Y.; Bonnin, R.A.; Pannaux, G.; Nordmann, P. Carbapenemase-producing Acinetobacter spp. in cattle, France. Emerg. Infect. Dis. 2012, 18, 523. [Google Scholar] [CrossRef]
- Wilharm, G.; Skiebe, E.; Higgins, P.G.; Poppel, M.T.; Blaschke, U.; Leser, S.; Heider, C.; Heindorf, M.; Brauner, P.; Jäckel, U.; et al. Relatedness of wildlife and livestock avian isolates of the nosocomial pathogen Acinetobacter baumannii to lineages spread in hospitals worldwide. Environ. Microbiol. 2017, 19, 4349–4364. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, A.; Findlay, J.; Al Hassan, L.; Amyes, S.G. Epidemiology of Acinetobacter baumannii of animal origin. Int. J. Antimicrob. Agents 2011, 38, 314–318. [Google Scholar] [CrossRef]
- Łopińska, A.; Indykiewicz, P.; Skiebe, E.; Pfeifer, Y.; Trček, J.; Jerzak, L.; Minias, P.; Nowakowski, J.; Ledwoń, M.; Betleja, J.; et al. Low occurrence of Acinetobacter baumannii in gulls and songbirds. Pol. J. Microbiol. 2020, 69, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Sobouti, B.; Mirshekar, M.; Fallah, S.; Tabaei, A.; Mehrabadi, J.F.; Darbandi, A. Pan drug-resistant Acinetobacter baumannii causing nosocomial infections among burnt children. Med. J. Islam. Repub. Iran. 2020, 34, 24. [Google Scholar] [PubMed]
- Goic-Barisic, I.; Music, M.S.; Kovacic, A.; Tonkic, M.; Hrenovic, J. Pan drug-resistant environmental isolate of Acinetobacter baumannii from Croatia. Microb. Drug Resist. 2017, 23, 494–496. [Google Scholar] [CrossRef]
- Quinones, D.; Carvajal, I.; Perez, Y.; Hart, M.; Perez, J.; Garcia, S.; Salazar, D.; Ghosh, S.; Kawaguchiya, M.; Aung, M.S.; et al. High prevalence of bla OXA-23 in Acinetobacter spp. and detection of bla NDM-1 in A. soli in Cuba: Report from national surveillance program (2010–2012). New Microbes New Infect. 2015, 7, 52–56. [Google Scholar] [CrossRef]
- Boluki, E.; Kazemian, H.; Peeridogaheh, H.; Alikhani, M.Y.; Shahabi, S.; Beytollahi, L.; Ghorbanzadeh, R. Antimicrobial activity of photodynamic therapy in combination with colistin against a pan-drug resistant Acinetobacter baumannii isolated from burn patient. Photodiagn. Photodyn. Ther. 2017, 18, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Al-Kadmy, I.M.S.; Ibrahim, S.A.; Al-Saryi, N.; Aziz, S.N.; Besinis, A.; Hetta, H.F. Prevalence of genes involved in colistin resistance in Acinetobacter baumannii: First report from Iraq. Microb. Drug Resist. 2020, 26, 616–622. [Google Scholar] [CrossRef]
- Nowak, J.; Zander, E.; Stefanik, D.; Higgins, P.G.; Roca, I.; Vila, J.; McConnell, M.J.; Cisneros, J.M.; Seifert, H. High incidence of pandrug-resistant Acinetobacter baumannii isolates collected from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J. Antimicrob. Chemother. 2017, 72, 3277–3282. [Google Scholar] [CrossRef]
- Falagas, M.E.; Bliziotis, I.A.; Siempos, I.I. Attributable mortality of Acinetobacter baumannii infections in critically ill patients: A systematic review of matched cohort and case-control studies. Crit. Care 2006, 10, R48. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Clark, N.M.; Lynch, J.P. Infections Due to Acinetobacter baumannii in the ICU: Treatment options. Semin. Respir. Crit. Care Med. 2017, 38, 311–325. [Google Scholar] [CrossRef]
- Scott, P.; Deye, G.; Srinivasan, A.; Murray, C.; Moran, K.; Hulten, E.; Fishbain, J.; Craft, D.; Riddell, S.; Lindler, L.; et al. An outbreak of multidrug-resistant Acinetobacter baumannii-calcoaceticus complex infection in the US military health care system associated with military operations in Iraq. Clin. Infect. Dis. 2007, 44, 1577–1584. [Google Scholar] [CrossRef]
- Öncül, O.; Keskin, Ö.; Acar, H.; Küçükardalı, Y.; Evrenkaya, R.; Atasoyu, E.; Top, C.; Nalbant, S.; Özkan, S.; Emekdaş, G.; et al. Hospital-acquired infections following the 1999 Marmara earthquake. J. Hosp. Infect. 2002, 51, 47–51. [Google Scholar] [CrossRef]
- Rafei, R.; Dabboussi, F.; Hamze, M.; Eveillard, M.; Lemarie, C.; Mallat, H.; Rolain, J.-M.; Joly-Guillou, M.-L.; Kempf, M. First report of blaNDM-1-producing Acinetobacter baumannii isolated in Lebanon from civilians wounded during the Syrian war. Int. J. Infect. Dis. 2014, 21, 21–23. [Google Scholar] [CrossRef]
- Tao, C.; Kang, M.; Chen, Z.; Xie, Y.; Fan, H.; Qin, L.; Ma, Y. Microbiologic study of the pathogens isolated from wound culture among Wenchuan earthquake survivors. Diagn. Microbiol. Infect. Dis. 2009, 63, 268–270. [Google Scholar] [CrossRef]
- Lai, C.-C.; Wang, C.-Y.; Hsueh, P.-R. Co-infections among patients with COVID-19: The need for combination therapy with non-anti-SARS-CoV-2 agents? J. Microbiol. Immunol. Infect. 2020, 53, 505–512. [Google Scholar] [CrossRef]
- Sharifipour, E.; Shams, S.; Esmkhani, M.; Khodadadi, J.; Fotouhi-Ardakani, R.; Koohpaei, A.; Doosti, Z.; Golzari, S.E. Evaluation of bacterial co-infections of the respiratory tract in COVID-19 patients admitted to ICU. BMC Infect. Dis. 2020, 20, 1–7. [Google Scholar] [CrossRef]
- Perez, S.; Innes, G.K.; Walters, M.S.; Mehr, J.; Arias, J.; Greeley, R.; Chew, D. Increase in hospital-acquired carbapenem-resistant Acinetobacter baumannii infection and colonization in an acute care hospital during a surge in COVID-19 admissions—New Jersey, February–July 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 1827. [Google Scholar] [CrossRef]
- Chen, X.; Liao, B.; Cheng, L.; Peng, X.; Xu, X.; Li, Y.; Hu, T.; Li, J.; Zhou, X.; Ren, B. The microbial coinfection in COVID-19. Appl. Microbiol. Biotechnol. 2020, 104, 1–9. [Google Scholar] [CrossRef]
- Anstey, N.M.; Currie, B.J.; Hassell, M.; Palmer, D.; Dwyer, B.; Seifert, H. Community-Acquired Bacteremic Acinetobacter Pneumonia in tropical Australia is caused by diverse strains of Acinetobacter baumannii, with carriage in the throat in at-risk groups. J. Clin. Microbiol. 2002, 40, 685–686. [Google Scholar] [CrossRef]
- Chen, M.-Z.; Hsueh, P.-R.; Lee, L.-N.; Yu, C.-J.; Yang, P.-C.; Luh, K.-T. Severe community-acquired pneumonia due to Acinetobacter baumannii. Chest 2001, 120, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Leung, W.-S.; Chu, C.-M.; Tsang, K.-Y.; Lo, F.-H.; Lo, K.-F.; Ho, P.-L. Fulminant community-acquired Acinetobacter baumannii pneumonia as a distinct clinical syndrome. Chest 2006, 129, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.W.; Lye, D.C.; Khoo, K.L.; Chua, G.S.W.; Yeoh, S.F.; Leo, Y.S.; Tambyah, P.A.; Chua, A.C. Severe community-acquired Acinetobacter baumannii pneumonia: An emerging highly lethal infectious disease in the Asia–Pacific. Respirology 2009, 14, 1200–1205. [Google Scholar] [CrossRef]
- Dexter, C.; Murray, G.L.; Paulsen, I.T.; Peleg, A.Y. Community-acquired Acinetobacter baumannii: Clinical characteris-tics, epidemiology and pathogenesis. Expert Rev. Anti-Infect. Ther. 2015, 13, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, D.N.; Elbourne, L.D.H.; Hassan, K.A.; Eijkelkamp, B.A.; Tetu, S.G.; Brown, M.H.; Shah, B.S.; Peleg, A.; Mabbutt, B.C.; Paulsen, I.T. The complete genome and phenome of a community-acquired Acinetobacter baumannii. PLoS ONE 2013, 8, e58628. [Google Scholar] [CrossRef]
- Liu, X.; Wu, X.; Tang, J.; Zhang, L.; Jia, X. Trends and development in the antibiotic-resistance of Acinetobacter baumannii: A scientometric research study (1991–2019). Infect. Drug Resist. 2020, 13, 3195–3208. [Google Scholar] [CrossRef] [PubMed]
- Central Asian and European Surveillance of Antimicrobial Resistance Annual Report 2019; WHO Regional Office for Europe: Geneva, Switzerland, 2019.
- Surveillance of Antimicrobial Resistance in Europe 2018: Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net); European Centre for Disease Prevention and Control: Solna, Sweden, 2019.
- Lob, S.H.; Hoban, D.J.; Sahm, D.F.; Badal, R.E. Regional differences and trends in antimicrobial susceptibility of Acinetobacter baumannii. Int. J. Antimicrob. Agents 2016, 47, 317–323. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Acinetobacter baumannii infections among patients at military medical facilities treating injured US service members, 2002–2004. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 1063.
- Moradi, J.; Hashemi, F.B.; Bahador, A. Antibiotic resistance of Acinetobacter baumannii in Iran: A systemic review of the published literature. Osong Public Health Res. Perspect. 2015, 6, 79–86. [Google Scholar] [CrossRef]
- Bolourchi, N.; Azizi, O.; Tabrizi, A.M.A.; Esmaeili, S.; Fereshteh, S.; Badmasti, F. Clonal relatedness of Acinetobacter baumannii isolated from the middle east: A systematic review. Rev. Med. Microbiol. 2020. [Google Scholar] [CrossRef]
- Griffith, M.E.; Ceremuga, J.M.; Ellis, M.W.; Guymon, C.H.; Hospenthal, D.R.; Murray, C.K. Acinetobacter skin colonization of US army soldiers. Infect. Control Hosp. Epidemiol. 2006, 27, 659–661. [Google Scholar] [CrossRef]
- Griffith, M.E.; Lazarus, D.R.; Mann, P.B.; Boger, J.A.; Hospenthal, D.R.; Murray, C.K. Acinetobacter skin carriage among US army soldiers deployed in Iraq. Infect. Control Hosp. Epidemiol. 2007, 28, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.F., 3rd; Robinson, B.J.; Hospenthal, D.R.; Aldous, W.K.; Wolf, S.E.; Chung, K.K.; Murray, C.K. Prevalence of multidrug-resistant organisms recovered at a military burn center. Burns 2010, 36, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Echols, R.; Magee, G.; Ferreira, J.C.A.; Morgan, G.; Ariyasu, M.; Sawada, T.; Nagata, T.D. Prevalence of carbapenem-resistant gram-negative infections in the United States predominated by Acinetobacter baumannii and Pseudomonas aeruginosa. Open Forum Infect. Dis. 2017, 4, ofx176. [Google Scholar] [CrossRef] [PubMed]
- Manikal, V.M.; Landman, D.; Saurina, G.; Oydna, E.; Lal, H.; Quale, J. Endemic carbapenem-resistant Acinetobacter species in Brooklyn, New York: Citywide prevalence, interinstitutional spread, and relation to antibiotic usage. Clin. Infect. Dis. 2000, 31, 101–106. [Google Scholar] [CrossRef]
- Yan, Z.; Yang, J.; Hu, R.; Hu, X.; Chen, K. Acinetobacter baumannii infection and IL-17 mediated immunity. Mediat. Inflamm. 2016, 2016, 1–5. [Google Scholar] [CrossRef]
- Choi, C.H.; Lee, J.S.; Lee, Y.C.; Park, T.I.; Lee, J.C. Acinetobacter baumannii invades epithelial cells and outer mem-brane protein A mediates interactions with epithelial cells. BMC Microbiol. 2008, 8, 216. [Google Scholar] [CrossRef]
- Choi, C.H.; Hyun, S.H.; Lee, J.Y.; Lee, J.S.; Lee, Y.S.; Kim, S.A.; Chae, J.P.; Yoo, S.M.; Lee, J.C. Acinetobacter baumannii outer membrane protein A targets the nucleus and induces cytotoxicity. Cell. Microbiol. 2008, 10, 309–319. [Google Scholar] [CrossRef]
- Choi, C.H.; Lee, E.Y.; Lee, Y.C.; Park, T.I.; Kim, H.J.; Hyun, S.H.; Kim, S.A.; Lee, S.-K.; Lee, J.C. Outer membrane protein 38 of Acinetobacter baumannii localizes to the mitochondria and induces apoptosis of epithelial cells. Cell. Microbiol. 2005, 7, 1127–1138. [Google Scholar] [CrossRef]
- Choi, J.; Park, Y.; Kim, C.; Park, Y.; Yoon, H.; Shin, S.; Kim, Y.A.; Song, Y.; Yong, D.; Lee, K. Mortality risk factors of Acinetobacter baumannii bacteraemia. Intern. Med. J. 2005, 35, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Smani, Y.; Dominguez-Herrera, J.; Pachón, J. Association of the outer membrane protein Omp33 with fitness and virulence of Acinetobacter baumannii. J. Infect. Dis. 2013, 208, 1561–1570. [Google Scholar] [CrossRef] [PubMed]
- Rumbo, C.; Tomás, M.; Moreira, E.F.; Soares, N.C.; Carvajal, M.; Santillana, E.; Beceiro, A.; Romero, A.; Bou, G. The Acinetobacter baumannii Omp33–36 porin is a virulence factor that induces apoptosis and modulates autophagy in human cells. Infect. Immun. 2014, 82, 4666–4680. [Google Scholar] [CrossRef]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect. Immun. 2011, 80, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Merino, M.; Rumbo-Feal, S.; Alvarez-Fraga, L.; Vallejo, J.A.; Beceiro, A.; Ohneck, E.J.; Mateos, J.; Fernandez-Puente, P.; Actis, L.A.; et al. The FhaB/FhaC two-partner secretion system is involved in adhesion of Acinetobacter baumannii AbH12O-A2 strain. Virulence 2017, 8, 959–974. [Google Scholar] [CrossRef] [PubMed]
- Weidensdorfer, M.; Chae, J.I.; Makobe, C.; Stahl, J.; Averhoff, B.; Muller, V.; Schurmann, C.; Brandes, R.P.; Wilharm, G.; Ballhorn, W.; et al. Analysis of endothelial adherence of Bartonella henselae and Acinetobacter baumannii using a dynamic human ex vivo infection model. Infect. Immun. 2015, 84, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Bentancor, L.V.; Camacho-Peiro, A.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litran, T. Identification of Ata, a multifunctional trimeric autotransporter of Acinetobacter baumannii. J. Bacteriol. 2012, 194, 3950–3960. [Google Scholar] [CrossRef]
- Weidensdorfer, M.; Ishikawa, M.; Hori, K.; Linke, D.; Djahanschiri, B.; Iruegas, R.; Ebersberger, I.; Riedel-Christ, S.; Enders, G.; Leukert, L.; et al. The Acinetobacter trimeric autotransporter adhesin Ata controls key virulence traits of Acinetobacter baumannii. Virulence 2019, 10, 68–81. [Google Scholar] [CrossRef]
- Liou, M.-L.; Soo, P.-C.; Ling, S.-R.; Kuo, H.-Y.; Tang, C.Y.; Chang, K.-C. The sensor kinase BfmS mediates virulence in Acinetobacter baumannii. J. Microbiol. Immunol. Infect. 2014, 47, 275–281. [Google Scholar] [CrossRef]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef]
- Wood, C.R.; Ohneck, E.J.; Edelmann, R.E.; Actis, L.A. A light-regulated type I pilus contributes to Acinetobacter baumannii biofilm, motility, and virulence functions. Infect. Immun. 2018, 86, e00442-18. [Google Scholar] [CrossRef]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Papadimitrious, M.S.; Paulsen, I.T.; Brown, M.H. Adherence and motility characteristics of clinical Acinetobacter baumannii isolates. FEMS Microbiol. Lett. 2011, 323, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, B.A.; Stroeher, U.H.; Hassan, K.A.; Elbourne, L.D.; Paulsen, I.T.; Brown, M.H. H-NS plays a role in expression of Acinetobacter baumannii virulence features. Infect. Immun. 2013, 81, 2574–2583. [Google Scholar] [CrossRef]
- de Breij, A.; Gaddy, J.; van der Meer, J.; Koning, R.; Koster, A.; van den Broek, P.; Actis, L.; Nibbering, P.; Dijkshoorn, L. CsuA/BABCDE-dependent pili are not involved in the adherence of Acinetobacter baumannii ATCC19606(T) to human airway epithelial cells and their inflammatory response. Res. Microbiol. 2009, 160, 213–218. [Google Scholar] [CrossRef] [PubMed]
- May, H.C.; Yu, J.-J.; Zhang, H.; Wang, Y.; Cap, A.P.; Chambers, J.P.; Guentzel, M.N.; Arulanandam, B.P. Thioredoxin-A is a virulence factor and mediator of the type IV pilus system in Acinetobacter baumannii. PLoS ONE 2019, 14, e0218505. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; Hood, I.; Boyd, K.L.; Olson, P.D.; Morrison, J.M.; Carson, S.; Sayood, K.; Iwen, P.C.; Skaar, E.P.; Dunman, P.M. Inactivation of phospholipase D diminishes Acinetobacter baumannii pathogenesis. Infect. Immun. 2010, 78, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- Fiester, S.E.; Arivett, B.A.; Schmidt, R.E.; Beckett, A.C.; Ticak, T.; Carrier, M.V.; Ghosh, R.; Ohneck, E.J.; Metz, M.L.; Jeffries, M.; et al. Iron-regulated phospholipase C activity contributes to the cytolytic activity and virulence of Acinetobacter baumannii. PLoS ONE 2016, 11, e0167068. [Google Scholar] [CrossRef]
- Stahl, J.; Bergmann, H.; Göttig, S.; Ebersberger, I.; Averhoff, B. Acinetobacter baumannii virulence is mediated by the concerted action of three phospholipases D. PLoS ONE 2015, 10, e0138360. [Google Scholar] [CrossRef]
- Smani, Y.; Docobo-Pérez, F.; López-Rojas, R.; Domínguez-Herrera, J.; Ibáñez-Martínez, J.; Pachón, J. Platelet-activating factor receptor initiates contact of Acinetobacter baumannii expressing phosphorylcholine with host cells. J. Biol. Chem. 2012, 287, 26901–26910. [Google Scholar] [CrossRef]
- Johansson, M.M.; Azzouz, M.; Häggendal, B.; Säljö, K.; Malmi, H.; Zavialov, A.; Teneberg, S. Glycosphingolipids recognized by Acinetobacter baumannii. Microorganisms 2020, 8, 612. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef]
- Erridge, C.; Moncayo-Nieto, O.L.; Morgan, R.; Young, M.; Poxton, I.R. Acinetobacter baumannii lipopolysaccharides are potent stimulators of human monocyte activation via Toll-like receptor 4 signalling. J. Med. Microbiol. 2007, 56, 165–171. [Google Scholar] [CrossRef]
- Luke, N.R.; Sauberan, S.L.; Russo, T.A.; Beanan, J.M.; Olson, R.; Loehfelm, T.W.; Cox, A.D.; Michael, F.S.; Vinogradov, E.V.; Campagnari, A.A. Identification and characterization of a glycosyltransferase involved in Acinetobacter baumannii lipopolysaccharide core biosynthesis. Infect. Immun. 2010, 78, 2017–2023. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Baquir, B.; Tomaras, A.; Montgomery, J.I.; Barbacci, E.G.; Hujer, K.; Bonomo, R.A. Inhibition of LpxC protects mice from resistant Acinetobacter baumannii by modulating inflammation and enhancing phagocytosis. MBio 2012, 3, e00312-12. [Google Scholar] [CrossRef] [PubMed]
- McQueary, C.N.; Kirkup, B.C.; Si, Y.; Barlow, M.; Actis, L.A.; Craft, D.W.; Zurawski, D.V. Extracellular stress and lipopolysaccharide modulate Acinetobacter baumannii surface-associated motility. J. Microbiol. 2012, 50, 434–443. [Google Scholar] [CrossRef] [PubMed]
- McConnell, M.J.; Actis, L.A.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-W.; Lee, S.-H.; Lee, D.-H. Relationship of serum gamma-glutamyltransferase levels with pulmonary function and chronic obstructive pulmonary disease. Lung 2014, 192, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Ermis, H.; Celik, M.R.; Gulbas, G.; Tavli, D.; Aytemur, Z.A. Relationship between serum γ-glutamyltransferase levels and acute exacerbation of chronic obstructive pulmonary disease. Pol. Arch. Intern. Med. 2013, 123, 85–90. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elhossieny, N.M.; Elhezawy, N.B.; Sayed, R.M.; Khattab, M.S.; el Far, M.Y.; Attia, A.S. Gamma-glutamyl transferase is a novel virulence factor of Acinetobacter baumannii that induces alveolar wall destruction and renal damage in systemic pathology. J. Infect. Dis. 2020, 222, 871–879. [Google Scholar] [CrossRef]
- Carruthers, M.D.; Nicholson, P.A.; Tracy, E.N.; Munson, R.S., Jr. Acinetobacter baumannii utilizes a type VI secretion system for bacterial competition. PLoS ONE 2013, 8, e59388. [Google Scholar] [CrossRef]
- Bannister, B.A.; Begg, N.T.; Gillespie, S.H. Infectious Disease, 2nd ed.; Blackwell Science: Hoboken, NJ, USA, 2000. [Google Scholar]
- Wyres, K.L.; Cahill, S.M.; Holt, K.E.; Hall, R.M.; Kenyon, J.J. Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 2020, 6, e000339. [Google Scholar] [CrossRef]
- Russo, T.A.; Luke, N.R.; Beanan, J.M.; Olson, R.; Sauberan, S.L.; Macdonald, U.; Schultz, L.W.; Umland, T.C.; Campagnari, A.A. The K1 capsular polysaccharide of Acinetobacter baumannii Strain 307-0294 is a major virulence factor. Infect. Immun. 2010, 78, 3993–4000. [Google Scholar] [CrossRef]
- Iwashkiw, J.A.; Seper, A.; Weber, B.S.; Scott, N.E.; Vinogradov, E.; Stratilo, C.; Reiz, B.; Cordwell, S.J.; Whittal, R.; Schild, S. Identification of a general O-linked protein glycosylation system in Acinetobacter baumannii and its role in virulence and biofilm formation. PLoS Pathog. 2012, 8, e1002758. [Google Scholar] [CrossRef]
- Lees-Miller, R.G.; Iwashkiw, J.A.; Scott, N.E.; Seper, A.; Vinogradov, E.; Schild, S.; Feldman, M.F. A common pathway for O-linked protein-glycosylation and synthesis of capsule in Acinetobacter baumannii. Mol. Microbiol. 2013, 89, 816–830. [Google Scholar] [CrossRef]
- Zimbler, D.L.; Park, T.M.; Arivett, B.A.; Penwell, W.F.; Greer, S.M.; Woodruff, T.M.; Tierney, D.L.; Actis, L.A. Stress response and virulence functions of the Acinetobacter baumannii NfuA FeS scaffold protein. J. Bacteriol. 2012, 194, 2884–2893. [Google Scholar] [CrossRef]
- Juttukonda, L.J.; Chazin, W.J.; Skaar, E.P. Acinetobacter baumannii coordinates urea metabolism with metal import to resist host-mediated metal limitation. mBio 2016, 7, e01475-16. [Google Scholar] [CrossRef]
- Mortensen, B.L.; Skaar, E.P. The contribution of nutrient metal acquisition and metabolism to Acinetobacter baumannii survival within the host. Front. Cell. Infect. Microbiol. 2013, 3, 95. [Google Scholar] [CrossRef]
- Nairn, B.L.; Lonergan, Z.; Wang, J.; Braymer, J.; Zhang, Y.; Calcutt, M.W.; Lisher, J.P.; Gilston, B.A.; Chazin, W.J.; De Crécy-Lagard, V.; et al. The response of Acinetobacter baumannii to zinc starvation. Cell Host Microbe 2016, 19, 826–836. [Google Scholar] [CrossRef]
- Koenigs, A.; Stahl, J.; Averhoff, B.; Göttig, S.; Wichelhaus, T.A.; Wallich, R.; Zipfel, P.F.; Kraiczy, P. CipA of Acinetobacter baumannii is a novel plasminogen binding and complement inhibitory protein. J. Infect. Dis. 2016, 213, 1388–1399. [Google Scholar] [CrossRef]
- Elhosseiny, N.M.; Amin, M.A.; Yassin, A.S.; Attia, A.S. Acinetobacter baumannii universal stress protein A plays a pivotal role in stress response and is essential for pneumonia and sepsis pathogenesis. Int. J. Med Microbiol. 2015, 305, 114–123. [Google Scholar] [CrossRef]
- Gebhardt, M.J.; Gallagher, L.A.; Jacobson, R.K.; Usacheva, E.A.; Peterson, L.R.; Zurawski, D.V.; Shuman, H.A. Joint transcritional control of virulence and resistance to antibiotic and environmental stress in Acinetobacter baumannii. MBio 2015, 6, e01660-15. [Google Scholar] [CrossRef]
- O’Connor, A.; McClean, S. The role of universal stress proteins in bacterial infections. Curr. Med. Chem. 2017, 24, 3970–3979. [Google Scholar] [CrossRef]
- Sun, D.; Crowell, S.A.; Harding, C.M.; De Silva, P.M.; Harrison, A.; Fernando, D.M.; Mason, K.M.; Santana, E.; Loewen, P.C.; Kumar, A.; et al. KatG and KatE confer Acinetobacter resistance to hydrogen peroxide but sensitize bacteria to killing by phagocytic respiratory burst. Life Sci. 2016, 148, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Murray, G.L.; Peleg, A.; Doi, Y. Acinetobacter baumannii: Evolution of Antimicrobial Resistance—Treatment Options. Semin. Respir. Crit. Care Med. 2015, 36, 085–098. [Google Scholar] [CrossRef]
- Gaddy, J.A.; Arivett, B.A.; McConnell, M.J.; Lopez-Rojas, R.; Pachon, J.; Actis, L.A. Role of acinetobactin-mediated iron acquisition functions in the interaction of Acinetobacter baumannii strain ATCC 19606T with human lung epithelial cells, Galleria mellonella caterpillars, and mice. Infect. Immun. 2012, 80, 1015–1024. [Google Scholar] [CrossRef]
- De Gregorio, E.; Del Franco, M.; Martinucci, M.; Roscetto, E.; Zarrilli, R.; Di Nocera, P.P. Biofilm-associated proteins: News from Acinetobacter. BMC Genom. 2015, 16, 933. [Google Scholar] [CrossRef]
- Roussin, M.; Rabarioelina, S.; Cluzeau, L.; Cayron, J.; Lesterlin, C.; Salcedo, S.P.; Bigot, S. Identification of a contact-dependent growth inhibition (CDI) system that reduces biofilm formation and host cell adhesion of Acinetobacter baumannii DSM30011 strain. Front. Microbiol. 2019, 10, 2450. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, G.M.; Kostoulias, X.; Khoo, C.; Aibinu, I.; Qu, Y.; Traven, A.; Peleg, A. A global virulence regulator in Acinetobacter baumannii and its control of the phenylacetic acid catabolic pathway. J. Infect. Dis. 2014, 210, 46–55. [Google Scholar] [CrossRef]
- Repizo, G.D.; Gagné, S.; Foucault-Grunenwald, M.-L.; Borges, V.; Charpentier, X.; Limansky, A.S.; Gomes, J.P.; Viale, A.M.; Salcedo, S.P. Differential role of the T6SS in Acinetobacter baumannii virulence. PLoS ONE 2015, 10, e0138265. [Google Scholar] [CrossRef]
- Seder, R.A.; Hill, A.V.S. Vaccines against intracellular infections requiring cellular immunity. Nat. Cell Biol. 2000, 406, 793–798. [Google Scholar] [CrossRef]
- McConnell, M.J.; Domínguez-Herrera, J.; Smani, Y.; López-Rojas, R.; Docobo-Pérez, F.; Pachón, J. Vaccination with outer membrane complexes elicits rapid protective immunity to multidrug-resistant Acinetobacter baumannii. Infect. Immun. 2010, 79, 518–526. [Google Scholar] [CrossRef]
- McConnell, M.J.; Pachón, J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine 2010, 29, 1–5. [Google Scholar] [CrossRef]
- McConnell, M.J.; Rumbo, C.; Bou, G.; Pachón, J. Outer membrane vesicles as an acellular vaccine against Acinetobacter baumannii. Vaccine 2011, 29, 5705–5710. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, S.; Ketter, P.M.; Yu, J.-J.; Grimm, R.C.; May, H.C.; Cap, A.P.; Chambers, J.P.; Guentzel, M.N.; Arulanan-dam, B.P. Vaccination with a live attenuated Acinetobacter baumannii deficient in thioredoxin provides protection against systemic Acinetobacter infection. Vaccine 2017, 35, 3387–3394. [Google Scholar] [CrossRef]
- Shu, M.H.; MatRahim, N.; NorAmdan, N.; Pang, S.P.; Hashim, S.H.; Phoon, W.H.; AbuBakar, S. An Inactivated antibiotic exposed whole-cell vaccine enhances bactericidal activities against multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 22332. [Google Scholar] [CrossRef]
- KuoLee, R.; Harris, G.; Yan, H.; Xu, H.H.; Conlan, W.J.; Patel, G.B.; Chen, W. Intranasal immunization protects against Acinetobacter baumannii-associated pneumonia in mice. Vaccine 2015, 33, 260–267. [Google Scholar] [CrossRef] [PubMed]
- García-Quintanilla, M.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Immunization with lipopolysaccharide-deficient whole cells provides protective immunity in an experimental mouse model of Acinetobacter baumannii Infection. PLoS ONE 2014, 9, e114410. [Google Scholar] [CrossRef]
- Wizemann, T.M.; Adamou, J.E.; Langermann, S. Adhesins as targets for vaccine development. Emerg. Infect. Dis. 1999, 5, 395–403. [Google Scholar] [CrossRef]
- Jahangiri, A.; Rasooli, I.; Owlia, P.; Fooladi, A.A.I.; Salimian, J. In silico design of an immunogen against Acinetobacter baumannii based on a novel model for native structure of outer membrane protein A. Microb. Pathog. 2017, 105, 201–210. [Google Scholar] [CrossRef]
- Nie, D.; Hu, Y.; Chen, Z.; Li, M.; Hou, Z.; Luo, X.; Mao, X.; Xue, X. Outer membrane protein A (OmpA) as a potential therapeutic target for Acinetobacter baumannii infection. J. Biomed. Sci. 2020, 27, 1–8. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, T.; Cao, J.; Sun, J.; Dai, W.; Zhang, L. Mucosal immunization with purified OmpA elicited protective immunity against infections caused by multidrug-resistant Acinetobacter baumannii. Microb. Pathog. 2016, 96, 20–25. [Google Scholar] [CrossRef]
- Luo, G.; Lin, L.; Ibrahim, A.S.; Baquir, B.; Pantapalangkoor, P.; Bonomo, R.A.; Doi, Y.; Adams, M.D.; Russo, T.A.; Spellberg, B. Active and Passive Immunization protects against lethal, extreme drug resistant-Acinetobacter baumannii infection. PLoS ONE 2012, 7, e29446. [Google Scholar] [CrossRef]
- Lin, L.; Tan, B.; Pantapalangkoor, P.; Ho, T.; Hujer, A.M.; Taracila, M.A.; Bonomo, R.A.; Spellberg, B. Acinetobacter baumannii rOmpA vaccine dose alters immune polarization and immunodominant epitopes. Vaccine 2013, 31, 313–318. [Google Scholar] [CrossRef]
- Islam, A.H.M.S.; Singh, K.-K.B.; Ismail, A. Demonstration of an outer membrane protein that is antigenically specific for Acinetobacter baumannii. Diagn. Microbiol. Infect. Dis. 2011, 69, 38–44. [Google Scholar] [CrossRef]
- Jahangiri, A.; Rasooli, I.; Owlia, P.; Fooladi, A.A.I.; Salimian, J. Highly conserved exposed immunogenic peptides of Omp34 against Acinetobacter baumannii: An innovative approach. J. Microbiol. Methods 2018, 144, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Bentancor, L.V.; Routray, A.; Bozkurt-Guzel, C.; Camacho-Peiro, A.; Pier, G.B.; Maira-Litran, T. Evaluation of the trimeric autotransporter Ata as a vaccine candidate against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 3381–3388. [Google Scholar] [CrossRef]
- Bentancor, L.V.; O’Malley, J.M.; Bozkurt-Guzel, C.; Pier, G.B.; Maira-Litran, T. Poly-N-acetyl-beta-(1–6)-glucosamine is a target for protective immunity against Acinetobacter baumannii infections. Infect. Immun. 2012, 80, 651–656. [Google Scholar] [CrossRef]
- Russo, T.A.; Beanan, J.M.; Olson, R.; Macdonald, U.; Cox, A.D.; Michael, F.S.; Vinogradov, E.V.; Spellberg, B.; Luke-Marshall, N.R.; Campagnari, A.A. The K1 capsular polysaccharide from Acinetobacter baumannii is a potential therapeutic target via passive immunization. Infect. Immun. 2013, 81, 915–922. [Google Scholar] [CrossRef]
- Fattahian, Y.; Rasooli, I.; Gargari, S.L.M.; Rahbar, M.R.; Astaneh, S.D.; Amani, J. Protection against Acinetobacter baumannii infection via its functional deprivation of biofilm associated protein (Bap). Microb. Pathog. 2011, 51, 402–406. [Google Scholar] [CrossRef]
- Aghajani, Z.; Rasooli, I.; Gargari, S.L.M. Exploitation of two siderophore receptors, BauA and BfnH, for protection against Acinetobacter baumannii infection. Apmis 2019, 127, 753–763. [Google Scholar] [CrossRef]
- Huang, W.; Yao, Y.; Wang, S.; Xia, Y.; Yang, X.; Long, Q.; Sun, W.; Liu, C.; Li, Y.; Chu, X.; et al. Immunization with a 22-kDa outer membrane protein elicits protective immunity to multidrug-resistant Acinetobacter baumannii. Sci. Rep. 2016, 6, 20724. [Google Scholar] [CrossRef]
- Casey, W.T.; Spink, N.; Cia, F.; Collins, C.; Romano, M.; Berisio, R.; Bancroft, G.J.; McClean, S. Identification of an OmpW homologue in Burkholderia pseudomallei, a protective vaccine antigen against melioidosis. Vaccine 2016, 34, 2616–2621. [Google Scholar] [CrossRef]
- Singh, R.; Capalash, N.; Sharma, P. Immunoprotective potential of BamA, the outer membrane protein assembly factor, against MDR Acinetobacter baumannii. Sci. Rep. 2017, 7, 12411. [Google Scholar] [CrossRef] [PubMed]
- Tohidinia, M.; Moshtaghioun, S.M.; Sefid, F.; Falahati, A. Functional exposed amino acids of Caro analysis as a potential vaccine candidate in Acinetobacter baumannii. Int. J. Pept. Res. Ther. 2019, 26, 1185–1197. [Google Scholar] [CrossRef]
- Catel-Ferreira, M.; Marti, S.; Guillon, L.; Jara, L.; Coadou, G.; Molle, V.; Bouffartigues, E.; Bou, G.; Shalk, I.; Jouenne, T. The outer membrane porin OmpW of Acinetobacter baumannii is involved in iron uptake and colistin binding. FEBS Lett. 2016, 590, 224–231. [Google Scholar] [CrossRef]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Long, Q.; Sun, W.; Liu, C.; Li, Y.; Ma, Y. OmpW is a potential target for eliciting protective immunity against Acinetobacter baumannii infections. Vaccine 2015, 33, 4479–4485. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.J.; Ren, S.; Xie, Y.E. Evaluation of the protective efficacy of a fused OmpK/Omp22 protein vaccine candidate against Acinetobacter baumannii infection in mice. Biomed. Environ. Sci. 2018, 31, 155–158. [Google Scholar]
- Yang, A.; Yang, H.; Guo, S.; Xie, Y. MF59 adjuvant enhances the immunogenicity and protective immunity of the OmpK/Omp22 fusion protein from Acineterbacter baumannii through intratracheal inoculation in mice. Scand. J. Immunol. 2019, 90, e12769. [Google Scholar] [CrossRef]
- García-Patiño, M.G.; García-Contreras, R.; Licona-Limón, P. The immune response against Acinetobacter baumannii, an emerging pathogen in nosocomial infections. Front. Immunol. 2017, 8, 441. [Google Scholar] [CrossRef]
- Henderson, J.C.; Zimmerman, S.M.; Crofts, A.; Boll, J.M.; Kuhns, L.G.; Herrera, C.M.; Trent, M.S. The power of asymmetry: Architecture and assembly of the gram-negative outer membrane lipid bilayer. Annu. Rev. Microbiol. 2016, 70, 255–278. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer membrane vesicles: Current status and future direction of these novel vaccine adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Li, K.; Sun, P.; Liu, C.; Sun, W.; Bai, H.; et al. Employing Escherichia coli-derived outer membrane vesicles as an antigen delivery platform elicits protective immunity against Acinetobacter baumannii infection. Sci. Rep. 2016, 6, 37242. [Google Scholar] [CrossRef]
- Lei, L.; Yang, F.; Zou, J.; Jing, H.; Zhang, J.; Xu, W.; Zou, Q.; Zhang, J.; Wang, X. DNA vaccine encoding OmpA and Pal from Acinetobacter baumannii efficiently protects mice against pulmonary infection. Mol. Biol. Rep. 2019, 46, 5397–5408. [Google Scholar] [CrossRef]
- Ansari, H.; Tahmasebi-Birgani, M.; Bijanzadeh, M.; Doosti, A.; Kargar, M. Study of the immunogenicity of outer membrane protein A (OmpA) gene from Acinetobacter baumannii as DNA vaccine candidate in vivo. Iran. J. Basic Med. Sci. 2019, 22, 669–675. [Google Scholar]
- Rasooli, I.; Abdolhamidi, R.; Jahangiri, A.; Astaneh, S.D.A. Outer membrane protein, Oma87 prevents Acinetobacter baumannii infection. Int. J. Pept. Res. Ther. 2020, 26, 2653–2660. [Google Scholar] [CrossRef]
- Hounsome, J.D.; Baillie, S.; Noofeli, M.; Riboldi-Tunnicliffe, A.; Burchmore, R.J.; Isaacs, N.W.; Davies, R.L. Outerembrane protein A of bovine and ovine isolates of Mannheimia haemolytica is surface exposed and contains host species-specific epitopes. Infect. Immun. 2011, 79, 4332–4341. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).