
Ageing of Immune System and Response to a Live-Attenuated Herpes Zoster Vaccine in Lung Transplant Candidates

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Assessment of Humoral Immune Response to HZ Vaccination
2.3. Assessment of Cellular Immune Response to HZ Vaccination
2.4. Flow Cytometry
2.5. Statistical Analyses
3. Results
3.1. Patient Characteristics and Safety of HZ Vaccine
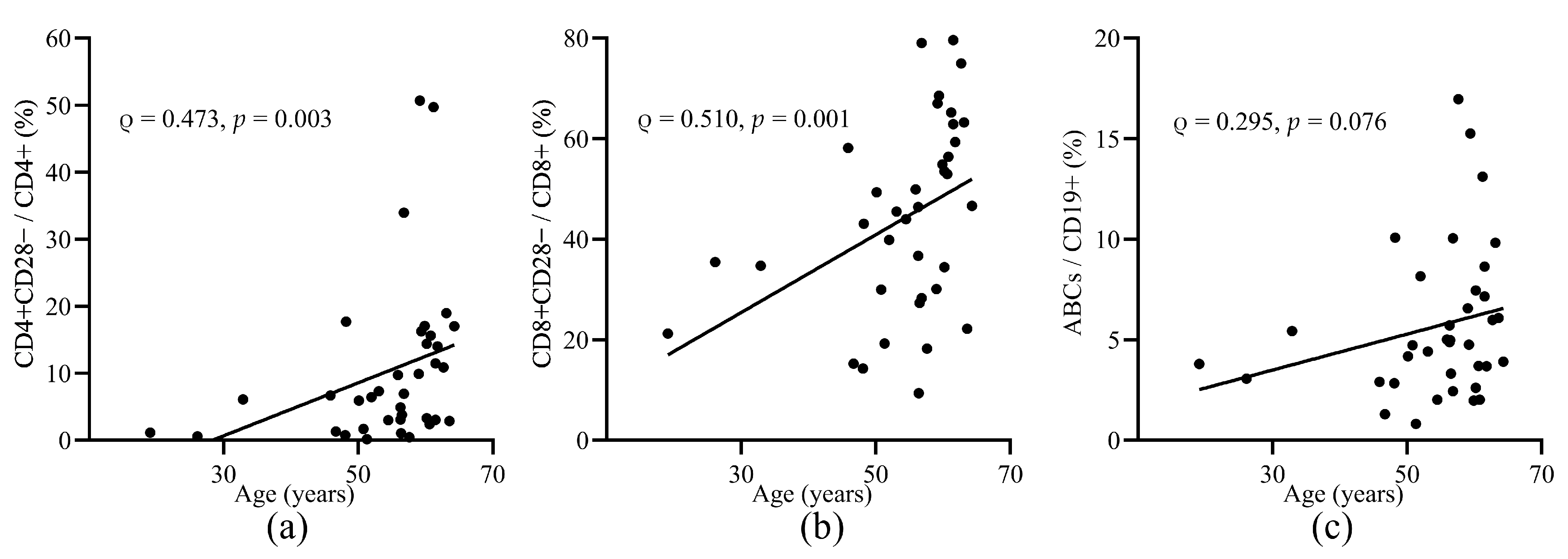
3.2. Aged T and B Cell Subsets before Vaccination
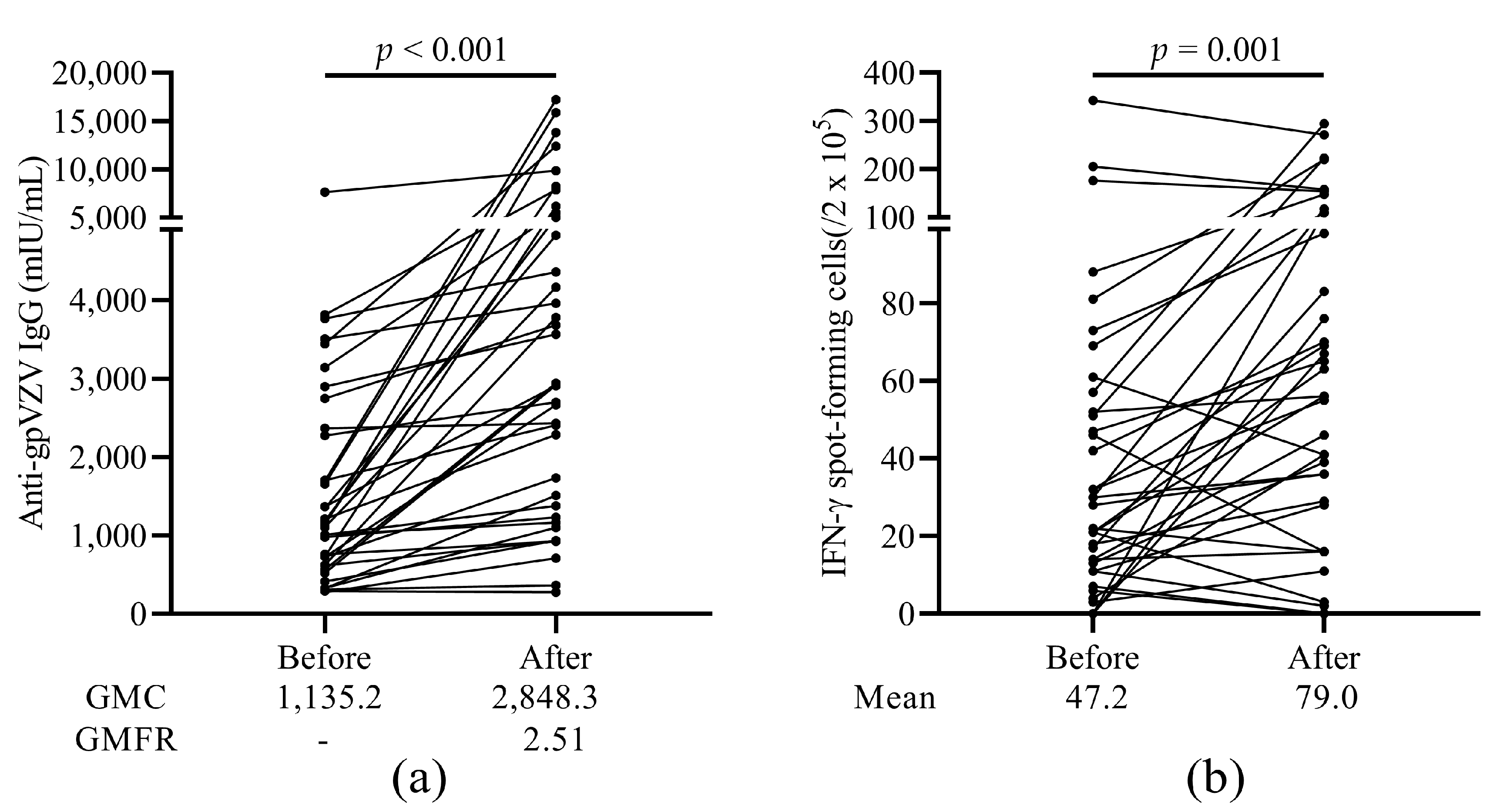
3.3. Humoral and Cellular Immune Response to HZ Vaccine
3.4. The Relationship between Response to HZ Vaccination and Baseline T and B Cell Subsets
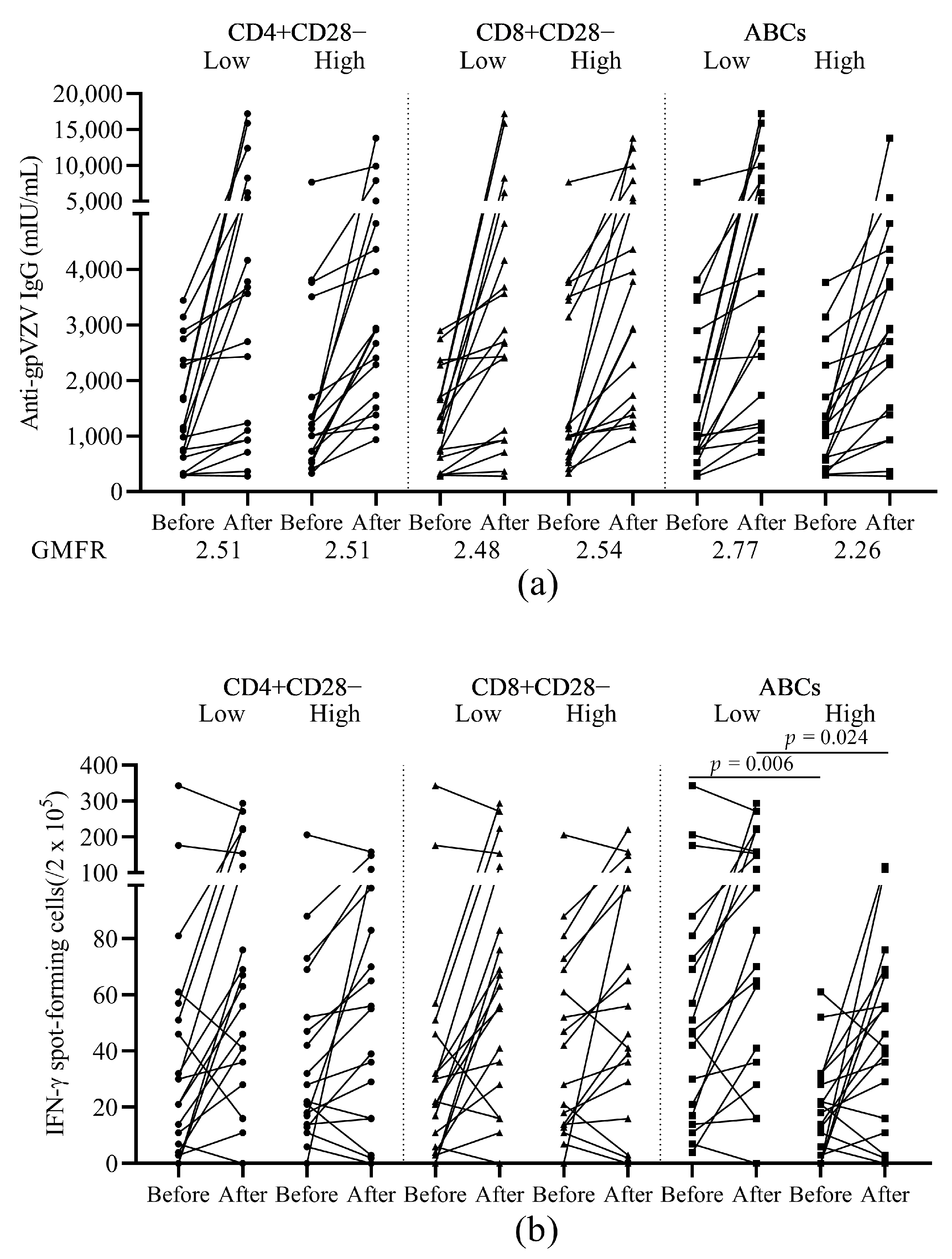
3.5. The Relationship between Response to HZ Vaccination and Baseline Ageing Cell Subsets
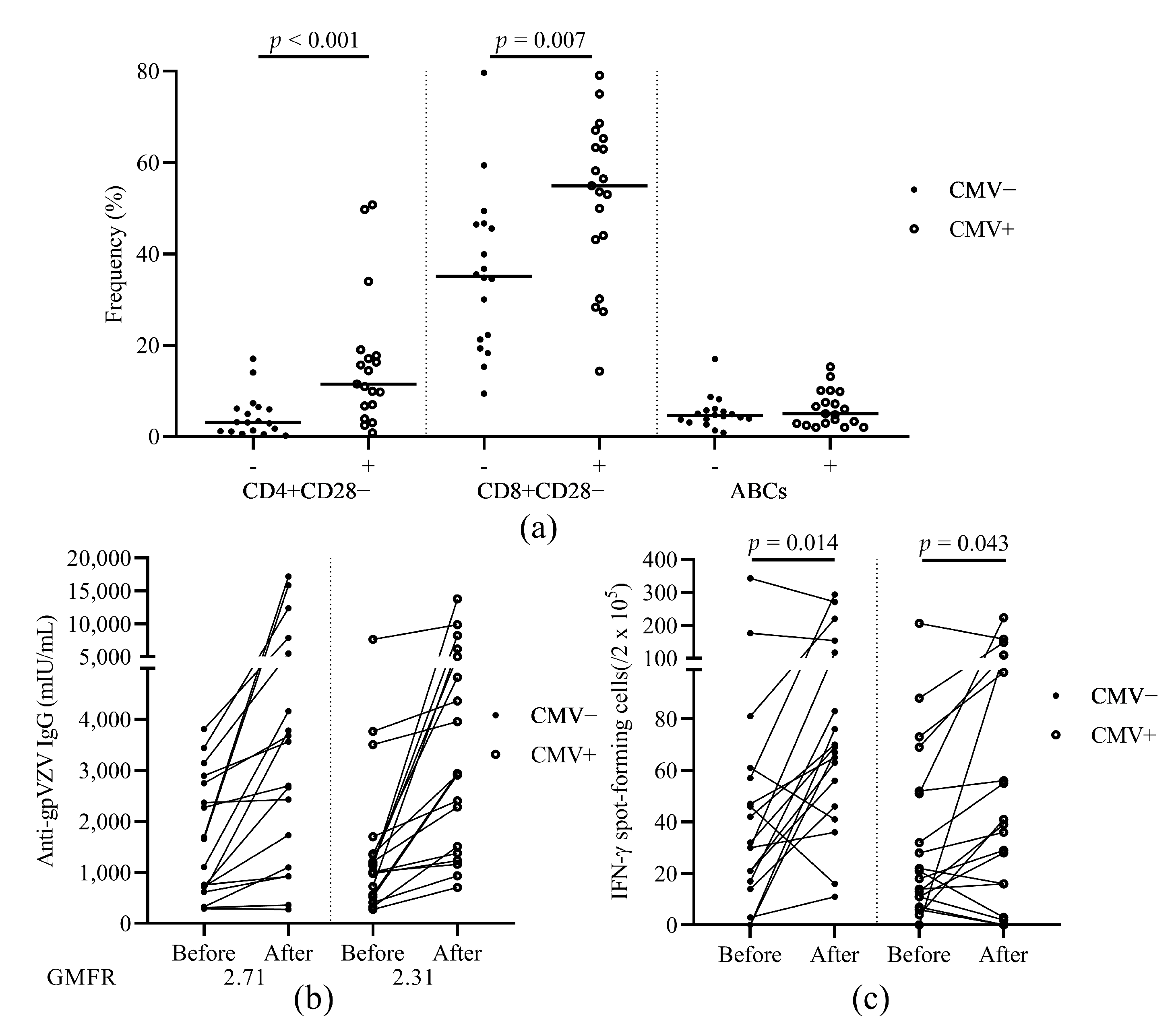
3.6. The Relationship of HZ Vaccination Response and CMV Latency
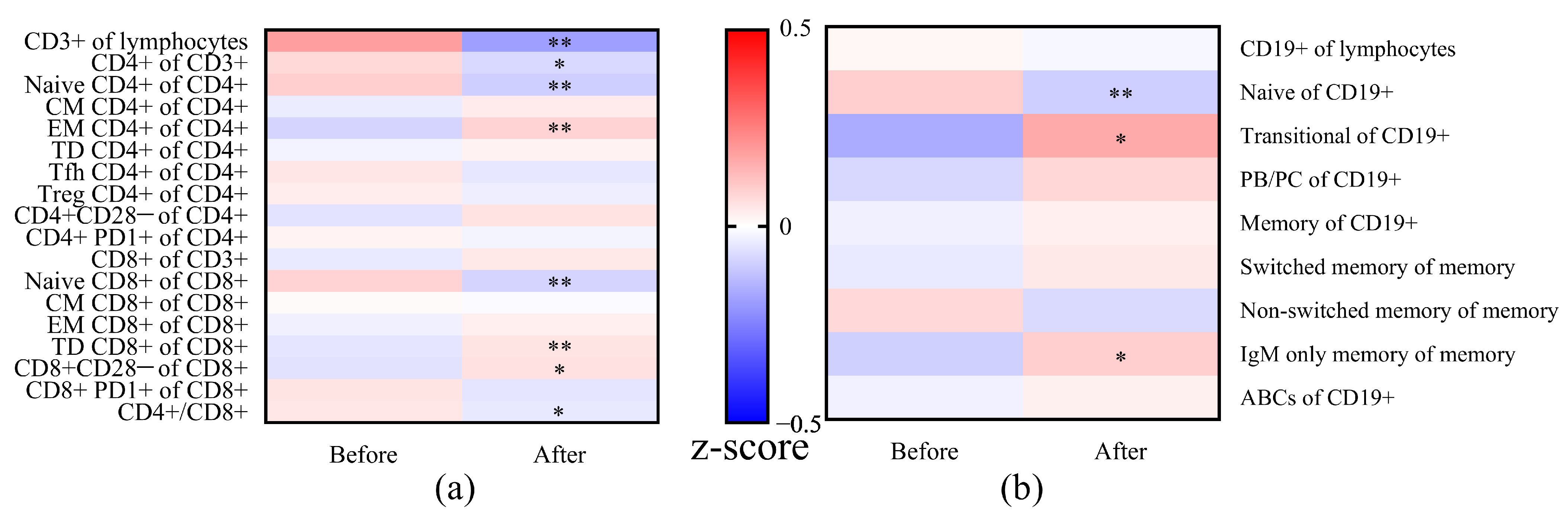
3.7. Changes in T and B Cell Subsets after Vaccination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subsets | Before | After | p-Value |
|---|---|---|---|
| CD3+/Lymphocytes (%) | 73.8 (66.8–81.2) | 70.7 (62.1–76.9) | <0.001 |
| CD4+/CD3+ (%) | 67.4 (58.3–75.8) | 65.8 (54.0–73.2) | 0.029 |
| Naïve CD4+/CD4+ (%) | 52.7 (28.7–61.3) | 45.6 (31.3–57.7) | <0.001 |
| CM CD4+/CD4+ (%) | 20.6 (13.1–28.3) | 20.4 (15.3–28.3) | ns |
| EM CD4+/CD4+ (%) | 21.3 (17.2–30.7) | 24.3 (20.4–33.6) | <0.001 |
| TD CD4+/CD4+ (%) | 3.8 (2.7–7.8) | 4.6 (2.0–8.0) | ns |
| Tfh CD4+/CD4+ (%) | 15.9 (11.4–22.1) | 16.2 (10.7–21.2) | ns |
| Treg CD4+/CD4+ (%) | 6.5 (5.2–7.8) | 6.5 (5.1–8.3) | ns |
| CD4+CD28−/CD4+ (%) | 6.5 (2.7–15.0) | 7.5 (2.5–16.5) | ns |
| CD4+PD1+/CD4+ (%) | 1.5 (1.0–2.1) | 1.8 (1.1–2.2) | ns |
| CD8+/CD3+ (%) | 24.7 (17.8–34.2) | 28.9 (20.4–35.9) | ns |
| Naïve CD8+/CD8+ (%) | 25.4 (11.8–40.4) | 20.4 (9.5–36.2) | 0.002 |
| CM CD8+/CD8+ (%) | 2.8 (1.5–5.8) | 2.8 (1.6–5.4) | ns |
| EM CD8+/CD8+ (%) | 28.2 (15.4–37.2) | 27.3 (17.3–40.3) | ns |
| TD CD8+/CD8+ (%) | 38.3 (21.4–59.0) | 38.8 (23.8–60.0) | 0.005 |
| CD8+CD28−/CD8+ (%) | 45.5 (29.2–58.8) | 52.9 (25.0–62.1) | 0.012 |
| CD8+PD1+/CD8+ (%) | 1.0 (0.6–2.0) | 0.9 (0.6–1.6) | ns |
| CD4+/CD8+ Ratio | 2.7 (1.7–4.2) | 2.3 (1.5–3.6) | 0.046 |
| CD19+/Lymphocytes (%) | 3.8 (2.3–5.5) | 3.8 (2.4–4.9) | ns |
| Naive CD19+/CD19+ (%) | 70.6 (59.7–76.2) | 67.4 (60.2–70.0) | 0.007 |
| Transitional CD19+/CD19+ (%) | 2.7 (1.4–6.2) | 4.9 (1.8–7.4) | 0.032 |
| PB/PC CD19+/CD19+ (%) | 1.3 (1.0–2.3) | 1.7 (1.1–2.8) | ns |
| Memory CD19+/CD19+ (%) | 22.7 (15.7–32.4) | 24.4 (17.3–33.2) | ns |
| Switched Memory CD19+/Memory CD19+ (%) | 18.4 (11.2–24.1) | 17.8 (12.4–25.0) | ns |
| Non-Switched Memory CD19+/Memory CD19+ (%) | 75.5 (65.2–81.1) | 73.2 (66.8–81.4) | ns |
| IgM Only Memory CD19+/Memory CD19+ (%) | 0.5 (0.3–0.9) | 0.6 (0.4–1.0) | 0.029 |
| ABCs/CD19+ (%) | 4.9 (3.0–7.3) | 4.4 (2.9–7.8) | ns |
| Frequencies before Vaccination | VZV-IgG a | VZV-SFCs a | ||||
|---|---|---|---|---|---|---|
| Low (N = 19) | High (N = 18) | p-Value | Low (N = 19) | High (N = 18) | p-Value | |
| CD3+/Lymphocytes (%) | 75.6 (66.5–80.5) | 73.3 (67.3–82.6) | ns | 76.8 (66.7–82.0) | 72.5 (66.8–76.7) | ns |
| CD4+/CD3+ (%) | 71.9 (64.8–78.1) | 61.7 (54.0–67.5) | 0.011 | 67.9 (53.1–78.1) | 66.9 (60.0–74.2) | ns |
| Naïve CD4+/CD4+ (%) | 56.8 (24.2–63.3) | 47.4 (34.7–56.1) | ns | 47.4 (27.0–61.6) | 55.4 (34.5–61.2) | ns |
| CM CD4+/CD4+ (%) | 17.8 (12.7–26.2) | 22.8 (13.8–28.8) | ns | 22.2 (12.7–28.9) | 18.8 (13.0–26.8) | ns |
| EM CD4+/CD4+ (%) | 18.7 (14.1–37.0) | 23.0 (19.8–29.8) | ns | 21.3 (17.9–37.0) | 19.7 (14.0–25.6) | ns |
| TD CD4+/CD4+ (%) | 4.2 (2.7–6.6) | 3.5 (2.4–9.3) | ns | 4.2 (2.8–8.9) | 3.3 (1.6–7.6) | ns |
| Tfh CD4+/CD4+ (%) | 15.3 (10.9–21.7) | 16.8 (12.1–23.0) | ns | 17.4 (10.9–21.9) | 14.8 (11.7–22.4) | ns |
| Treg CD4+/CD4+ (%) | 5.9 (4.9–7.7) | 6.8 (5.5–8.0) | ns | 6.3 (4.8–7.8) | 6.6 (5.5–8.0) | ns |
| CD4+CD28−/CD4+ (%) | 3.9 (1.7–16.3) | 6.8 (3.1–12.5) | ns | 9.9 (2.4–17.0) | 6.1 (2.6–10.8) | ns |
| CD4+PD1+/CD4+ (%) | 1.4 (0.9–1.9) | 1.6 (1.2–2.8) | ns | 1.3 (0.9–2.0) | 1.6 (1.2–2.3) | ns |
| CD8+/CD3+ (%) | 20.6 (16.3–27.9) | 30.6 (23.5–35.2) | 0.022 | 21.4 (16.4–35.8) | 25.7 (21.0–33.2) | ns |
| Naïve CD8+/CD8+ (%) | 22.5 (6.6–31.6) | 27.2 (14.1–46.5) | ns | 19.3 (9.0–41.7) | 27.8 (15.4–415) | ns |
| CM CD8+/CD8+ (%) | 3.9 (1.8–6.4) | 2.6 (0.9–4.5) | ns | 2.8 (1.7–5.9) | 2.5 (0.9–5.0) | ns |
| EM CD8+/CD8+ (%) | 34.0 (24.3–47.0) | 23.1 (13.2–30.5) | 0.024 | 28.3 (13.4–38.5) | 27.2 (18.8–37.7) | ns |
| TD CD8+/CD8+ (%) | 38.3 (21.6–50.6) | 38.7 (20.7–59.4) | ns | 38.5 (24.1–61.6) | 36.3 (19.5–58.9) | ns |
| CD8+CD28−/CD8+ (%) | 36.8 (22.2–59.4) | 46.6 (34.1–59.4) | ns | 46.7 (28.3–65.2) | 44.8 (28.1–56.9) | ns |
| CD8+PD1+/CD8+ (%) | 1.0 (0.6–2.4) | 1.0 (0.5–1.7) | ns | 0.8 (0.5–1.5) | 1.1 (0.7–2.4) | ns |
| CD4+/CD8+ Ratio | 3.7 (2.3–4.7) | 2.0 (1.6–2.9) | 0.019 | 3.3 (1.5–4.5) | 2.6 (1.9–3.6) | ns |
| CD19+/Lymphocytes (%) | 3.7 (2.2–5.8) | 4.1 (2.3–5.6) | ns | 4.5 (2.0–5.8) | 3.8 (2.3–5.6) | ns |
| Naive CD19+/CD19+ (%) | 66.5 (59.9–81.2) | 72.6 (58.4–75.5) | ns | 70.6 (55.4–81.2) | 71.1 (59.8–75.5) | ns |
| Transitional CD19+/CD19+ (%) | 2.1 (1.6–5.2) | 3.6 (1.2–7.0) | ns | 1.9 (0.6–3.6) | 4.4 (2.1–7.0) | 0.030 |
| PB/PC CD19+/CD19+ (%) | 1.2 (0.7–2.8) | 1.6 (1.3–2.2) | ns | 1.2 (0.9–2.8) | 1.4 (1.2–2.2) | ns |
| Memory CD19+/CD19+ (%) | 25.1 (15.8–32.8) | 21.6 (15.4–31.8) | ns | 25.0 (15.8–36.8) | 21.6 (15.4–30.2) | ns |
| Switched Memory CD19+/Memory CD19+ (%) | 20.3 (11.9–25.2) | 17.6 (9.5–20.6) | ns | 19.5 (11.9–32.0) | 16.7 (9.5–20.7) | ns |
| Non-Switched Memory CD19+/Memory CD19+ (%) | 75.5 (63.8–81.0) | 76.0 (69.8–81.6) | ns | 72.4 (62.4–80.6) | 77.4 (68.3–82.5) | ns |
| IgM Only Memory CD19+/Memory CD19+ (%) | 0.4 (0.3–0.6) | 0.8 (0.4–1.3) | ns | 0.5 (0.3–1.3) | 0.5 (0.3–0.9) | ns |
| ABCs/CD19+ (%) | 4.9 (3.3–7.5) | 4.3 (2.9–7.4) | ns | 5.0 (3.1–8.6) | 4.6 (2.9-5.8) | ns |
References
- Young, K.A.; Dilling, D.F. The future of lung transplantation. Chest 2019, 155, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Hachem, R.R. advancing lung transplantation. Clin. Transpl. 2015, 31, 239–247. [Google Scholar]
- Yusen, R.D.; Christie, J.D.; Edwards, L.B.; Kucheryavaya, A.Y.; Benden, C.; Dipchand, A.I.; Dobbels, F.; Kirk, R.; Lund, L.H.; Stehlik, J.; et al. The registry of the International Society for Heart and Lung Transplantation: Thirtieth Adult Lung and Heart-Lung Transplant Report—2013; focus theme: Age. J. Heart Lung Transplant. 2013, 32, 965–978. [Google Scholar] [CrossRef] [PubMed]
- Hartert, M.; Senbaklavacin, O.; Gohrbandt, B.; Fischer, B.M.; Buhl, R.; Vahld, C.-F. Lung transplantation: A treatment option in end-stage lung disease. Dtsches Arzteblatt Int. 2014, 111, 107–116. [Google Scholar] [CrossRef]
- Colvin, M.M.; Smith, C.A.; Tullius, S.G.; Goldstein, D.R. Aging and the immune response to organ transplantation. J. Clin. Invest. 2017, 127, 2523–2529. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.M. The effect of ageing of the immune system on vaccination responses. Hum. Vaccines Immunother. 2013, 9, 1364–1367. [Google Scholar] [CrossRef]
- Cancro, M.P. Age-associated B cells. Annu. Rev. Immunol. 2020, 38, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Zerboni, L.; Sen, N.; Oliver, S.L.; Arvin, A.M. Molecular mechanisms of varicella zoster virus pathogenesis. Nat. Rev. Microbiol. 2014, 12, 197–214. [Google Scholar] [CrossRef]
- Oxman, M.N.; Levin, M.J.; Johnson, G.R.; Schmader, K.E.; Straus, S.E.; Gelb, L.D.; Arbeit, R.D.; Simberkoff, M.S.; Gershon, A.A.; Davis, L.E.; et al. A vaccine to prevent herpes zoster and postherpetic neuralgia in older adults. N. Engl. J. Med. 2005, 352, 2271–2284. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.J.; Weinberg, A. Immune responses to zoster vaccines. Hum. Vaccines Immunother. 2019, 15, 772–777. [Google Scholar] [CrossRef]
- Wang, L.; Verschuuren, E.A.M.; van Leer-Buter, C.C.; Bakker, S.J.L.; de Joode, A.A.E.; Westra, J.; Bos, N.A. Herpes zoster and immunogenicity and safety of zoster vaccines in transplant patients: A narrative review of the literature. Front. Immunol. 2018, 9, 1632. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence and human vaccine immune responses. Immun. Ageing 2019, 16, 25. [Google Scholar] [CrossRef]
- Weinberg, A.; Canniff, J.; Rouphael, N.; Mehta, A.; Mulligan, M.; Whitaker, J.A.; Levin, M.J. Varicella-zoster virus-specific cellular immune responses to the live attenuated zoster vaccine in young and older adults. J. Immunol. 2017, 199, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Verschuuren, E.A.M.; Paap, D.; Rondaan, C.; Raveling-Eelsing, E.; Westra, J.; Bos, N.A. Prophylactic vaccination with a live-attenuated herpes zoster vaccine in lung transplant candidates. J. Heart Lung Transplant. 2020, 12, 1445–1454. [Google Scholar] [CrossRef]
- Rondaan, C.; de Haan, A.; Horst, G.; Hempel, J.C.; van Leer, C.; Bos, N.A.; van Assen, S.; Bijl, M.; Westra, J. Altered cellular and humoral immunity to varicella-zoster virus in patients with autoimmune diseases. Arthritis Rheumatol. 2014, 66, 3122–3128. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, A.; Pang, L.; Johnson, M.J.; Caldas, Y.; Cho, A.; Tovar-Salazar, A.; Cannif, J.; Schmader, K.E.; Popmihajlov, Z.; Levin, M.J. The effect of age on the immunogenicity of the live attenuated zoster vaccine is predicted by baseline regulatory T Cells and varicella-zoster virus-specific t cell immunity. J. Virol. 2019, 93, e00305-19. [Google Scholar] [CrossRef]
- Vermeulen, J.N.; Lange, J.M.A.; Tyring, S.K.; Peters, P.H.; Nunez, M.; Poland, G.; Levin, M.J.; Freeman, C.; Chalikondam, I.; Smith, J.G.; et al. Safety tolerability, and immunogenicity after 1 and 2 doses of zoster vaccine in healthy adults ≥60 years of age. Vaccine 2012, 30, 904–910. [Google Scholar] [CrossRef]
- Levin, M.J.; Schmader, K.E.; Pang, L.; Williams-Diaz, A.; Zerbe, G.; Canniff, J.; Johnson, M.J.; Caldas, Y.; Cho, A.; Lang, N.; et al. Cellular and humoral responses to a second dose of herpes zoster vaccine administered 10 years after the first dose among older adults. J. Infect. Dis. 2016, 213, 14–22. [Google Scholar] [CrossRef]
- Li, S.; Sullivan, N.L.; Rouphael, N.; Yu, T.; Banton, S.; Maddur, M.S.; McCausland, M.; Chiu, C.; Canniff, J.; Dubey, S.; et al. Metabolic phenotypes of response to vaccination in humans. Cell 2017, 169, 862–877. [Google Scholar] [CrossRef]
- Van der Heiden, M.; de Rond, L.G.H.; van Zelm, M.C.; Berbers, G.A.M.; Boots, A.M.H.; Buisman, A.-M. Age-dependent pre-vaccination immunity affects the immunogenicity of varicella zoster vaccination in middle-aged adults. Front. Immunol. 2018, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Schmader, K.E.; Levin, M.J.; Gnann, J.W.; McNeil, S.A.; Vesikari, T.; Betts, R.F.; Keay, S.; Syek, J.E.; Bundick, N.D.; Su, S.-C.; et al. Efficacy, safety, and tolerability of herpes zoster vaccine in persons aged 50–59 years. Clin. Infect. Dis. 2012, 54, 922–928. [Google Scholar] [CrossRef]
- Levin, M.J.; Buchwald, U.K.; Gardner, J.; Martin, J.; Stek, J.E.; Brown, E.; Popmihajlov, Z. Immunogenicity and safety of zoster vaccine live administered with quadrivalent influenza virus vaccine. Vaccine 2018, 36, 179–185. [Google Scholar] [CrossRef]
- Weinberger, B.; Welzl, K.; Herndler-Brandstetter, D.; Parson, W.; Grubeck-Loebenstein, B. CD28(-)CD8(+) T cells do not contain unique clonotypes and are therefore dispensable. Immunol. Lett. 2009, 127, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Dugan, H.L.; Henry, C.; Wilson, P.C. Aging and influenza vaccine-induced immunity. Cell. Immunol. 2020, 348, 103998. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; O’Neill, P.; Naradikian, M.S.; Soholz, J.L.; Canoro, M.P. A B-cell subset uniquely responsive to innate stimuli accumulates in aged mice. Blood 2011, 118, 1294–1304. [Google Scholar] [CrossRef] [PubMed]
- Rubtsov, A.V.; Rubtsova, K.; Fischer, A.; Meehan, R.T.; Gillis, J.Z.; Kappler, J.W.; Marraok, P. Toll-like receptor 7 (TLR7)–driven accumulation of a novel CD11c+ B-cell population is important for the development of autoimmunity. Blood 2011, 118, 1305–1315. [Google Scholar] [CrossRef]
- Nipper, A.J.; Smithey, M.J.; Shah, R.C.; Canaday, D.H.; Landay, A.L. Diminished antibody response to influenza vaccination is characterized by expansion of an age-associated B-cell population with low PAX5. Clin. Immunol. 2018, 193, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Frasca, D.; Blomberg, B.B. Aging, cytomegalovirus (CMV) and influenza vaccine responses. Hum. Vaccines Immunother. 2015, 12, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Derhovanessian, E.; Theeten, H.; Hähnel, K.; Van Damme, P.; Cools, N.; Pawelec, G. Cytomegalovirus-associated accumulation of late-differentiated CD4 T-cells correlates with poor humoral response to influenza vaccination. Vaccine 2013, 31, 685–690. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Landin, A.M.; Blomberg, B.B. Cytomegalovirus (CMV) seropositivity decreases B cell responses to the influenza vaccine. Vaccine 2015, 33, 1433–1439. [Google Scholar] [CrossRef]
- Lal, H.; Cunningham, A.L.; Godeaux, O.; Chlibek, R.; Diez-Domingo, J.; Hwang, S.-J.; Levin, M.J.; McElhaney, J.E.; Poder, A.; Puig-Barberà, J.; et al. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults. N. Engl. J. Med. 2015, 372, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Racine, É.; Gilca, V.; Amini, R.; Tunis, M.; Ismail, S.; Sauvageau, C. A systematic literature review of the recombinant subunit herpes zoster vaccine use in immunocompromised 18–49 year old patients. Vaccine 2020, 38, 6205–6214. [Google Scholar] [CrossRef] [PubMed]





| Characteristics | N = 37 |
|---|---|
| Age at Vaccination in Years, Median (Range) | 56.8 (19.1–64.3) |
| Months After Vaccination, Median (Range) | 2.2 (0.7–5.8) |
| Gender, N (%) | |
| Male | 20 (54.1) |
| Female | 17 (45.9) |
| End Stage Pulmonary Disease, N (%) | |
| Chronic Obstructive Pulmonary Disease/Emphysema | 19 (51.4) |
| α1-Antitrypsin Deficiency | 9 (24.3) |
| Pulmonary Fibrosis/Interstitial Lung Disease | 3 (8.1) |
| Pulmonary Arterial Hypertension | 2 (5.4) |
| Cystic Fibrosis/Bronchiectasis | 4 (10.8) |
| Baseline Cytomegalovirus Serostatus, N (%) | |
| + | 19 (51.4) |
| – | 18 (48.6) |
| Age at Vaccination, N (%) | |
| ≤50 Years Old | 7 (18.9) |
| 50–60 Years Old | 18 (48.6) |
| >60 Years Old | 12 (32.4) |
| Follow Up Time After Vaccination in Months, Median (Range) | 23.7 (9.3–36.8) |
| Lung Transplantation During Follow Up, N (%) | 16 (43.2) 1 |
| Months After Vaccination at Transplantation, Median (Range) | 13.2 (2.8–29.6) |
| Subsets | Before | After | p-Value |
|---|---|---|---|
| CD3+/Lymphocytes (%) | 73.8 (66.8–81.2) | 70.7 (62.1–76.9) | <0.001 |
| CD4+/CD3+ (%) | 67.4 (58.3–75.8) | 65.8 (54.0–73.2) | 0.029 |
| Naïve CD4+/CD4+ (%) | 52.7 (28.7–61.3) | 45.6 (31.3–57.7) | <0.001 |
| EM CD4+/CD4+ (%) | 21.3 (17.2–30.7) | 24.3 (20.4–33.6) | <0.001 |
| Naïve CD8+/CD8+ (%) | 25.4 (11.8–40.4) | 20.4 (9.5–36.2) | 0.002 |
| TD CD8+/CD8+ (%) | 38.3 (21.4–59.0) | 38.8 (23.8–60.0) | 0.005 |
| CD8+CD28−/CD8+ (%) | 45.5 (29.2–58.8) | 52.9 (25.0–62.1) | 0.012 |
| CD4+/CD8+ Ratio | 2.7 (1.7–4.2) | 2.3 (1.5–3.6) | 0.046 |
| Naive CD19+/CD19+ (%) | 70.6 (59.7–76.2) | 67.4 (60.2–70.0) | 0.007 |
| Transitional CD19+/CD19+ (%) | 2.7 (1.4–6.2) | 4.9 (1.8–7.4) | 0.032 |
| IgM Only Memory CD19+/Memory CD19+ (%) | 0.5 (0.3–0.9) | 0.6 (0.4–1.0) | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Verschuuren, E.A.M.; Paap, D.; Rondaan, C.; Raveling-Eelsing, E.; Liu, S.; Westra, J.; Bos, N.A. Ageing of Immune System and Response to a Live-Attenuated Herpes Zoster Vaccine in Lung Transplant Candidates. Vaccines 2021, 9, 202. https://doi.org/10.3390/vaccines9030202
Wang L, Verschuuren EAM, Paap D, Rondaan C, Raveling-Eelsing E, Liu S, Westra J, Bos NA. Ageing of Immune System and Response to a Live-Attenuated Herpes Zoster Vaccine in Lung Transplant Candidates. Vaccines. 2021; 9(3):202. https://doi.org/10.3390/vaccines9030202
Chicago/Turabian StyleWang, Lei, Erik A.M. Verschuuren, Davy Paap, Christien Rondaan, Elisabeth Raveling-Eelsing, Siqi Liu, Johanna Westra, and Nicolaas A. Bos. 2021. "Ageing of Immune System and Response to a Live-Attenuated Herpes Zoster Vaccine in Lung Transplant Candidates" Vaccines 9, no. 3: 202. https://doi.org/10.3390/vaccines9030202
APA StyleWang, L., Verschuuren, E. A. M., Paap, D., Rondaan, C., Raveling-Eelsing, E., Liu, S., Westra, J., & Bos, N. A. (2021). Ageing of Immune System and Response to a Live-Attenuated Herpes Zoster Vaccine in Lung Transplant Candidates. Vaccines, 9(3), 202. https://doi.org/10.3390/vaccines9030202