
Association between Venom Immunotherapy and Changes in Serum Protein—Peptide Patterns

 , ,
, ,
Abstract
1. Introduction
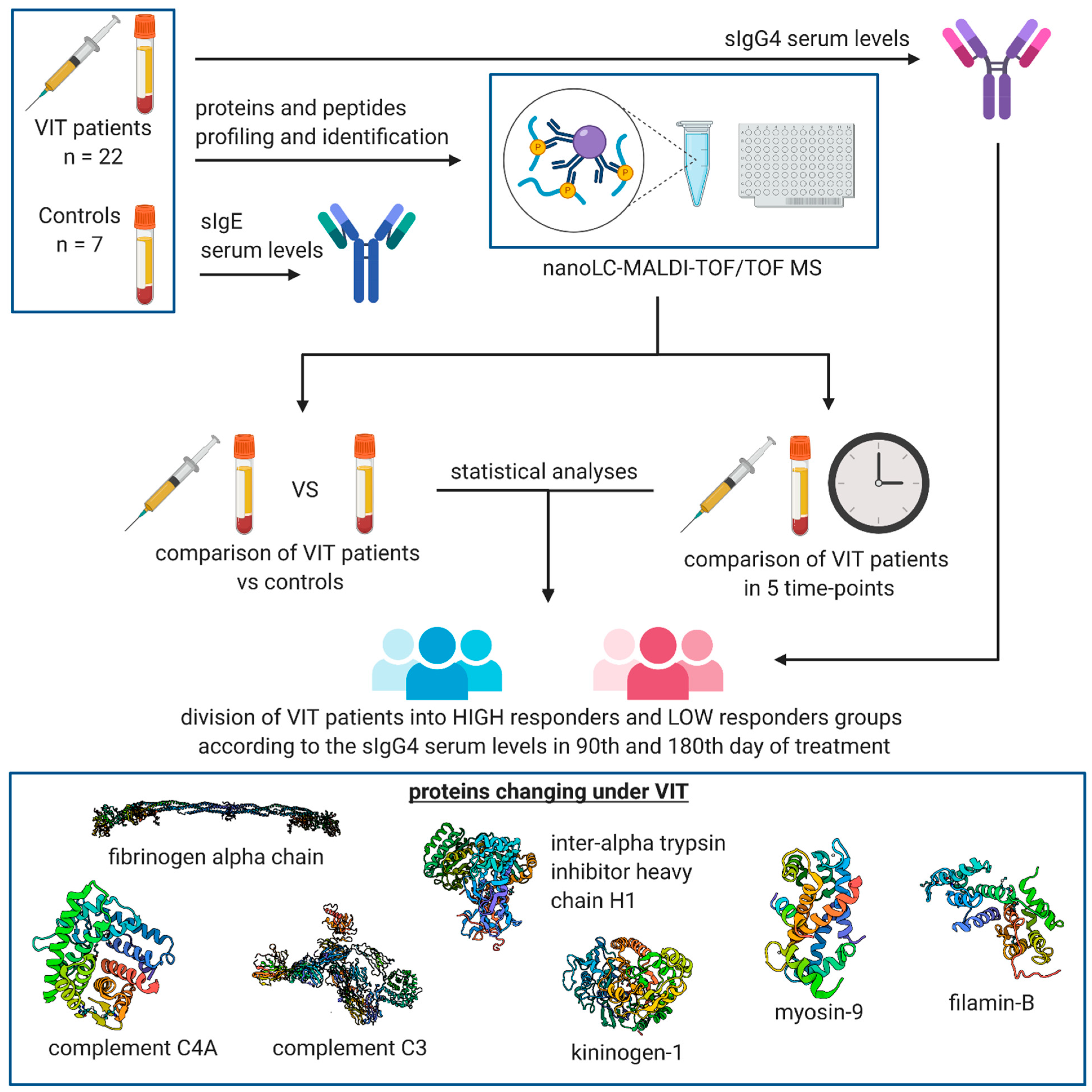
2. Materials and Methods
2.1. Characteristics of the Study Group
2.2. Measurement of sIgE and sIgG4 Serum Levels
2.3. Pre-Treatment of the Serum Samples
2.4. MALDI-TOF MS Proteomic Profiling
2.5. NanoLC-MALDI-TOF/TOF MS Identification of Discriminative Proteomic Features
2.6. Data Analysis
3. Results
3.1. Serum sIgG4 Levels before and on the 90th and 180th Days of VIT
3.2. MALDI-TOF MS Protein–Peptide Profiling and Identification of the Discriminatory Features
3.3. Comparison of Protein–Peptide Profiles Derived from Allergic Patients Undergoing VIT Classified as Low and High IgG4 Responder Groups
- The ratio of serum sIgG4 levels at two time-points < 15 (sIgG4 value of the 90th/180th day divided by the sIgG4 value of baseline)
- Differences between sIgG4 on the 90th/180th day and baseline < 6
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bilò, M.B.; Bonifazi, F. The natural history and epidemiology of insect venom allergy: Clinical implications. Clin. Exp. Allergy 2009, 39, 1467–1476. [Google Scholar] [CrossRef]
- Bilò, B.M.; Bonifazi, F. Epidemiology of insect-venom anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Incorvaia, C.; Mauro, M.; Gritti, B.L.; Makri, E.; Ridolo, E. Venom immunotherapy in patients with allergic reactions to insect stings. Expert Rev. Clin. Immunol. 2018, 14, 53–59. [Google Scholar] [CrossRef]
- Dhami, S.; Zaman, H.; Varga, E.-M.; Sturm, G.J.; Muraro, A.; Akdis, C.A.; Antolín-Amérigo, D.; Bilò, M.B.; Bokanovic, D.; Calderon, M.A.; et al. Allergen immunotherapy for insect venom allergy: A systematic review and meta-analysis. Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 342–365. [Google Scholar] [CrossRef]
- Kosnik, M.; Korosec, P. Venom immunotherapy: Clinical efficacy, safety and contraindications. Expert Rev. Clin. Immunol. 2015, 11, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Sturm, G.J.; Varga, E.M.; Roberts, G.; Mosbech, H.; Bilò, M.B.; Akdis, C.A.; Antolin-Amerigo, D.; Cichocka-Jarosz, E.; Gawlik, R.; Jakob, T.; et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy 2018, 73, 744–764. [Google Scholar] [CrossRef]
- Goldberg, A.; Confino-Cohen, R. Bee venom immunotherapy—How early is it effective? Allergy Eur. J. Allergy Clin. Immunol. 2010, 65, 391–395. [Google Scholar] [CrossRef]
- Heneberg, P.; Riegerová, K.; Kučera, P. Pimecrolimus Is a Potent Inhibitor of Allergic Reactions to Hymenopteran Venom Ex-tracts and Birch Pollen Allergen In Vitro. PLoS ONE 2015, 10, e0142953. [Google Scholar] [CrossRef] [PubMed]
- Heneberg, P.; Riegerová, K.; Říhová, A.; Šimčíková, D.; Kučera, P. Updates on the surface antigens of basophils: CD16 on baso-phils of patients with respiratory or insect venom allergy and the rejection of CD203c and CD63 externalization decoupling by bisindolylmaleimides. Clin. Exp. Allergy 2019, 49, 54–67. [Google Scholar] [CrossRef]
- Rodríguez Trabado, A.; Cámara Hijón, C.; Ramos Cantariño, A.; Romero-Chala, S.; García-Trujillo, J.A.; Fernández Pereira, L.M. Short-, Intermediate-, and Long-Term Changes in Basophil Reactivity Induced by Venom Immunotherapy. Allergy Asthma Immunol. Res. 2016, 8, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Žitnik, S.E.K.; Vesel, T.; Avčin, T.; Šilar, M.; Košnik, M.; Korošec, P. Monitoring honeybee venom immunotherapy in children with the basophil activation test. Pediatr. Allergy Immunol. 2012, 23, 167–172. [Google Scholar] [CrossRef]
- Ebo, D.G.; Hagendorens, M.M.; Schuerwegh, A.J.; Beirens, L.M.-N.; Bridts, C.H.; De Clerck, L.S.; Stevens, W.J. Flow-assisted quantification of in vitro activated basophils in the diagnosis of wasp venom allergy and follow-up of wasp venom immunotherapy. Cytom. Part B Clin. Cytom. 2007, 72, 196–203. [Google Scholar] [CrossRef]
- Bidad, K.; Nawijn, M.C.; van Oosterhout, A.J.M.; van der Heide, S.; Oude Elberink, J.N.G. Basophil activation test in the diagnosis and monitoring of mastocytosis patients with wasp venom allergy on immunotherapy. Cytom. Part B Clin. Cytom. 2014, 86, 183–190. [Google Scholar] [CrossRef]
- Eržen, R.; Košnik, M.; Šilar, M.; Korošec, P. Basophil response and the induction of a tolerance in venom immunotherapy: A long-term sting challenge study. Allergy 2012, 67, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Lerch, E.; Müller, U.R. Long-term protection after stopping venom immunotherapy: Results of re-stings in 200 patients. J. Allergy Clin. Immunol. 1998, 101, 606–612. [Google Scholar] [CrossRef]
- Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy: Multiple suppressor factors at work in immune toler-ance to allergens. J. Allergy Clin. Immunol. 2014, 133, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Ozdemir, C.; Kucuksezer, U.C.; Akdis, M.; Akdis, C.A. Mechanisms of immunotherapy to wasp and bee venom. Clin. Exp. Allergy 2011, 41, 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Graham, F.; Bégin, P.; Paradis, L.; Lacombe-Barrios, J.; Paradis, J.; Des Roches, A. Comparison of ImmunoCAP and Immulite serum specific IgE assays for the assessment of egg allergy. Allergy Asthma Clin. Immunol. 2016, 12, 129. [Google Scholar] [CrossRef]
- Bonifazi, F.; Jutel, M.; Bilo, B.M.; Birnbaum, J.; Muller, U. Prevention and treatment of hymenoptera venom allergy: Guidelines for clinical practice. Allergy 2005, 60, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Swiatly, A.; Horala, A.; Hajduk, J.; Matysiak, J.; Nowak-Markwitz, E.; Kokot, Z.J. MALDI-TOF-MS analysis in discovery and identification of serum proteomic patterns of ovarian cancer. BMC Cancer 2017, 17, 472. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wen, Q.; Li, Z.-J.; Xu, R.-C.; Peng, F.-F.; Yu, X.-Q. Optimization and evaluation of magnetic bead separation combined with matrix-assisted laser desorption/ionization time-of-flight mass spectroscopy (MALDI-TOF MS) for proteins profiling of peri-toneal dialysis effluent. Int. J. Mol. Sci. 2014, 15, 1162–1175. [Google Scholar] [CrossRef]
- Klupczynska, A.; Swiatly, A.; Hajduk, J.; Matysiak, J.; Dyszkiewicz, W.; Pawlak, K.; Kokot, Z.J. Identification of Serum Peptidome Signa-tures of Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2016, 17, 410. [Google Scholar] [CrossRef]
- Matuszewska, E.; Matysiak, J.; Kycler, Z.; Kokot, Z.J.; Matysiak, J. Proteomic features characterization of Hymenoptera venom allergy. Allergy Asthma Clin. Immunol. 2019, 15, 77. [Google Scholar] [CrossRef]
- Naik, R.S.; Wala, M.S. Inflammation, Allergy and Asthma, Complex Immune Origin Diseases: Mechanisms and Therapeutic Agents. Recent Pat. Inflamm. Allergy Drug. Discov. 2013, 7, 62–95. [Google Scholar] [CrossRef]
- Gabay, C.; Kushner, I. Acute-Phase Proteins and Other Systemic Responses to Inflammation. N. Engl. J. Med. 1999, 340, 448–454. [Google Scholar] [CrossRef]
- Smiley, S.T.; King, J.A.; Hancock, W.W. Fibrinogen Stimulates Macrophage Chemokine Secretion through Toll-Like Receptor 4. J. Immunol. 2001, 167, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Davalos, D.; Akassoglou, K. Fibrinogen as a key regulator of inflammation in disease. Semin. Immunopathol. 2012, 34, 43–62. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, J.Y.; Smith, T.D.; Meli, V.S.; Tran, T.N.; Botvinick, E.L.; Liu, W.F. Differential regulation of macrophage inflammatory activation by fibrin and fibrinogen. Acta Biomater. 2017, 47, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Rhim, T.; Choi, Y.-S.; Nam, B.-Y.; Uh, S.T.; Park, J.S.; Kim, Y.-H.; Paik, Y.-K.; Park, C.-S. Plasma protein profiles in early asthmatic responses to inhalation allergen challenge. Allergy Eur. J. Allergy Clin. Immunol. 2009, 64, 47–54. [Google Scholar] [CrossRef]
- Wang, H.; Gottfries, J.; Barrenäs, F.; Benson, M. Identification of novel biomarkers in seasonal allergic rhinitis by combining pro-teomic, multivariate and pathway analysis. PLoS ONE 2011, 6, e23563. [Google Scholar] [CrossRef]
- Całkosiński, I.; Dobrzyński, M.; Całkosińska, M.; Seweryn, E.; Bronowicka-Szydełko, A.; Dzierzba, K.; Ceremuga, I.; Gamian, A. Characterization of an in ammatory response. Postepy Hig. Med. Dosw. 2009, 63, 395–408. [Google Scholar]
- Pulani, D.; Rudan, I. The Past Decade: Fibrinogen. Coll. Antropol. 2005, 29, 341–349. [Google Scholar]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Kim, B.Y.; Sohn, M.R.; Roh, J.Y.; Je, Y.H.; Kim, N.J.; Kim, I.; Woo, S.D.; et al. Dual function of a bee venom serine protease: Prophenoloxi-dase-activating factor in arthropods and fibrin(ogen)olytic enzyme in mammals. PLoS ONE 2010, 5, e10393. [Google Scholar] [CrossRef]
- Matysiak, J.; Hajduk, J.; Mayer, F.; Hebeler, R.; Kokot, Z.J. Hyphenated LC–MALDI–ToF/ToF and LC–ESI–QToF approach in pro-teomic characterization of honeybee venom. J. Pharm. Biomed. Anal. 2016, 12, 69–76. [Google Scholar] [CrossRef]
- Matysiak, J.; Hajduk, J.; Pietrzak, Ł.; Schmelzer, C.E.; Kokot, Z.J. Shotgun proteome analysis of honeybee venom using targeted enrichment strategies. Toxicon 2014, 90, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Qiu, Y.; Wan, H.; Sohn, M.R.; Sohn, H.D.; Jin, B.R. Antifibrinolytic Role of a Bee Venom Serine Protease Inhibitor That Acts as a Plasmin Inhibitor. PLoS ONE 2012, 7, e32269. [Google Scholar] [CrossRef] [PubMed]
- Oikonomopoulou, K.; Ricklin, D.; Ward, P.A.; Lambris, J.D. Interactions between coagulation and complement—Their role in inflammation. Semin. Immunopathol. 2012, 34, 151–165. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Song, W.C. Complement and its role in innate and adaptive immune responses. Cell Res. 2010, 20, 34–50. [Google Scholar] [CrossRef]
- Markiewski, M.M.; Lambris, J.D. The Role of Complement in Inflammatory Diseases from Behind the Scenes into the Spotlight. Am. J. Pathol. 2007, 171, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Pidde-Queiroz, G.; de Fatima Furtado, M.; Filgueiras, C.F.; Pessoa, L.A.; Spadafora-Ferreira, M.; van den Berg, C.W.; Tambourgi, D.V. Human comple-ment activation and anaphylatoxins generation induced by snake venom toxins from Bothrops genus. Mol. Immunol. 2010, 47, 2537–2544. [Google Scholar] [CrossRef] [PubMed]
- Garantziotis, S.; Hollingsworth, J.W.; Ghanayem, R.B.; Timberlake, S.; Zhuo, L.; Kimata, K.; Schwartz, D.A. Inter-alpha-Trypsin Inhibitor Attenuates Complement Activation and Complement-Induced Lung Injury. J. Immunol. 2007, 179, 4187–4192. [Google Scholar] [CrossRef]
- Subbannayya, Y.; Mir, S.A.; Renuse, S.; Manda, S.S.; Pinto, S.M.; Puttamallesh, V.N.; Solanki, H.S.; Manju, H.; Syed, N.; Sharma, R.; et al. Identification of differentially expressed serum proteins in gastric adenocarcinoma. J. Proteom. 2015, 127, 80–88. [Google Scholar] [CrossRef]
- Cortelazzo, A.; Guerranti, R.; Bini, L.; Hope-Onyekwere, N.; Muzzi, C.; Leoncini, R.; Pagani, R. Effects of snake venom proteases on hu-man fibrinogen chains. Blood Transfus. 2010, 8 (Suppl. S3), S120–S125. [Google Scholar] [PubMed]
- Pandya, B.V.; Rubin, R.N.; Olexa, S.A.; Budzynski, A.Z. Unique degradation of human fibrinogen by proteases from western dia-mondback rattlesnake (Crotalus atrox) venom. Toxicon 1983, 21, 515–526. [Google Scholar] [CrossRef]
- Patra, A.; Kalita, B.; Chanda, A.; Mukherjee, A.K. Proteomics and antivenomics of Echis carinatus carinatus venom: Correlation with pharmacological properties and pathophysiology of envenomation. Sci. Rep. 2017, 7, 17119. [Google Scholar] [CrossRef] [PubMed]
- Galera, C.; Soohun, N.; Zankar, N.; Caimmi, S.; Gallen, C.; Demoly, P. Severe anaphylaxis to bee venom immunotherapy: Efficacy of pretreatment and concurrent treatment with omalizumab. J. Investig. Allergol. Clin. Immunol. 2009, 19, 225–229. [Google Scholar]
- Xu, Q.; Wu, N.; Cui, L.; Wu, Z.; Qiu, G. Filamin B: The next hotspot in skeletal research? J. Genet. Genom. 2017, 44, 335–342. [Google Scholar] [CrossRef]
- Baldassarre, M.; Razinia, Z.; Burande, C.F.; Lamsoul, I.; Lutz, P.G.; Calderwood, D.A. Filamins Regulate Cell Spreading and Initiation of Cell Migration. PLoS ONE 2009, 4, e7830. [Google Scholar] [CrossRef]
- Baltz, A.G.; Munschauer, M.; Schwanhäusser, B.; Vasile, A.; Murakawa, Y.; Schueler, M.; Youngs, N.; Penfold-Brown, D.; Drew, K.; Milek, M.; et al. The mRNA-Bound Proteome and Its Global Occupancy Profile on Protein-Coding Transcripts. Mol. Cell 2012, 46, 674–690. [Google Scholar] [CrossRef]
- Mattson, L.; Lentini, A.; Gawel, D.R.; Badam, T.V.S.; Benson, M.; Ledin, T.; Nestor, C.E.; Gustafsson, M.; Serra-Musach, J.; Bjorkander, J.; et al. Potential Involvement of Type I Interferon Signaling in Immunotherapy in Seasonal Allergic Rhinitis. J. Immunol. Res. 2016, 2016, 5153184. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, S.R.G.; Farrar, J.D. Interferon at the crossroads of allergy and viral infections. J. Leukoc. Biol. 2015, 98, 185–194. [Google Scholar] [CrossRef]
- Jeon, Y.J.; Choi, J.S.; Lee, J.Y.; Yu, K.R.; Ka, S.H.; Cho, Y.; Choi, E.-J.; Baek, S.H.; Seol, J.H.; Park, D.; et al. Filamin B Serves as a Molecular Scaffold for Type I Interferon-induced c-Jun NH2-terminal Kinase Signaling Pathway. Mol. Biol. Cell 2008, 19, 5116–5130. [Google Scholar] [CrossRef][Green Version]
- Jeon, Y.J.; Choi, J.S.; Lee, J.Y.; Yu, K.R.; Kim, S.M.; Ka, S.H.; Oh, K.H.; Kim, K.; Zhang, D.; Bang, O.S.; et al. ISG15 modification of filamin B negatively regulates the type I interferon-induced JNK signalling pathway. EMBO Rep. 2009, 10, 374–380. [Google Scholar] [CrossRef]
- Whitmarsh, A.J. Filamin B: A scaffold for interferon signalling. EMBO Rep. 2009, 10, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Narain, N.R.; Diers, A.R.; Lee, A.; Lao, S.; Chan, J.Y.; Schofield, S.; Andreazi, J.; Ouro-Djobo, R.; Jimenez, J.J.; Friss, T.; et al. Identification of Filamin-A and -B as potential biomarkers for prostate cancer. Futur. Sci. OA 2017, 3, FSO161. [Google Scholar] [CrossRef]
- Pecci, A.; Ma, X.; Savoia, A.; Adelstein, R.S. MYH9: Structure, functions and role of non-muscle myosin IIA in human disease. Gene 2018, 664, 152–167. [Google Scholar] [CrossRef]
- Vicente-Manzanares, M.; Ma, X.; Adelstein, R.S.; Horwitz, A.R. Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat. Rev. Mol. Cell Biol. 2009, 10, 778–790. [Google Scholar] [CrossRef]
- Heissler, S.M.; Manstein, D.J. Nonmuscle myosin-2: Mix and match. Cell. Mol. Life Sci. 2012, 70, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, R.; Chen, X.-X.; Zhi, Y.; Deng, R.; Zhou, E.-M.; Qiao, S.; Zhang, G. Nonmuscle Myosin Heavy Chain IIA Recognizes Sialic Acids on Si-alylated RNA Viruses to Suppress Proinflammatory Responses via the DAP12-Syk Pathway. mBio 2019, 10, e00574-19. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Xue, B.; Li, L.; Nan, Y.; Zhang, L.; Li, K.; Zhao, Q.; Hiscox, J.A.; Stewart, J.P.; Wu, C.; et al. Direct Interaction Between CD163 N-Terminal Domain and MYH9 C-Terminal Domain Contributes to Porcine Reproductive and Respiratory Syndrome Virus Internalization by Permissive Cells. Front. Microbiol. 2019, 10, 1815. [Google Scholar] [CrossRef]
- Högger, P.; Sorg, C. Soluble CD163 Inhibits Phorbol Ester-Induced Lymphocyte Proliferation. Biochem. Biophys. Res. Commun. 2001, 288, 841–843. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Lin, S.; Chen, C.; Wang, C.; Ma, Q.; Jiang, B. Identification of Kininogen-1 as a Serum Biomarker for the Early De-tection of Advanced Colorectal Adenoma and Colorectal Cancer. PLoS ONE 2013, 8, e70519. [Google Scholar] [CrossRef]
- Yousef, G.M.; Diamandis, E.P. The New Human Tissue Kallikrein Gene Family: Structure, Function, and Association to Disease. Endocr. Rev. 2001, 22, 184–204. [Google Scholar] [CrossRef]
- Golias, C.; Charalabopoulos, A.; Stagikas, D.; Charalabopoulos, K.; Batistatou, A. The kinin system-bradykinin: Biological effects and clinical implications. Multiple role of the kinin system-bradykinin. Hippokratia 2007, 11, 124–128. [Google Scholar]
- Couture, R.; Harrisson, M.; Vianna, R.M.; Cloutier, F. Kinin receptors in pain and inflammation. Eur. J. Pharmacol. 2001, 429, 161–176. [Google Scholar] [CrossRef]
- Lopes-Ferreira, M.; da Silva Emim, J.A.; Oliveira, V.; Puzer, L.; Cezari, M.H.; da Silva Araujo, M.; Juliano, L.; Jose Lapa, A.; Souccar, C.; Moura-da-Silva, A. Kininogenase activity of Thalassophryne nattereri fish venom. Biochem. Pharmacol. 2004, 68, 2151–2157. [Google Scholar] [CrossRef]
- Vaiyapuri, S.; Harrison, R.A.; Bicknell, A.B.; Gibbins, J.M.; Hutchinson, G. Purification and Functional Characterisation of Rhinocerase, a Novel Serine Protease from the Venom of Bitis gabonica rhinoceros. PLoS ONE 2010, 5, e9687. [Google Scholar] [CrossRef]
- Bilò, M.B.; Martini, M.; Corsi, A.; Tontini, C.; Antonicelli, L. Venom immunotherapy in Europe and the United States. Allergo J. Int. 2020, 29, 29–37. [Google Scholar] [CrossRef]
- Tracy, J.M.; Golden, D.B. Hymenoptera Venom Extracts in Clinical Practice. J. Allergy Clin. Immunol. Pr. 2018, 6, 1856–1862. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef]
- Boyle, R.J.; Elremeli, M.; Hockenhull, J.; Cherry, M.G.; Bulsara, M.K.; Daniels, M.; Elberink, J.O. Venom immunotherapy for preventing allergic reactions to insect stings. Cochrane Database Syst. Rev. 2012, 10, CD008838. [Google Scholar] [CrossRef]
- Ruëff, F.; Vos, B.; Oude Elberink, J.; Bender, A.; Chatelain, R.; Dugas-Breit, S.; Horny, H.-P.; Kuchenhoff, H.; Linhardt, A.; Mastnik, S.; et al. Predictors of clinical effectiveness of Hymenoptera venom immunotherapy. Clin. Exp. Allergy 2014, 44, 736–746. [Google Scholar] [CrossRef]
- Jarkvist, J.; Salehi, C.; Akin, C.; Gülen, T. Venom immunotherapy in patients with clonal mast cell disorders: IgG4 correlates with protection. Allergy Eur. J. Allergy Clin. Immunol. 2020, 75, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, M.; Nico, A.; Sinisi, A.; Giliberti, L.; Rossi, M.P.; Rossini, M.; Kourtis, G.; Rucco, A.S.; LoConte, F.; Muolo, L.; et al. A 13-year real-life study on efficacy, safety and biological effects of Vespula venom immunotherapy. Clin. Mol. Allergy 2018, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Arzt, L.; Bokanovic, D.; Schrautzer, C.; Laipold, K.; Möbs, C.; Pfützner, W.; Herzog, S.A.; Vollmann, J.; Reider, N.; Bohle, B.; et al. Immunological differences between insect venom-allergic patients with and without immunotherapy and asymptomatically sensitized subjects. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 1223–1231. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Number | Age (Years) | Insect That Causes an Allergic Reaction | Grade of the Allergic Reaction according to the Mueller Scale | Additional Diseases |
|---|---|---|---|---|
| Study Group | ||||
| UR-1 | 64 | Wasp | IV | Hypertension |
| UR-2 | 58 | Wasp | III | Allergic rhinitis, asthma |
| UR-3 | 38 | Wasp | III | Asthma |
| UR-4 | 28 | Wasp | III | Asthma |
| UR-5 | 31 | Wasp | III | - |
| UR-7 | 64 | Wasp | III | Asthma, hypertension |
| UR-8 | 54 | Wasp | III | Allergic rhinitis, asthma |
| UR-9 | 67 | Wasp | IV | Hypertension |
| UR-10 | 52 | Wasp | III | Tachycardia |
| UR-11 | 57 | Wasp | III | - |
| UR-19 | 48 | Wasp | III | - |
| UR-28 | 51 | Bee | III | - |
| UR-29 | 62 | Hornet | IV | - |
| UR-31 | 50 | Hornet | IV | Hypertension |
| UR-32 | 47 | Bee | III | - |
| UR-33 | 42 | Wasp | IV | - |
| UR-34 | 37 | Wasp | III | Asthma |
| UR-35 | 45 | Bee | III | - |
| UR-36 | 53 | Wasp | III | - |
| UR-37 | 40 | Bee | III | - |
| UR-38 | 46 | Wasp | III | - |
| UR-40 | 40 | Wasp | III | Allergic rhinitis, asthma |
| Control Group | ||||
| UR-13 | 29 | Wasp | III | Allergic rhinitis |
| UR-14 | 30 | Wasp | I | - |
| UR-20 | 48 | Wasp | III | - |
| UR-21 | 61 | Wasp | IV | Hypertension |
| UR-23 | 40 | Wasp | III | - |
| UR-24 | 51 | Wasp | III | Allergic rhinitis, hypertension, ischemic heart disease |
| UR-25 | 36 | Wasp | I | Allergic rhinitis, hypothyroidism |
| Precursor Ion m/z | Protein Name | 1st Day | 11th Day | 90th Day | |||
|---|---|---|---|---|---|---|---|
| p-Value | FDR | p-Value | FDR | p-Value | FDR | ||
| 1617.57 | Fibrinogen alpha chain | 0.00080919 | 0.029131 | >0.05 | >0.05 | 0.0010915 | 0.0056133 |
| 1449.73 | Complement C4-A | 0.0019183 | 0.03357 | >0.05 | >0.05 | >0.05 | >0.05 |
| 2554.56 | Fibrinogen alpha chain | 0.0027975 | 0.03357 | >0.05 | >0.05 | >0.05 | >0.05 |
| 3315.61 | x | 0.0042908 | 0.038618 | >0.05 | >0.05 | 0.018102 | 0.047737 |
| 1077.93 | Fibrinogen alpha chain | >0.05 | >0.05 | 0.0018253 | 0.044054 | 0.00084769 | 0.0050861 |
| 1020.92 | Fibrinogen alpha chain | >0.05 | >0.05 | 0.0031328 | 0.044054 | 0.00072854 | 0.0050861 |
| 1519.87 | Complement C3 | 0.030118 | >0.05 | 0.0036712 | 0.044054 | 0.00012064 | 0.0014477 |
| 1466.66 | Fibrinogen alpha chain | 0.04616 | >0.05 | >0.05 | >0.05 | 1.1921 × 10−5 | 0.00028324 |
| 1207.25 | Fibrinogen alpha chain | >0.05 | >0.05 | >0.05 | >0.05 | 1.5736 × 10−5 | 0.00028324 |
| 1351.37 | Fibrinogen alpha chain | 0.031164 | >0.05 | >0.05 | >0.05 | 0.0005094 | 0.0045846 |
| 2093.20 | Fibrinogen alpha chain | >0.05 | >0.05 | >0.05 | >0.05 | 0.0035243 | 0.015859 |
| 4053.21 | m/z > 3500 | >0.05 | >0.05 | >0.05 | >0.05 | 0.0042908 | 0.017163 |
| 1537.73 | Fibrinogen alpha chain | 0.011473 | >0.05 | >0.05 | >0.05 | 0.008316 | 0.029938 |
| 1262.25 | Fibrinogen alpha chain | 0.018957 | >0.05 | >0.05 | >0.05 | 0.014095 | 0.042284 |
| 2660.77 | Fibrinogen alpha chain | >0.05 | >0.05 | >0.05 | >0.05 | 0.014095 | 0.042284 |
| 1420.10 | Filamin-B | >0.05 | >0.05 | >0.05 | >0.05 | 0.018564 | 0.047737 |
| Precursor Ion m/z | Protein Name | 90th Day | 180th Day | ||
|---|---|---|---|---|---|
| p-Value | FDR | p-Value | FDR | ||
| 1945.24 | Kininogen-1 | 0.000254 | 0.009156 | 0.000618 | 0.01857 |
| 2093.20 | Fibrinogen alpha chain | 0.000584 | 0.009573 | 0.001032 | 0.01857 |
| 2660.77 | Fibrinogen alpha chain | 0.000798 | 0.009573 | 0.008021 | 0.035102 |
| 1546.56 | Fibrinogen alpha chain | 0.001546 | 0.012693 | 0.00605 | 0.035102 |
| 1331.52 | Myosin-9 | 0.001806 | 0.012693 | >0.05 | >0.05 |
| 3315.61 | x | 0.002116 | 0.012693 | 0.005189 | 0.035102 |
| 4053.21 | m/z > 3500 | 0.004214 | 0.018932 | 0.008776 | 0.035102 |
| 1519.87 | Complement C3 | 0.004545 | 0.018932 | 0.008021 | 0.035102 |
| 1262.25 | Fibrinogen alpha chain | 0.005259 | 0.018932 | 0.006973 | 0.035102 |
| 1537.73 | Fibrinogen alpha chain | 0.005259 | 0.018932 | 0.006973 | 0.035102 |
| 1207.25 | Fibrinogen alpha chain | 0.006998 | 0.022903 | 0.013758 | 0.045027 |
| 1466.66 | Fibrinogen alpha chain | 0.009238 | 0.027159 | 0.012058 | 0.04341 |
| 1866.3 | Complement C3 | 0.009807 | 0.027159 | 0.018957 | >0.05 |
| 1221.15 | Inter-alpha-trypsin inhibitor heavy chain H1 | 0.016742 | 0.043051 | 0.046026 | >0.05 |
| 1020.92 | Fibrinogen alpha chain | 0.019475 | 0.046741 | 0.022328 | >0.05 |
| 3240.91 | Fibrinogen alpha chain | 0.022615 | 0.048532 | 0.04588 | >0.05 |
| 1617.57 | Fibrinogen alpha chain | 0.022918 | 0.048532 | 0.046123 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matysiak, J.; Matuszewska, E.; Kowalski, M.L.; Kosiński, S.W.; Smorawska-Sabanty, E.; Matysiak, J. Association between Venom Immunotherapy and Changes in Serum Protein—Peptide Patterns. Vaccines 2021, 9, 249. https://doi.org/10.3390/vaccines9030249
Matysiak J, Matuszewska E, Kowalski ML, Kosiński SW, Smorawska-Sabanty E, Matysiak J. Association between Venom Immunotherapy and Changes in Serum Protein—Peptide Patterns. Vaccines. 2021; 9(3):249. https://doi.org/10.3390/vaccines9030249
Chicago/Turabian StyleMatysiak, Joanna, Eliza Matuszewska, Marek L. Kowalski, Sławomir W. Kosiński, Ewa Smorawska-Sabanty, and Jan Matysiak. 2021. "Association between Venom Immunotherapy and Changes in Serum Protein—Peptide Patterns" Vaccines 9, no. 3: 249. https://doi.org/10.3390/vaccines9030249
APA StyleMatysiak, J., Matuszewska, E., Kowalski, M. L., Kosiński, S. W., Smorawska-Sabanty, E., & Matysiak, J. (2021). Association between Venom Immunotherapy and Changes in Serum Protein—Peptide Patterns. Vaccines, 9(3), 249. https://doi.org/10.3390/vaccines9030249