
Pneumococcal Competition Modulates Antibiotic Resistance in the Pre-Vaccination Era: A Modelling Study



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
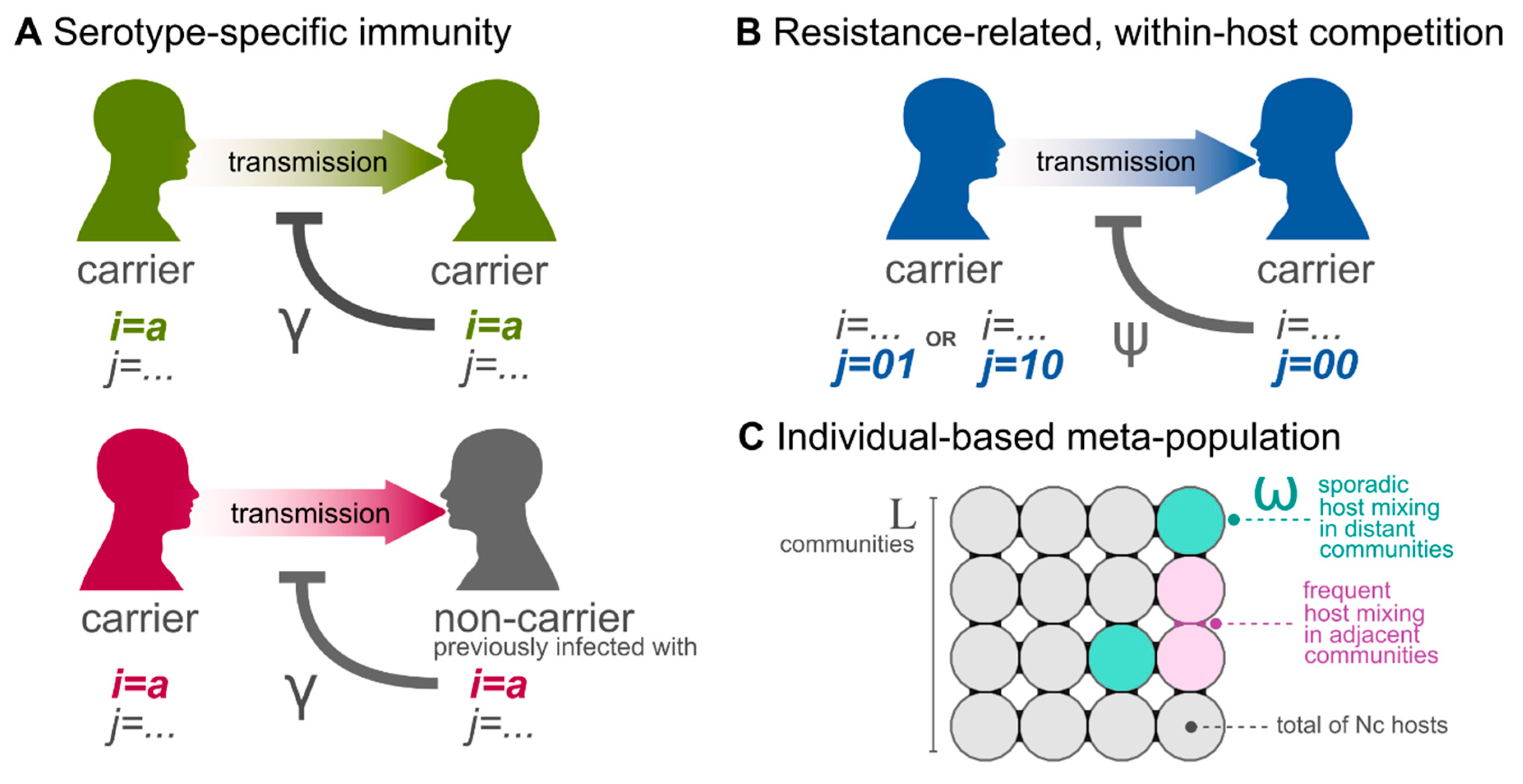
2.1. Epidemiological Framework
2.2. Antibiotic Resistance and Consumption Data
2.3. Approximate Bayesian Computation
3. Results
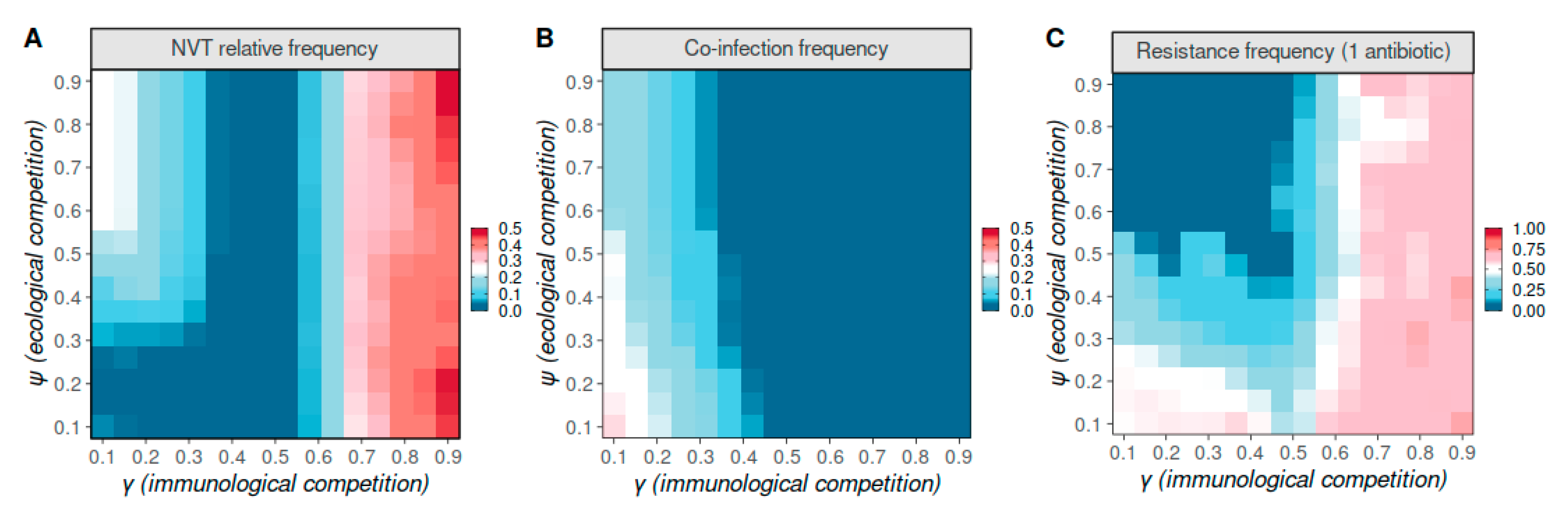
3.1. Pneumococcal Strain Dynamics
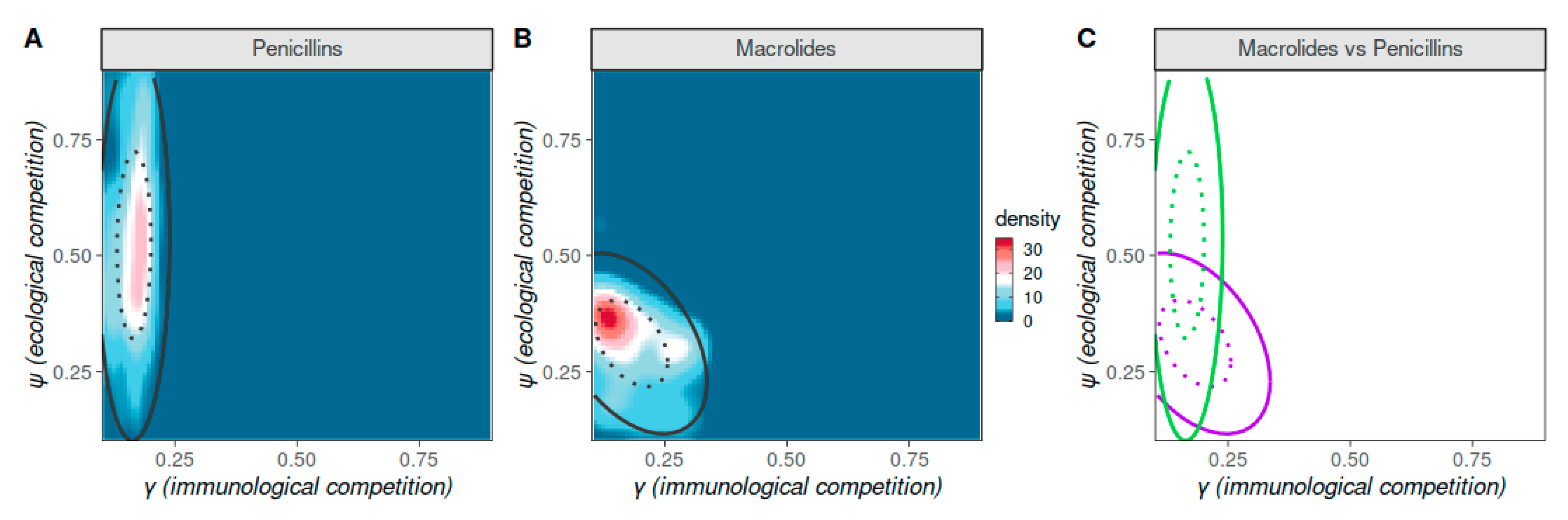
3.2. Strain Competition as Determinants of Antibiotic Resistance
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Disclaimer
Conflicts of Interest
References
- O’Brien, K.L.; Wolfson, L.J.; Watt, J.P.; Henkle, E.; Deloria-Knoll, M.; McCall, N.; Lee, E.; Mulholland, K.; Levine, O.S.; Cherian, T. Burden of disease caused by Streptococcus pneumoniae in children younger than 5 years: Global estimates. Lancet 2009, 374, 893–902. [Google Scholar] [CrossRef]
- Simell, B.; Auranen, K.; Käyhty, H.; Goldblatt, D.; Dagan, R.; O’Brien, K.L.; for the Pneumococcal Carriage Group. The fundamental link between pneumococcal car-riage and disease. Expert Rev. Vaccines 2012, 11, 841–855. [Google Scholar] [CrossRef]
- Tsaban, G.; Ben-Shimol, S. Indirect (herd) protection, following pneumococcal conjugated vaccines introduction: A systematic review of the literature. Vaccine 2017, 35, 2882–2891. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.H.; Camacho, L.A.B.; Coutinho, E.S.F.; Martinez-Silveira, M.S.; Carvalho, A.F.; Ruiz-Matus, C.; Toscano, C.M. Impact and Effec-tiveness of 10 and 13-Valent Pneumococcal Conjugate Vaccines on Hospitalization and Mortality in Children Aged Less than 5 Years in Latin American Countries: A Systematic Review. PLoS ONE 2016, 11, e0166736. [Google Scholar] [CrossRef] [PubMed]
- Hausdorff, W.P.; Feikin, D.R.; Klugman, K.P. Epidemiological differences among pneumococcal serotypes. Lancet Infect. Dis. 2005, 5, 83–93. [Google Scholar] [CrossRef]
- Bentley, S.D.; Aanensen, D.M.; Mavroidi, A.; Saunders, D.; Rabbinowitsch, E.; Collins, M.; Donohoe, K.; Harris, D.; Murphy, L.; Quail, M.A.; et al. Genetic Analysis of the Capsular Biosynthetic Locus from All 90 Pneumococcal Serotypes. PLoS Genet. 2006, 2, e31. [Google Scholar] [CrossRef] [PubMed]
- Flasche, S.; Van Hoek, A.J.; Sheasby, E.; Waight, P.; Andrews, N.; Sheppard, C.; George, R.; Miller, E. Effect of Pneumococcal Conjugate Vaccination on Serotype-Specific Carriage and Invasive Disease in England: A Cross-Sectional Study. PLoS Med. 2011, 8, e1001017. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.R.; Link-Gelles, R.; Schaffner, W.; Lynfield, R.; Lexau, C.; Bennett, N.M.; Petit, S.; Zansky, S.M.; Harrison, L.H.; Reingold, A.; et al. Effect of use of 13-valent pneumococcal con-jugate vaccine in children on invasive pneumococcal disease in children and adults in the USA: Analysis of multisite, popula-tion-based surveillance. Lancet Infect. Dis. 2015, 15, 301–309. [Google Scholar] [CrossRef]
- Gidding, H.F.; Sheridan, S.; Fathima, P.; Moore, H.C.; Liu, B.; McIntyre, P.B.; Palmu, A.A. Impact of Childhood Pneumococcal Conjugate Vaccine on Nonnotified Clinically Suspected Invasive Pneumococcal Disease in Australia. Pediatr. Infect. Dis. J. 2019, 38, 860–865. [Google Scholar] [CrossRef]
- Lourenço, J.; Obolski, U.; Swarthout, T.D.; Gori, A.; Bar-Zeev, N.; Everett, D.; Kamng’Ona, A.W.; Mwalukomo, T.S.; Mataya, A.A.; Mwansambo, C.; et al. Determinants of high residual post-PCV13 pneumococcal vaccine-type carriage in Blantyre, Malawi: A modelling study. BMC Med. 2019, 17, 219. [Google Scholar] [CrossRef]
- Swarthout, T.D.; Fronterre, C.; Lourenço, J.; Obolski, U.; Gori, A.; Bar-Zeev, N.; Everett, D.; Kamng’Ona, A.W.; Mwalukomo, T.S.; Mataya, A.A.; et al. High residual carriage of vaccine-serotype Streptococcus pneumoniae after introduction of pneumococcal conjugate vaccine in Malawi. Nat. Commun. 2020, 11, 2222. [Google Scholar] [CrossRef]
- Moore, M.R.; Gertz, R.E., Jr.; Woodbury, R.L.; Barkocy-Gallagher, G.A.; Schaffner, W.; Lexau, C.; Gershman, K.; Reingold, A.; Farley, M.; Harrison, L.H.; et al. Population Snapshot of EmergentStreptococcus pneumoniaeSerotype 19A in the United States, 2005. J. Infect. Dis. 2008, 197, 1016–1027. [Google Scholar] [CrossRef]
- Hanage, W.P.; Huang, S.S.; Lipsitch, M.; Bishop, C.J.; Godoy, D.; Pelton, S.I.; Goldstein, R.; Huot, H.; Finkelstein, J.A. Diversity and antibiotic resistance among nonvac-cine serotypes of Streptococcus pneumoniae carriage isolates in the post-heptavalent conjugate vaccine era. J. Infect. Dis. 2007, 195, 347–352. [Google Scholar] [CrossRef]
- Kyaw, M.H.; Lynfield, R.; Schaffner, W.; Craig, A.S.; Hadler, J.; Reingold, A.; Thomas, A.R.; Harrison, L.H.; Bennett, N.M.; Farley, M.M.; et al. Effect of introduction of the pneumococcal conju-gate vaccine on drug-resistant Streptococcus pneumoniae. N. Engl. J. Med. 2006, 354, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Moore, M.R.; Bruden, D.L.; Hyde, T.B.; Reasonover, A.L.; Harker-Jones, M.; Rudolph, K.M.; Hurlburt, D.A.; Parks, D.J.; Parkinson, A.J. Impact of conjugate vaccine on transmis-sion of antimicrobial-resistant Streptococcus pneumoniae among Alaskan children. Pediatr. Infect. Dis. J. 2008, 27, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Platt, R.; Rifas-Shiman, S.L.; Pelton, S.I.; Goldmann, D.; Finkelstein, J.A. Post-PCV7 changes in colonizing pneumococcal serotypes in 16 Massachusetts communities, 2001 and 2004. Pediatrics 2005, 116, e408–e413. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, G.; D’Ambrosio, F.; Visaggio, D.; Dicuonzo, G.; Del Grosso, M.; Pantosti, A. Serotype and Clonal Evolution of Penicillin-Nonsusceptible Invasive Streptococcus pneumoniae in the 7-Valent Pneumococcal Conjugate Vaccine Era in Italy. Antimicrob. Agents Chemother. 2012, 56, 4965–4968. [Google Scholar] [CrossRef] [PubMed]
- Farrell, D.J.; Klugman, K.P.; Pichichero, M. Increased Antimicrobial Resistance among Nonvaccine Serotypes of Streptococcus pneumoniae in the Pediatric Population After the Introduction of 7-Valent Pneumococcal Vaccine in the United States. Pediatr. Infect. Dis. J. 2007, 26, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Pillai, D.R.; Shahinas, D.; Buzina, A.; A Pollock, R.; Lau, R.; Khairnar, K.; Wong, A.; Farrell, D.J.; Green, K.; McGeer, A.; et al. Genome-wide dissection of globally emergent multi-drug resistant serotype 19A Streptococcus pneumoniae. BMC Genom. 2009, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M. Vaccination against colonizing bacteria with multiple serotypes. Proc. Natl. Acad. Sci. USA 1997, 94, 6571–6576. [Google Scholar] [CrossRef]
- Cobey, S.; Lipsitch, M. Niche and Neutral Effects of Acquired Immunity Permit Coexistence of Pneumococcal Serotypes. Science 2012, 335, 1376–1380. [Google Scholar] [CrossRef]
- Bottomley, C.; Roca, A.; Hill, P.C.; Greenwood, B.; Isham, V. A mathematical model of serotype replacement in pneumococcal car-riage following vaccination. J. R. Soc. Interface 2013, 10, 20130786. [Google Scholar] [CrossRef]
- Flasche, S.; Edmunds, W.J.; Miller, E.; Goldblatt, D.; Robertson, C.; Choi, Y.H. The impact of specific and non-specific immunity on the ecology of Streptococcus pneumoniae and the implications for vaccination. Proc. R. Soc. B Boil. Sci. 2013, 280, 20131939. [Google Scholar] [CrossRef] [PubMed]
- Melegaro, A.; Choi, Y.H.; George, R.; Edmunds, W.J.; Miller, E.; Gay, N.J. Dynamic models of pneumococcal carriage and the impact of the Heptavalent Pneumococcal Conjugate Vaccine on invasive pneumococcal disease. BMC Infect. Dis. 2010, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Beall, B.W.; Gertz, R.E.; Hulkower, R.L.; Whitney, C.G.; Moore, M.R.; Brueggemann, A.B. Shifting Genetic Structure of Invasive Serotype 19A Pneumococci in the United States. J. Infect. Dis. 2011, 203, 1360–1368. [Google Scholar] [CrossRef] [PubMed]
- Croucher, N.J.; Finkelstein, J.A.; Pelton, S.I.; Mitchell, P.K.; Lee, G.M.; Parkhill, J.; Bentley, S.D.; Hanage, W.P.; Lipsitch, M. Population genomics of post-vaccine changes in pneumococcal epidemiology. Nat. Genet. 2013, 45, 656–663. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Ferguson, N.M.; Anderson, R.M. Vaccination and the population structure of antigenically diverse pathogens that ex-change genetic material. Proc. Biol. Sci. 1997, 264, 1435–1443. [Google Scholar] [CrossRef]
- Watkins, E.R.; Penman, B.S.; Lourenço, J.; Buckee, C.O.; Maiden, M.C.J.; Gupta, S. Vaccination Drives Changes in Metabolic and Viru-lence Profiles of Streptococcus pneumoniae. PLoS Pathog. 2015, 11, e1005034. [Google Scholar] [CrossRef]
- Lourenço, J.; Watkins, E.R.; Obolski, U.; Peacock, S.J.; Morris, C.; Maiden, M.C.J.; Gupta, S. Lineage structure of Streptococcus pneumoniae may be driven by immune selection on the groEL heat-shock protein. Sci. Rep. 2017, 7, 9023. [Google Scholar] [CrossRef]
- Parks, T.; Barrett, L.; Jones, N. Invasive streptococcal disease: A review for clinicians. Br. Med Bull. 2015, 115, 77–89. [Google Scholar] [CrossRef]
- Dagan, R.; Givon-Lavi, N.; Zamir, O.; Fraser, D. Effect of a nonavalent conjugate vaccine on carriage of antibiotic-resistant Strep-tococcus pneumoniae in day-care centers. Pediatr. Infect. Dis. J. 2003, 22, 532–540. [Google Scholar] [CrossRef]
- Mbelle, N.; Huebner, R.E.; Wasas, A.D.; Kimura, A.; Chang, I.; Klugman, K.P. Immunogenicity and impact on nasopharyngeal car-riage of a nonavalent pneumococcal conjugate vaccine. J. Infect. Dis. 1999, 180, 1171–1176. [Google Scholar] [CrossRef]
- Mishra, R.P.N.; Oviedo-Orta, E.; Prachi, P.; Rappuoli, R.; Bagnoli, F. Vaccines and antibiotic resistance. Curr. Opin. Microbiol. 2012, 15, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Chiba, N.; Morozumi, M.; Shouji, M.; Wajima, T.; Iwata, S.; Sunakawa, K.; Ubukata, A.; Invasive Pneumococcal Diseases Surveillance Study Group. Rapid Decrease of 7-Valent Conjugate Vaccine Coverage for Invasive Pneumococcal Diseases in Pediatric Patients in Japan. Microb. Drug Resist. 2013, 19, 308–315. [Google Scholar] [CrossRef]
- Dagan, R.; Klugman, K.P. Impact of conjugate pneumococcal vaccines on antibiotic resistance. Lancet Infect. Dis. 2008, 8, 785–795. [Google Scholar] [CrossRef]
- Danino, D.; Givon-Lavi, N.; Ben-Shimol, S.; Greenberg, D.; Dagan, R. Understanding the Evolution of Antibiotic-nonsusceptible Pneumococcal Nasopharyngeal Colonization Following Pneumococcal Conjugate Vaccine Implementation in Young Children. Clin. Infect. Dis. 2019, 69, 648–656. [Google Scholar] [CrossRef]
- Obolski, U.; Lourenço, J.; Thompson, C.; Thompson, R.; Gori, A.; Gupta, S. Vaccination can drive an increase in frequencies of anti-biotic resistance among nonvaccine serotypes of. Proc. Natl. Acad. Sci. USA 2018, 115, 3102–3107. [Google Scholar] [CrossRef]
- Reinert, R. The antimicrobial resistance profile of Streptococcus pneumoniae. Clin. Microbiol. Infect. 2009, 15, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Annual Epidemiological Report 2009 [2007 Data]. European Centre for Disease Control [Internet]. 12 October 2009. Available online: https://www.ecdc.europa.eu/en/publications-data/annual-epidemiological-report-2009-2007-data (accessed on 21 September 2020).
- Willem, L.; Verelst, F.; Bilcke, J.; Hens, N.; Beutels, P. Lessons from a decade of individual-based models for infectious disease transmission: A systematic review (2006–2015). BMC Infect. Dis. 2017, 17, 612. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, J.; Recker, M. Natural, Persistent Oscillations in a Spatial Multi-Strain Disease System with Application to Dengue. PLoS Comput. Biol. 2013, 9, e1003308. [Google Scholar] [CrossRef]
- Lourenço, J.; Recker, M. Dengue serotype immune-interactions and their consequences for vaccine impact predictions. Epidemics 2016, 16, 40–48. [Google Scholar] [CrossRef]
- Flasche, S.; Jit, M.; Rodríguez-Barraquer, I.; Coudeville, L.; Recker, M.; Koelle, K.; Milne, G.; Hladish, T.J.; Perkins, T.A.; Cummings, D.A.T.; et al. The Long-Term Safety, Public Health Impact, and Cost-Effectiveness of Routine Vaccination with a Recombinant, Live-Attenuated Dengue Vaccine (Dengvaxia): A Model Comparison Study. PLoS Med. 2016, 13, e1002181. [Google Scholar] [CrossRef]
- European Centre for Disease Control (ECDC) Antibiotic Resistance Data. ECDC [Internet]. Available online: https://www.ecdc.europa.eu/en/antimicrobial-resistance (accessed on 22 July 2020).
- Minter, A.; Retkute, R. Approximate Bayesian Computation for infectious disease modelling. Epidemics 2019, 29, 100368. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, S.; Karimi, A.; Fallah, F.; Shiva, F.; Shamshiri, M.; Gooya, M.; Zahraei, M. Rate of co-colonization with serotypes of strep pneumonia isolated from nasopharyngeal swab. Int. J. Infect. Dis. 2014, 21, 418. [Google Scholar] [CrossRef][Green Version]
- Kamng’Ona, A.W.; Hinds, J.; Bar-Zeev, N.; Gould, K.A.; Chaguza, C.; Msefula, C.; Cornick, J.E.; Kulohoma, B.W.; Gray, K.; Bentley, S.D.; et al. High multiple carriage and emergence of Streptococcus pneumoniae vaccine serotype variants in Malawian children. BMC Infect. Dis. 2015, 15, 234. [Google Scholar] [CrossRef]
- Melegaro, A.; Choi, Y.; Pebody, R.; Gay, N. Pneumococcal Carriage in United Kingdom Families: Estimating Serotype-specific Transmission Parameters from Longitudinal Data. Am. J. Epidemiol. 2007, 166, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Melegaro, A.; Gay, N.J.; Medley, G.F. Estimating the transmission parameters of pneumococcal carriage in households. Epidemiol. Infect. 2004, 132, 433–441. [Google Scholar] [CrossRef]
- Lipsitch, M.; Abdullahi, O.; DʼAmour, A.; Xie, W.; Weinberger, D.M.; Tchetgen Tchetgen, E.; Scott, J.A.G. Estimating rates of carriage acqui-sition and clearance and competitive ability for pneumococcal serotypes in Kenya with a Markov transition model. Epidemiology 2012, 23, 510–519. [Google Scholar] [CrossRef]
- Adetifa, I.M.O.; Antonio, M.; Okoromah, C.A.N.; Ebruke, C.; Inem, V.; Nsekpong, D.; Bojang, A.; Adegbola, R.A. Pre-Vaccination Nasopharyngeal Pneumococcal Carriage in a Nigerian Population: Epidemiology and Population Biology. PLoS ONE 2012, 7, e30548. [Google Scholar] [CrossRef]
- Heinsbroek, E.; Tafatatha, T.; Phiri, A.; Swarthout, T.D.; Alaerts, M.; Crampin, A.C.; Chisambo, C.; Mwiba, O.; Read, J.M.; French, N. Pneumococcal carriage in households in Karonga District, Malawi, before and after introduction of 13-valent pneumococcal conjugate vaccination. Vaccine 2018, 36, 7369–7376. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Gregson, J.; Parkin, N.; Haile-Selassie, H.; Tanuri, A.; Forero, L.A.; Kaleebu, P.; Watera, C.; Aghokeng, A.; Mutenda, N.; et al. HIV-1 drug resistance before initiation or re-initiation of first-line antiretroviral therapy in low-income and middle-income countries: A systematic review and meta-regression analysis. Lancet Infect. Dis. 2018, 18, 346–355. [Google Scholar] [CrossRef]
- Puertas, M.C.; Ploumidis, G.; Ploumidis, M.; Fumero, E.; Clotet, B.; Walworth, C.M.; Petropoulos, C.J.; Martinez-Picado, J. Pan-resistant HIV-1 emergence in the era of integrase strand-transfer inhibitors: A case report. Lancet Microbe 2020, 1, e130–e135. [Google Scholar] [CrossRef]
- Gao, B.; Saralamba, S.; Lubell, Y.; White, L.J.; Dondorp, A.M.; Aguas, R. Determinants of MDA impact and designing MDAs towards malaria elimination. eLife 2020, 9. [Google Scholar] [CrossRef]
- Kremer, K.; Van Der Werf, M.J.; Au, B.K.Y.; Anh, D.D.; Kam, K.M.; Van Doorn, H.R.; Borgdorff, M.W.; Van Soolingen, D. Vaccine-induced Immunity Circumvented by Typical Mycobacterium tuberculosis Beijing Strains. Emerg. Infect. Dis. 2009, 15, 335–339. [Google Scholar] [CrossRef]
- Austin, D.J.; Kristinsson, K.G.; Anderson, R.M. The relationship between the volume of antimicrobial consumption in human communities and the frequency of resistance. Proc. Natl. Acad. Sci. USA 1999, 96, 1152–1156. [Google Scholar] [CrossRef]
- Lehtinen, S.; Blanquart, F.; Croucher, N.J.; Turner, P.; Lipsitch, M.; Fraser, C. Evolution of antibiotic resistance is linked to any genet-ic mechanism affecting bacterial duration of carriage. Proc. Natl. Acad. Sci. USA 2017, 114, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Colijn, C.; Cohen, T.; Fraser, C.; Hanage, W.; Goldstein, E.; Givon-Lavi, N.; Dagan, R.; Lipsitch, M. What is the mechanism for persistent coexistence of drug-susceptible and drug-resistant strains of Streptococcus pneumoniae? J. R. Soc. Interface 2009, 7, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Cobey, S.; Baskerville, E.B.; Colijn, C.; Hanage, W.; Fraser, C.; Lipsitch, M. Host population structure and treatment frequency maintain balancing selection on drug resistance. J. R. Soc. Interface 2017, 14, 20170295. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Beggs, J.J.; Walsh, T.R.; Gandra, S.; Laxminarayan, R. Anthropological and socioeconomic factors contributing to global antimicrobial resistance: A univariate and multivariable analysis. Lancet Planet. Health 2018, 2, e398–e405. [Google Scholar] [CrossRef]
- Blanquart, F.; Lehtinen, S.; Fraser, C. An evolutionary model to predict the frequency of antibiotic resistance under seasonal antibiotic use, and an application to Streptococcus pneumoniae. Proc. R. Soc. B Boil. Sci. 2017, 284, 20170679. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.G.; Flasche, S.; Jit, M.; Atkins, K.E. Within-host dynamics shape antibiotic resistance in commensal bacteria. Nat. Ecol. Evol. 2019, 3, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M.; Colijn, C.; Cohen, T.; Hanage, W.P.; Fraser, C. No coexistence for free: Neutral null models for multistrain pathogens. Epidemics 2009, 1, 2–13. [Google Scholar] [CrossRef]
- Melnyk, A.H.; Wong, A.; Kassen, R. The fitness costs of antibiotic resistance mutations. Evol. Appl. 2015, 8, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Janoir, C.; Cohen, R.; Levy, C.; Bingen, E.; Lepoutre, A.; Gutmann, L.; Varon, E.; the Observatoires Régionaux du Pneumocoque (ORP) Network. Clonal Expansion of the Macrolide Resistant ST386 within Pneumococcal Serotype 6C in France. PLoS ONE 2014, 9, e90935. [Google Scholar] [CrossRef] [PubMed]
- Keenan, J.D.; Klugman, K.P.; McGee, L.; Vidal, J.E.; Chochua, S.; Hawkins, P.; Cevallos, V.; Gebre, T.; Tadesse, Z.; Emerson, P.M.; et al. Evidence for clonal expansion after antibiotic se-lection pressure: Pneumococcal multilocus sequence types before and after mass azithromycin treatments. J. Infect. Dis. 2015, 211, 988–994. [Google Scholar] [CrossRef] [PubMed]
- MacFadden, D.R.; McGough, S.F.; Fisman, D.; Santillana, M.; Brownstein, J.S. Antibiotic Resistance Increases with Local Tempera-ture. Nat. Clim. Chang. 2018, 8, 510–514. [Google Scholar] [CrossRef]
- Bruinsma, N.; Hutchinson, J.M.; van den Bogaard, A.E.; Giamarellou, H.; Degener, J.; Stobberingh, E.E. Influence of population densi-ty on antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 385–390. [Google Scholar] [CrossRef]
- Blommaert, A.; Marais, C.; Hens, N.; Coenen, S.; Muller, A.; Goossens, H.; Beutels, P. Determinants of between-country differences in ambulatory antibiotic use and antibiotic resistance in Europe: A longitudinal observational study. J. Antimicrob. Chemother. 2013, 69, 535–547. [Google Scholar] [CrossRef]
- Brown, J.; Hammerschmidt, S.; Orihuela, C. Streptococcus Pneumoniae: Molecular Mechanisms of Host-Pathogen Interactions; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Jefferson, T.; Ferroni, E.; Curtale, F.; Rossi, P.G.; Borgia, P. Streptococcus pneumoniae in western Europe: Serotype distribution and incidence in children less than 2 years old. Lancet Infect. Dis. 2006, 6, 405–410. [Google Scholar] [CrossRef]
- Feikin, D.R.; Klugman, K.P. Historical Changes in Pneumococcal Serogroup Distribution: Implications for the Era of Pneumo-coccal Conjugate Vaccines. Clin. Infect. Dis. 2002, 35, 547–555. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lourenço, J.; Daon, Y.; Gori, A.; Obolski, U. Pneumococcal Competition Modulates Antibiotic Resistance in the Pre-Vaccination Era: A Modelling Study. Vaccines 2021, 9, 265. https://doi.org/10.3390/vaccines9030265
Lourenço J, Daon Y, Gori A, Obolski U. Pneumococcal Competition Modulates Antibiotic Resistance in the Pre-Vaccination Era: A Modelling Study. Vaccines. 2021; 9(3):265. https://doi.org/10.3390/vaccines9030265
Chicago/Turabian StyleLourenço, José, Yair Daon, Andrea Gori, and Uri Obolski. 2021. "Pneumococcal Competition Modulates Antibiotic Resistance in the Pre-Vaccination Era: A Modelling Study" Vaccines 9, no. 3: 265. https://doi.org/10.3390/vaccines9030265
APA StyleLourenço, J., Daon, Y., Gori, A., & Obolski, U. (2021). Pneumococcal Competition Modulates Antibiotic Resistance in the Pre-Vaccination Era: A Modelling Study. Vaccines, 9(3), 265. https://doi.org/10.3390/vaccines9030265