
Chimeric Protein IPath® with Chelating Activity Improves Atlantic Salmon’s Immunity against Infectious Diseases




Abstract
1. Introduction
2. Materials and Methods
2.1. Design and Expression of a Recombinant Chimeric Protein (IPath®)
2.2. IPath® Iron-Chelating Activity
2.3. Immunization and Infection Trials
2.4. RNA Extraction and RT-qPCR Analysis in Atlantic Salmon Exposed to Different Pathogens
3. Results
3.1. The Iron-Chelating Activity of Chimeric Protein
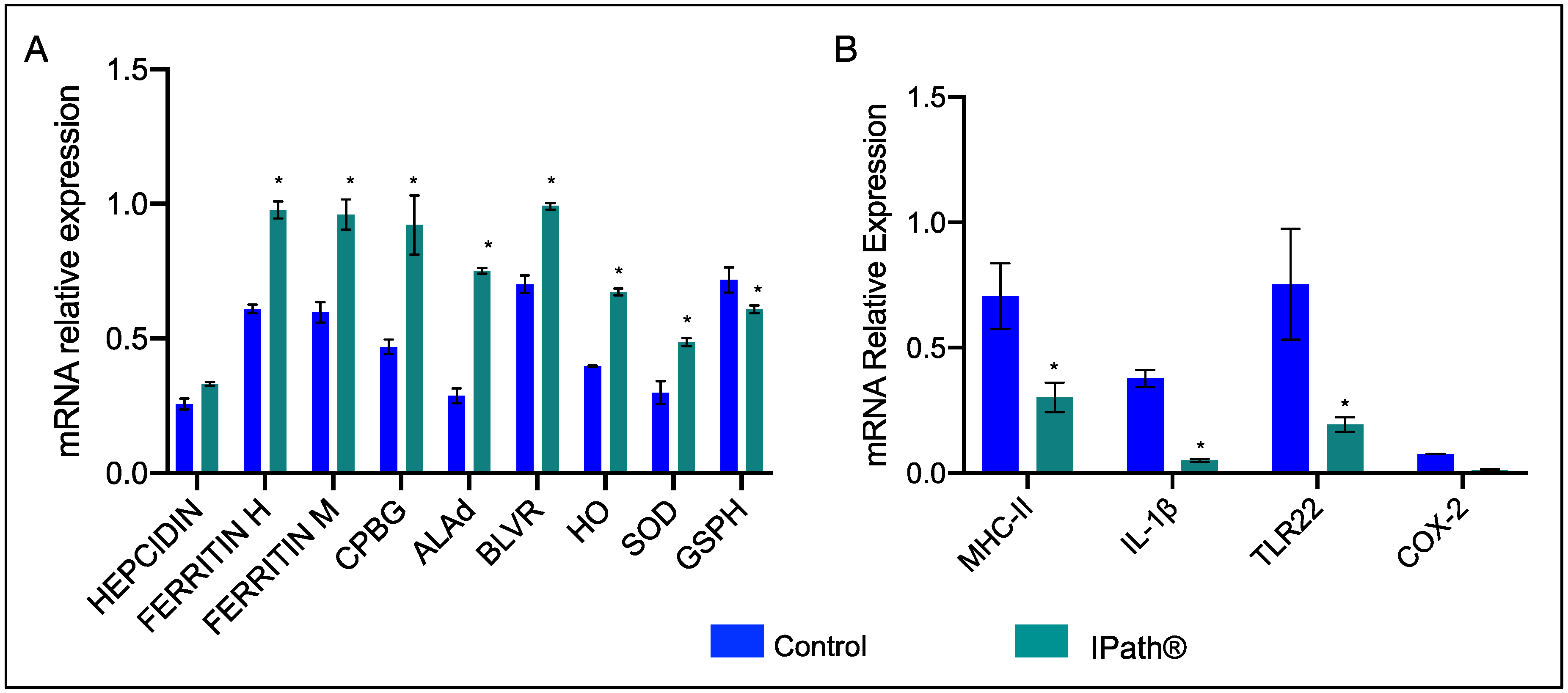
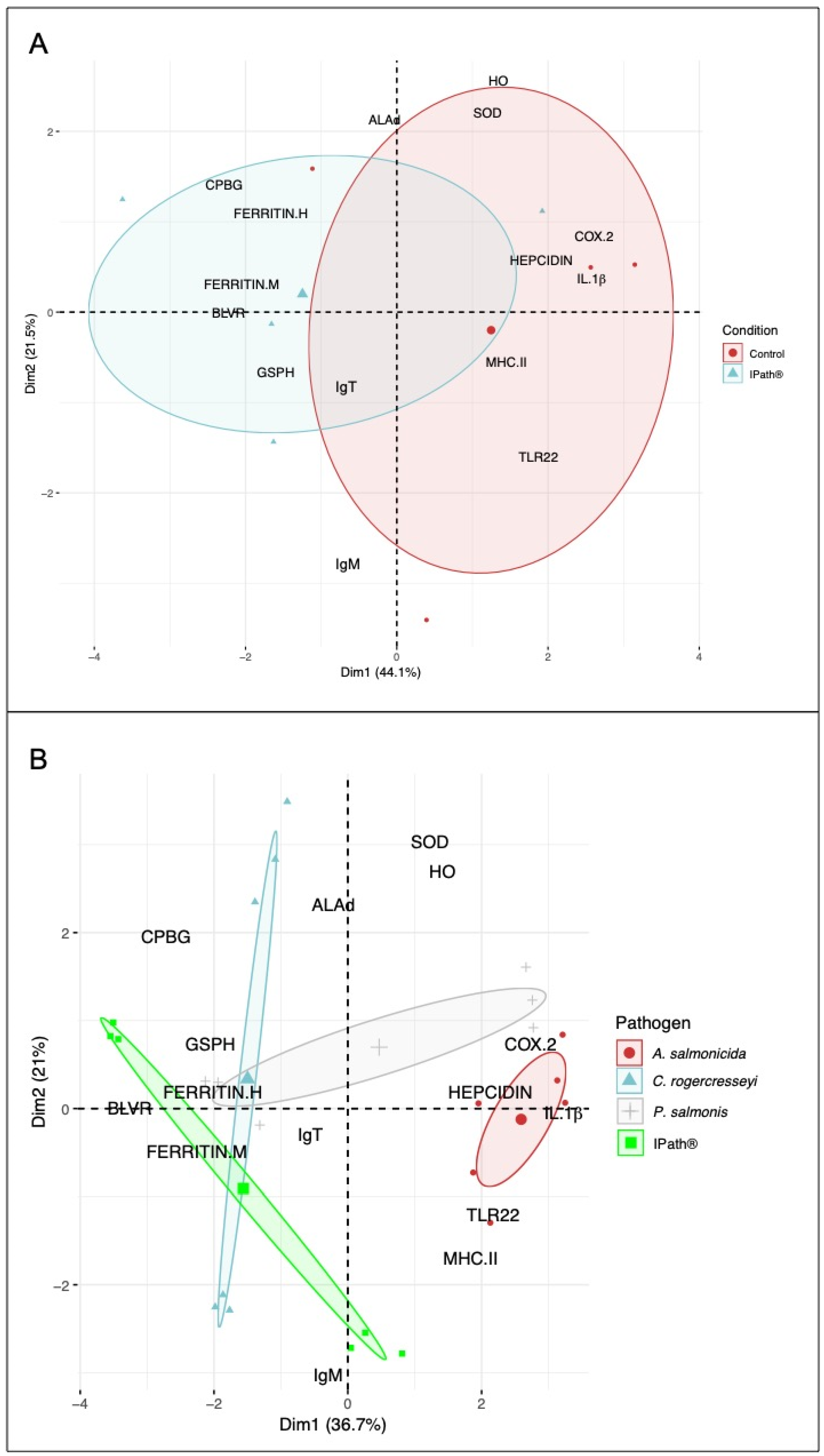
3.2. Modulation of IPath® Vaccine in Atlantic Salmon Genes
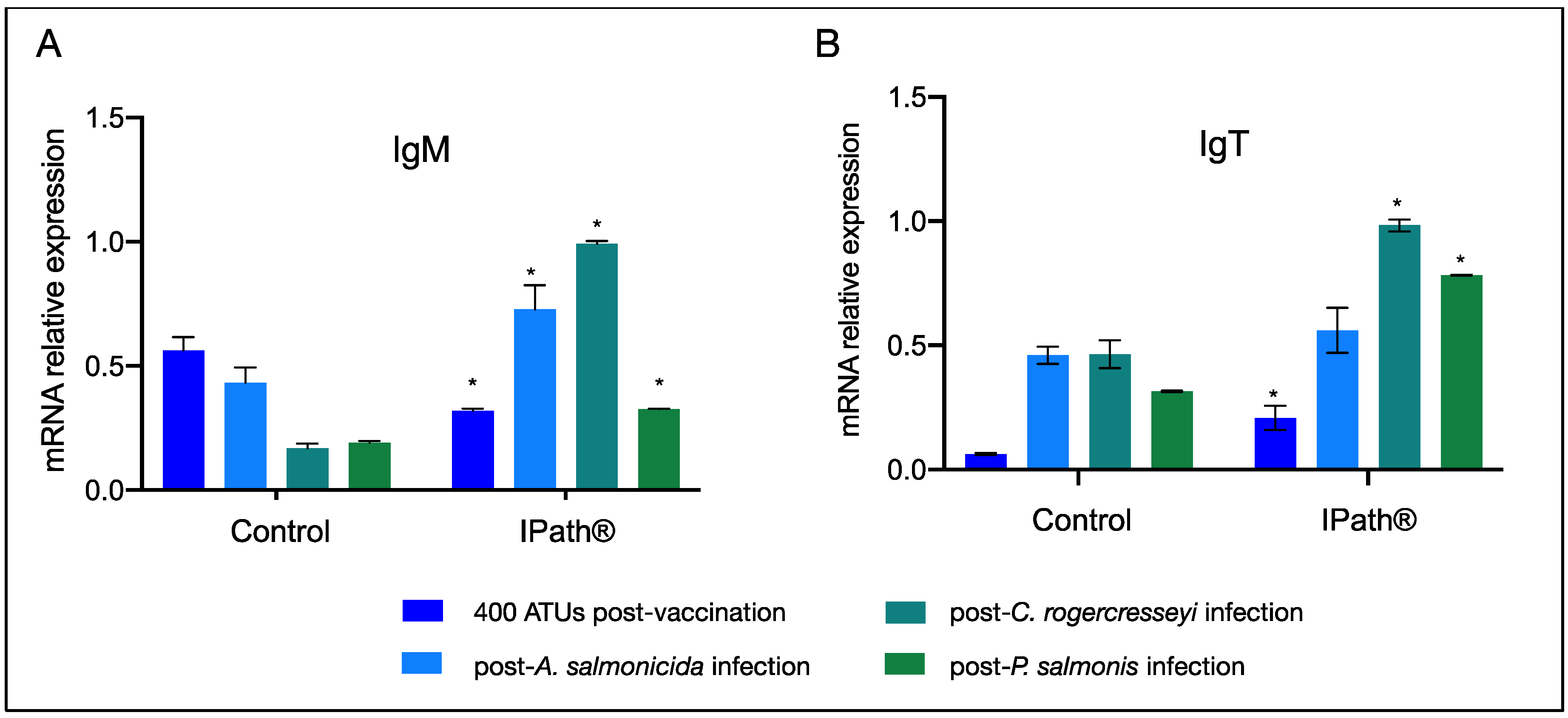
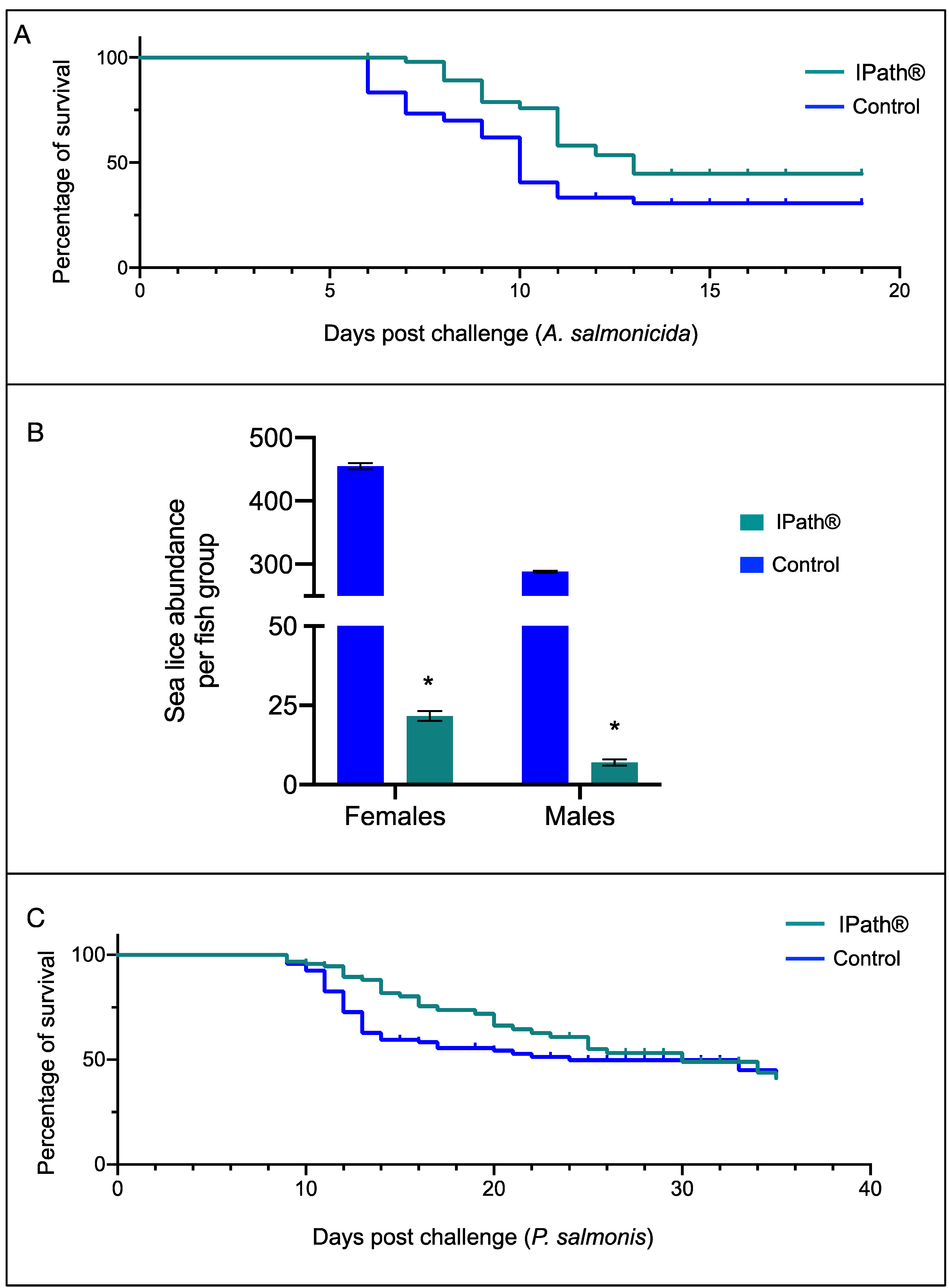
3.3. IPath® Vaccine Effects in Atlantic Salmon’s Response to Pathogen Infection
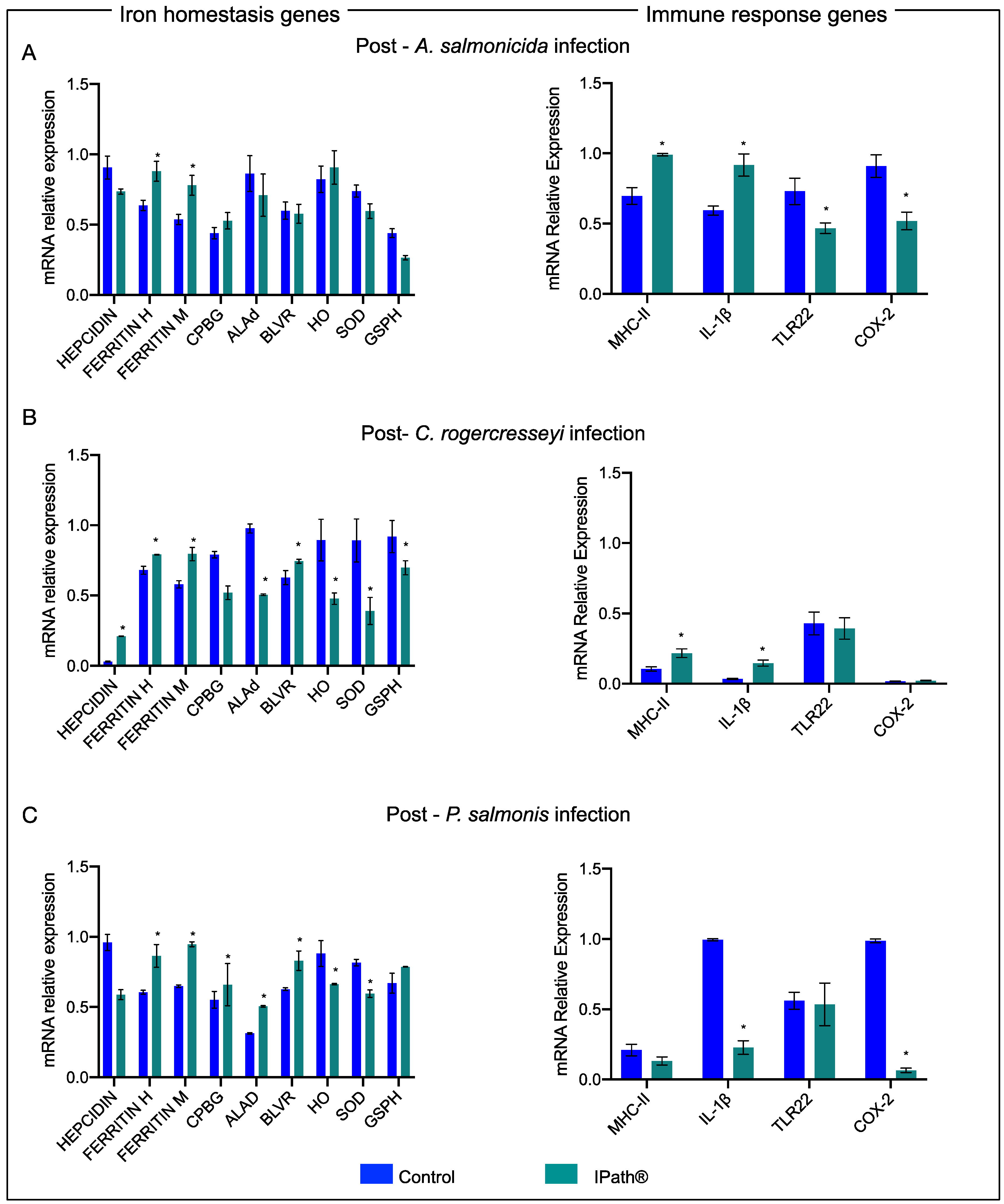
3.4. IPath®-Vaccinated Atlantic Salmon Transcription Expression during the Pathogen Infections
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barragán, G.; Santoyo, A.; Ramos, C. Iron deficiency anaemia. Rev. Med. Hosp. Gen. Mex. 2016, 79, 88–97. [Google Scholar] [CrossRef]
- Chifman, J.; Laubenbacher, R.; Torti, S.V. A systems biology approach to iron metabolism. Adv. Exp. Med. Biol. 2014, 844, 201–225. [Google Scholar]
- Hood, M.; Skaar, E. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Marles-Wright, J. Ferritin family proteins and their use in bionanotechnology. New Biotechnol. 2015, 32, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.V.; Wilson, J.M.; Rodrigues, P.N. Transferrin and ferritin response to bacterial infection: The role of the liver and brain in fish. Dev. Comp. Immunol. 2009, 33, 848–857. [Google Scholar] [CrossRef]
- Rosolen, G.; Eberle, E.; Torres, F.; Musso, A. Conceptos actuales sobre fisiología y patología del hierro. Hematología 2010, 14, 48–57. [Google Scholar]
- Torti, F.; Torti, S. Regulation of ferritin genes and protein. Blood 2002, 99, 3505–3516. [Google Scholar] [CrossRef]
- Kvingedal, A.M.; Rørvik, K.A.; Alestrøm, P. Cloning and characterization of Atlantic salmon (Salmo salar) serum transferrin cDNA. Mol. Mar. Biol. Biotechnol. 1993, 2, 233–238. [Google Scholar]
- Cuesta, A.; Meseguer, J.; Esteban, M.Á. The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol. 2008, 45, 2333–2342. [Google Scholar] [CrossRef]
- Yang, M.; Wang, K.-J.; Chen, J.-H.; Qu, H.-D.; Li, S.-J. Genomic organization and tissue-specific expression analysis of hepcidin-like genes from black porgy (Acanthopagrus schlegelii B.). Fish Shellfish Immunol. 2007, 23, 1060–1071. [Google Scholar] [CrossRef]
- Johnson, E.E.; Wessling-Resnick, M. Iron metabolism and the innate immune response to infection. Microbes Infect. 2012, 14, 207–216. [Google Scholar] [CrossRef]
- Nűũez, G.; Sakamoto, K.; Soares, P. Innate Nutritional Immunity. J. Immunol. 2018, 201, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.N.S.; Vázquez-Dorado, S.; Neves, J.V.; Wilson, J.M. Dual function of fish hepcidin: Response to experimental iron overload and bacterial infection in sea bass (Dicentrarchus labrax). Dev. Comp. Immunol. 2006, 30, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.-j.; Hu, Y.-h.; Sun, L. Identification and analysis of a Scophthalmus maximus ferritin that is regulated at transcription level by oxidative stress and bacterial infection. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 156, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Miranda, D.; Gallardo-Escárate, C. Novel insights into the response of Atlantic salmon (Salmo salar) to Piscirickettsia salmonis: Interplay of coding genes and lncRNAs during bacterial infection. Fish Shellfish Immunol. 2016, 59, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Muñoz, V.; Gallardo-Escárate, C. Iron metabolism modulation in Atlantic salmon infested with the sea lice Lepeophtheirus salmonis and Caligus rogercresseyi: A matter of nutritional immunity? Fish Shellfish Immunol. 2017, 60, 97–102. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Uncovering iron regulation with species-specific transcriptome patterns in Atlantic and coho salmon during a Caligus rogercresseyi infestation. J. Fish Dis. 2017, 40, 1169–1184. [Google Scholar] [CrossRef]
- Ye, T.; Wu, X.; Wu, W.; Dai, C.; Yuan, J. Ferritin protect shrimp Litopenaeus vannamei from WSSV infection by inhibiting virus replication. Fish Shellfish Immunol. 2015, 42, 138–143. [Google Scholar] [CrossRef]
- Yang, H.; Liu, Z.; Jiang, Q.; Xu, J.; An, Z.; Zhang, Y.; Xiong, D.; Wang, L. A novel ferritin gene from Procambarus clarkii involved in the immune defense against Aeromonas hydrophila infection and inhibits WSSV replication. Fish Shellfish Immunol. 2019, 86, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.-j.; Hu, Y.-h.; Xiao, Z.-z.; Sun, L. Cloning and analysis of a ferritin subunit from turbot (Scophthalmus maximus). Fish Shellfish Immunol. 2010, 28, 829–836. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, L.-p.; Li, M.-f.; Sun, L. Turbot (Scophthalmus maximus) hepcidin-1 and hepcidin-2 possess antimicrobial activity and promote resistance against bacterial and viral infection. Fish Shellfish Immunol. 2014, 38, 127–134. [Google Scholar] [CrossRef]
- Yaacob, E.N.; De Geest, B.G.; Goethals, J.; Bajek, A.; Dierckens, K.; Bossier, P.; Vanrompay, D. Recombinant ferritin-H induces immunosuppression in European sea bass larvae (Dicentrarchus labrax) rather than immunostimulation and protection against a Vibrio anguillarum infection. Vet. Immunol. Immunopathol. 2018, 204, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Galay, R.L.; Miyata, T.; Umemiya-Shirafuji, R.; Maeda, H.; Kusakisako, K.; Tsuji, N.; Mochizuki, M.; Fujisaki, K.; Tanaka, T. Evaluation and comparison of the potential of two ferritins as anti-tick vaccines against haemaphysalis longicornis. Parasites Vectors 2014, 7, 482. [Google Scholar] [CrossRef] [PubMed]
- Arriagada, G.; Hamilton-West, C.; Nekouei, O.; Foerster, C.; Müller, A.; Lara, M.; Gallardo-Escárate, C. Caligus rogercresseyi infestation is associated with Piscirickettsia salmonis-attributed mortalities in farmed salmonids in Chile. Prev. Vet. Med. 2019, 171, 104771. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Escarate, C.; Arriagada, G.; Carrera, C.; Goncalves, A.T.; Nunez-Acuna, G.; Valenzuela-Miranda, D.; Valenzuela-Munoz, V. The race between host and sea lice in the Chilean salmon farming: A genomic approach. Rev. Aquacult. 2019, 11, 325–339. [Google Scholar] [CrossRef]
- Gallardo-Escarate, C.; Valenzuela-Munoz, V.; Nunez-Acuna, G.; Carrera, C.; Goncalves, A.T.; Valenzuela-Miranda, D.; Benavente, B.P.; Roberts, S. Catching the complexity of salmon-louse interactions. Fish Shellfish Immunol. 2019, 90, 199–209. [Google Scholar] [CrossRef]
- Rozas, M.; Enríquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: A review. J. Fish Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Miranda, D.; Valenzuela-Munoz, V.; Farlora, R.; Gallardo-Escarate, C. MicroRNA-based transcriptomic responses of Atlantic salmon during infection by the intracellular bacterium Piscirickettsia salmonis. Dev. Comp. Immunol. 2017, 77, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Pontigo, J.P.; Espinoza, C.; Hernandez, M.; Nourdin, G.; Oliver, C.; Avendaño-Herrera, R.; Figueroa, J.; Rauch, C.; Troncoso, J.M.; Vargas-Chacoff, L.; et al. Protein-Based Vaccine Protect Against Piscirickettsia salmonis in Atlantic Salmon (Salmo salar). Front. Immunol. 2021, 12, 602689. [Google Scholar] [CrossRef]
- Valenzuela-Miranda, D.; Gallardo-Escarate, C. Dual RNA-Seq Uncovers Metabolic Amino Acids Dependency of the Intracellular Bacterium Piscirickettsia salmonis Infecting Atlantic salmon. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar] [CrossRef]
- Valdes, N.; Espinoza, C.; Sanhueza, L.; Gonzalez, A.; Corsini, G.; Tello, M. Draft genome sequence of the Chilean isolate Aeromonas salmonicida strain CBA100. FEMS Microbiol. Lett. 2015, 362. [Google Scholar] [CrossRef]
- Zhang, Z.; Niu, C.; Storset, A.; Bogwald, J.; Dalmo, R.A. Comparison of Aeromonas salmonicida resistant and susceptible salmon families: A high immune response is beneficial for the survival against Aeromonas salmonicida challenge. Fish Shellfish Immunol. 2011, 31, 1–9. [Google Scholar] [CrossRef]
- Valderrama, K.; Soto-Davila, M.; Segovia, C.; Vasquez, I.; Dang, M.; Santander, J. Aeromonas salmonicida infects Atlantic salmon (Salmo salar) erythrocytes. J. Fish Dis. 2019, 42, 1601–1608. [Google Scholar] [CrossRef]
- Lee, J.-H.; Pooley, N.J.; Mohd-Adnan, A.; Martin, S.A. Cloning and characterisation of multiple ferritin isoforms in the Atlantic salmon (Salmo salar). PLoS ONE 2014, 9, e103729. [Google Scholar] [CrossRef] [PubMed]
- Pulgar, R.; Hödar, C.; Travisany, D.; Zuñiga, A.; Domínguez, C.; Maass, A.; González, M.; Cambiazo, V. Transcriptional response of Atlantic salmon families to Piscirickettsia salmonis infection highlights the relevance of the iron-deprivation defence system. BMC Genom. 2015, 16, 495. [Google Scholar] [CrossRef] [PubMed]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- De Zoysa, M.; Lee, J. Two ferritin subunits from disk abalone (Haliotis discus discus): Cloning, characterization and expression analysis. Fish Shellfish Immunol. 2007, 23, 624–635. [Google Scholar] [CrossRef]
- Rauta, P.; Nayak, B.; Das, S. Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunol. Lett. 2012, 148, 23–33. [Google Scholar] [CrossRef]
- Tadiso, T.M.; Krasnov, A.; Skugor, S.; Afanasyev, S.; Hordvik, I.; Nilsen, F. Gene expression analyses of immune responses in Atlantic salmon during early stages of infection by salmon louse (Lepeophtheirus salmonis) revealed bi-phasic responses coinciding with the copepod-chalimus transition. BMC Genom. 2011, 12, 141. [Google Scholar] [CrossRef]
- Anderson, G.J.; Vulpe, C.D. Mammalian iron transport. Cell. Mol. Life Sci. 2009, 66, 3241. [Google Scholar] [CrossRef]
- Gkouvatsos, K.; Papanikolaou, G.; Pantopoulos, K. Regulation of iron transport and the role of transferrin. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 188–202. [Google Scholar] [CrossRef]
- Ganz, T. Molecular control of iron transport. J. Am. Soc. Nephrol. 2007, 18, 394–400. [Google Scholar] [CrossRef]
- Nemeth, E.; Ganz, T. Regulation of iron metabolism by hepcidin. Annu. Rev. Nutr. 2006, 26, 323–342. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Hepcidin—a regulator of intestinal iron absorption and iron recycling by macrophages. Best Pract. Res. Clin. Haematol. 2005, 18, 171–182. [Google Scholar] [CrossRef]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of hepcidin gene expression and severe tissue iron overload in upstream stimulatory factor 2 (USF2) knockout mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef]
- Montosi, G.; Corradini, E.; Garuti, C.; Barelli, S.; Recalcati, S.; Cairo, G.; Valli, L.; Pignatti, E.; Vecchi, C.; Ferrara, F. Kupffer cells and macrophages are not required for hepatic hepcidin activation during iron overload. Hepatology 2005, 41, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Standal, H.; RØrvik, K.A.; Lien, H.; Andersen, Ø. Effects of acute iron overload on Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss). Biol. Trace Elem. Res. 1997, 59, 13–22. [Google Scholar] [CrossRef]
- Singh, M.; Barman, A.S.; Devi, A.L.; Devi, A.G.; Pandey, P.K. Iron mediated hematological, oxidative and histological alterations in freshwater fish Labeo rohita. Ecotoxicol. Environ. Saf. 2019, 170, 87–97. [Google Scholar] [CrossRef]
- Rørvik, K.A.; Dehli, A.; Thomassen, M.; Ruyter, B.; Steien, S.H.; Salte, R. Synergistic effects of dietary iron and omega-3 fatty acid levels on survival of farmed Atlantic salmon, Salmo salar L., during natural outbreaks of furunculosis and cold water vibriosis. J. Fish Dis. 2003, 26, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Almarza, O.; Valderrama, K.; Ayala, M.; Segovia, C.; Santander, J. A functional ferric uptake regulator (Fur) protein in the fish pathogen Piscirickettsia salmonis. Int. Microbiol. 2016, 19, 49–55. [Google Scholar] [PubMed]
- Vera-Bizama, F.; Valenzuela-Muñoz, V.; Gonçalves, A.T.; Marambio, J.P.; Hawes, C.; Wadsworth, S.; Gallardo-Escárate, C. Transcription expression of immune-related genes from Caligus rogercresseyi evidences host-dependent patterns on Atlantic and coho salmon. Fish Shellfish Immunol. 2015, 47, 725–731. [Google Scholar] [CrossRef]
- Hajdusek, O.; Sima, R.; Perner, J.; Loosova, G.; Harcubova, A.; Kopacek, P. Tick iron and heme metabolism—New target for an anti-tick intervention. Ticks Tick Borne Dis. 2016, 7, 565–572. [Google Scholar] [CrossRef]
- Perner, J.; Sobotka, R.; Sima, R.; Konvickova, J.; Sojka, D.; Oliveira, P.L.; Hajdusek, O.; Kopacek, P. Acquisition of exogenous haem is essential for tick reproduction. eLife 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Wessling-Resnick, M. Iron homeostasis and the inflammatory response. Annu. Rev. Nutr. 2010, 30, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Tamilselvam, B.; Hansen, E.J.; Daefler, S. Modulation of iron homeostasis in macrophages by bacterial intracellular pathogens. BMC Microbiol. 2010, 10, 64. [Google Scholar] [CrossRef]
- Thorson, J.A.; Smith, K.M.; Gomez, F.; Naumann, P.W.; Kemp, J.D. Role of iron in T cell activation: Th1 clones differ from TH2 clones in their sensitivity to inhibition of DNA synthesis caused by IGG mabs against the transferrin receptor and the iron chelator deferoxamine. Cell. Immunol. 1991, 134, 126–137. [Google Scholar] [CrossRef]
- De Sousa, M. Immune cell functions in iron overload. Clin. Exp. Immunol. 1989, 75, 1. [Google Scholar] [PubMed]
- Rosen, G.M.; Pou, S.; Ramos, C.L.; Cohen, M.S.; Britigan, B.E. Free radicals and phagocytic cells. FASEB J. 1995, 9, 200–209. [Google Scholar] [CrossRef]
- Babitt, J.L.; Huang, F.W.; Wrighting, D.M.; Xia, Y.; Sidis, Y.; Samad, T.A.; Campagna, J.A.; Chung, R.T.; Schneyer, A.L.; Woolf, C.J. Bone morphogenetic protein signaling by hemojuvelin regulates hepcidin expression. Nat. Genet. 2006, 38, 531–539. [Google Scholar] [CrossRef]
- Omara, F.O.; Blakley, B.R. The effects of iron deficiency and iron overload on cell-mediated immunity in the mouse. Br. J. Nutr. 1994, 72, 899–909. [Google Scholar] [CrossRef]
- Valenzuela-Muñoz, V.; Boltaña, S.; Gallardo-Escárate, C. Comparative immunity of Salmo salar and Oncorhynchus kisutch during infestation with the sea louse Caligus rogercresseyi: An enrichment transcriptome analysis. Fish Shellfish Immunol. 2016, 59, 276–287. [Google Scholar] [CrossRef]
- Yamashita, M.; Harada, G.; Matsumoto, S.-e.; Aiba, Y.; Ichikawa, A.; Fujiki, T.; Udono, M.; Kabayama, S.; Yoshida, T.; Zhang, P. Suppression of immunoglobulin production in human peripheral blood mononuclear cells by monocytes via secretion of heavy-chain ferritin. Immunobiology 2014, 219, 149–157. [Google Scholar] [CrossRef]
- Feng, C.Y.; Johnson, S.C.; Hori, T.S.; Rise, M.; Hall, J.R.; Gamperl, A.K.; Hubert, S.; Kimball, J.; Bowman, S.; Rise, M.L. Identification and analysis of differentially expressed genes in immune tissues of Atlantic cod stimulated with formalin-killed, atypical Aeromonas salmonicida. Physiol. Genom. 2009, 37, 149–163. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Blaney, S.C.; Houlihan, D.; Secombes, C.J. Transcriptome response following administration of a live bacterial vaccine in Atlantic salmon (Salmo salar). Mol. Immunol. 2006, 43, 1900–1911. [Google Scholar] [CrossRef] [PubMed]
- Vanya Ewart, K.; Belanger, J.C.; Williams, J.; Karakach, T.; Penny, S.; Tsoi, S.C.M.; Richards, R.C.; Douglas, S.E. Identification of genes differentially expressed in Atlantic salmon (Salmo salar) in response to infection by Aeromonas salmonicida using cDNA microarray technology. Dev. Comp. Immunol. 2005, 29, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Chart, H. Acquisition of iron by Aeromonas salmonicida. J. Bacteriol. 1983, 156, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Rozas-Serri, M.; Peña, A.; Maldonado, L. Gene expression associated with immune response in Atlantic salmon head- kidney vaccinated with inactivated whole-cell bacterin of Piscirickettsia salmonis and pathogenic isolates. Fish Shellfish Immunol. 2019, 93, 789–795. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene | Sequence 5′-3′ | Tm | Efficiency (%) |
|---|---|---|---|---|
| EF1_F2 | Elongation factor (reference gene) | TGCTGGTGGTGTGGGTGAGT | 60 | 95.18 |
| EF1_R2 | CCTCAAACCGCTTCTGGCTGT | |||
| IgM_F | Immunoglobulin M | TGAGGAGAACTGTGGGCTACACT | 60 | 99.98 |
| IgM_R | TGTTAATGACCACTGAATGTGCAT | |||
| IgM_Probe | CATCAGATGCAGGTCC | |||
| IgT_F | Immunoglobulin T | CAACACTGACTGGAACAACAAGGT | 60 | 107.77 |
| IgT_R | CGTCAGCGGTTCTGTTTTGGA | |||
| IgT_Probe | AGTACAGCTGTGTGGTGCA | |||
| IL1b_F6 | Interleukin-1b | GATCTGGAGGTATCCCATCA | 60 | 122.36 |
| IL1b _R6 | CACAGCACTCTCCAGCAAGA | |||
| COX-2_F | Cyclooxygenase 2 | CAGTGCTCCCAGATGCCAAG | 60 | 102.34 |
| COX-2_R | GCGAAGAAGGCGAACATGAG | |||
| TLR22_3_F | Toll-like receptor 22 | TGCCTTCTAACCCTCTCCCT | 61 | 91.36 |
| TLR22_3_R | CGCTGCTCTCAGACAGGAAG | |||
| MHCII_F | Major histocompatibility complex II | CTCCTCAAAGGACCTGCAGG | 60 | 104.72 |
| MHCII_R | TCAGGACCTTTGTTCCAGGC | |||
| GSHPx_F1 | Glutathione peroxidase | TAAAGTGGTGCTGATCGAGA | 54 | 100.23 |
| GSHPx_R1 | GTTCTCCTGATGTCCGAACT | |||
| SOD_F1 | Superoxide dismutase | CCGTATTCTTTGAGCAGGAG | 54 | 104.98 |
| SOD_R1 | AGCCGTTGGTGTTGTCTC | |||
| ALA d_F2 | Aminolevulinate dehydratase | CCACTCGCCCATCCATCATA | 59 | 126.72 |
| ALA d_R2 | ACACCTCACATGGACACTGT | |||
| ALA s_F1 | Aminolevulinate synthase | GGTAGGATGCCTGCTGACTG | 63 | 105.52 |
| ALA s_R1 | CCCCAAGCCTGTTTTGCTGA | |||
| HO_F2 | Heme oxygenase | GTCCTCTCGAGTGGTGAAGC | 61 | 94.21 |
| HO_R2 | ATCTCTGAGTCCCTGGCCAA | |||
| Blv r_F1 | Biliverdin reductase | AAACAGATCCACCAGCCAGG | 59 | 106.7 |
| Blv r_R1 | ACAGCCGACTTTAAGCAGCT | |||
| Hep_F1 | Hepcidin | GCTGTTCCTTTCTCCGAGGTGC | 59 | 111.07 |
| Hep_R1 | GTGACAGCAGTTGCAGCACCA | |||
| FerritinM_F1 | Ferritin M | TATCACCACGATTGCGAAGC | 60 | 102.66 |
| FerritinM_R1 | CTCGTCGCTGTTCTCCTTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenzuela-Muñoz, V.; Benavente, B.P.; Casuso, A.; Leal, Y.; Gallardo-Escárate, C. Chimeric Protein IPath® with Chelating Activity Improves Atlantic Salmon’s Immunity against Infectious Diseases. Vaccines 2021, 9, 361. https://doi.org/10.3390/vaccines9040361
Valenzuela-Muñoz V, Benavente BP, Casuso A, Leal Y, Gallardo-Escárate C. Chimeric Protein IPath® with Chelating Activity Improves Atlantic Salmon’s Immunity against Infectious Diseases. Vaccines. 2021; 9(4):361. https://doi.org/10.3390/vaccines9040361
Chicago/Turabian StyleValenzuela-Muñoz, Valentina, Bárbara P. Benavente, Antonio Casuso, Yeny Leal, and Cristian Gallardo-Escárate. 2021. "Chimeric Protein IPath® with Chelating Activity Improves Atlantic Salmon’s Immunity against Infectious Diseases" Vaccines 9, no. 4: 361. https://doi.org/10.3390/vaccines9040361
APA StyleValenzuela-Muñoz, V., Benavente, B. P., Casuso, A., Leal, Y., & Gallardo-Escárate, C. (2021). Chimeric Protein IPath® with Chelating Activity Improves Atlantic Salmon’s Immunity against Infectious Diseases. Vaccines, 9(4), 361. https://doi.org/10.3390/vaccines9040361