The Roles of Osteopontin in the Pathogenesis of West Nile Encephalitis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Osteopontin
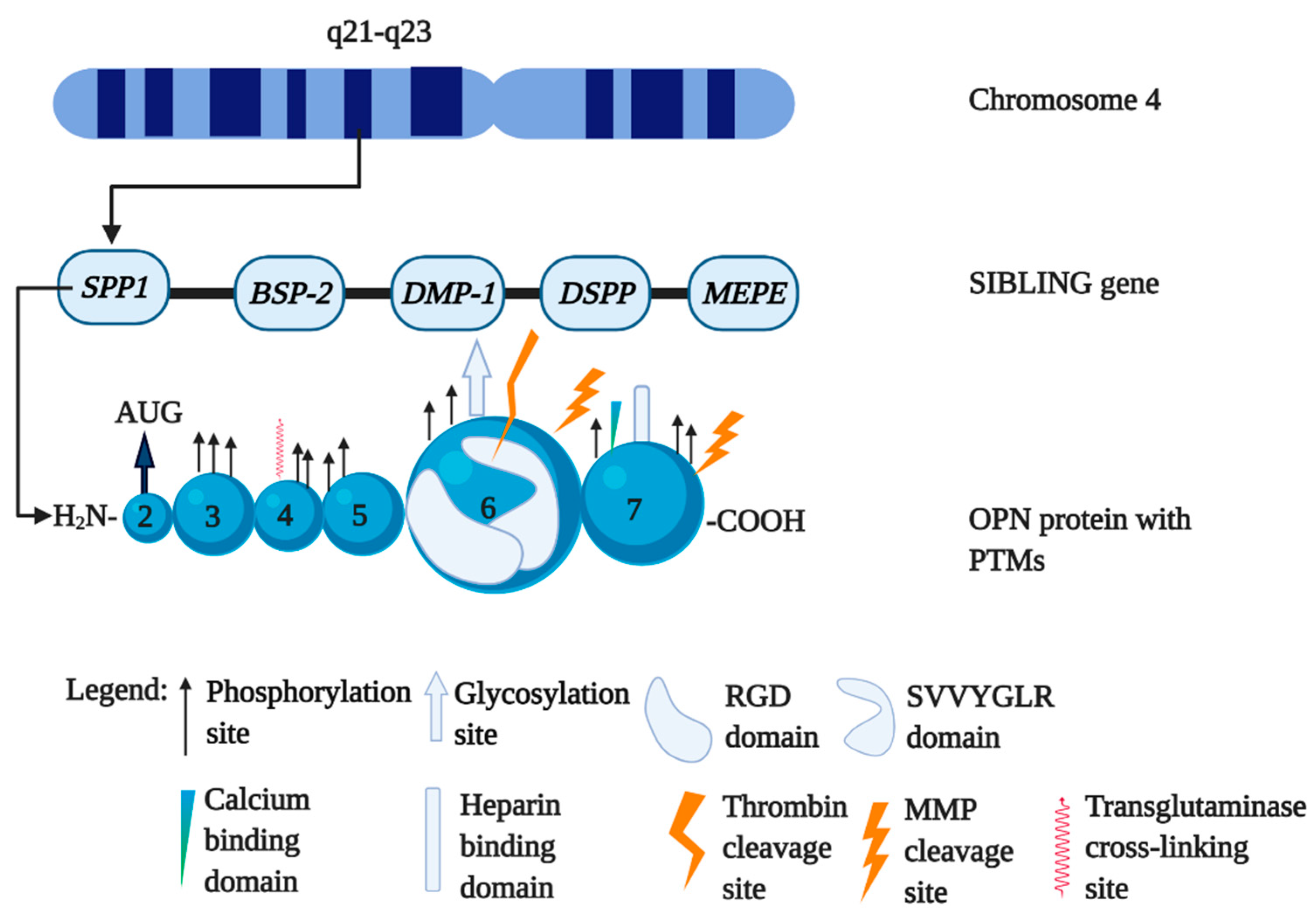
1.1. The Biology of Osteopontin
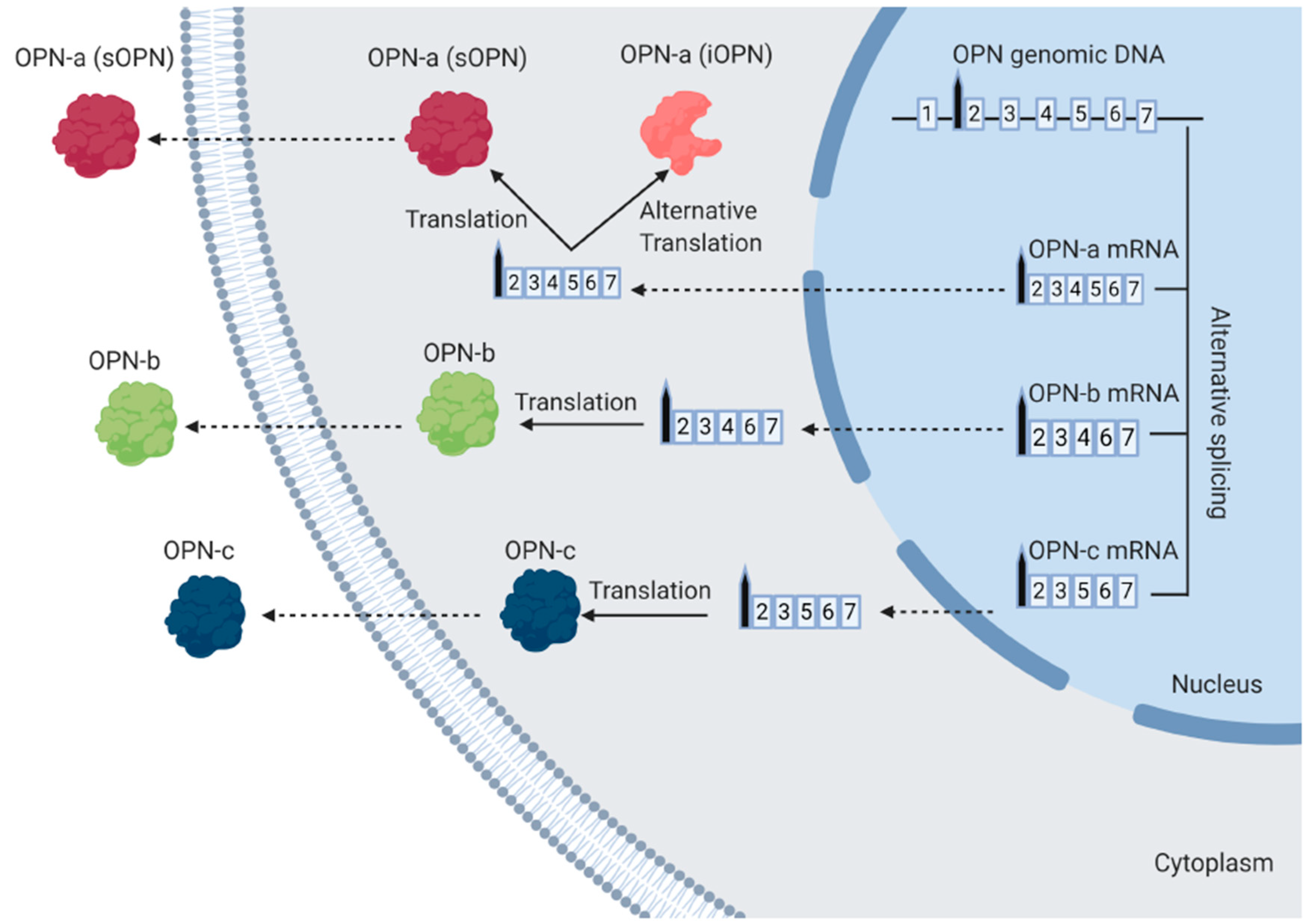
1.2. The Different Isoforms of OPN
1.3. The Function of Special Forms of OPN
1.3.1. Polymerized OPN
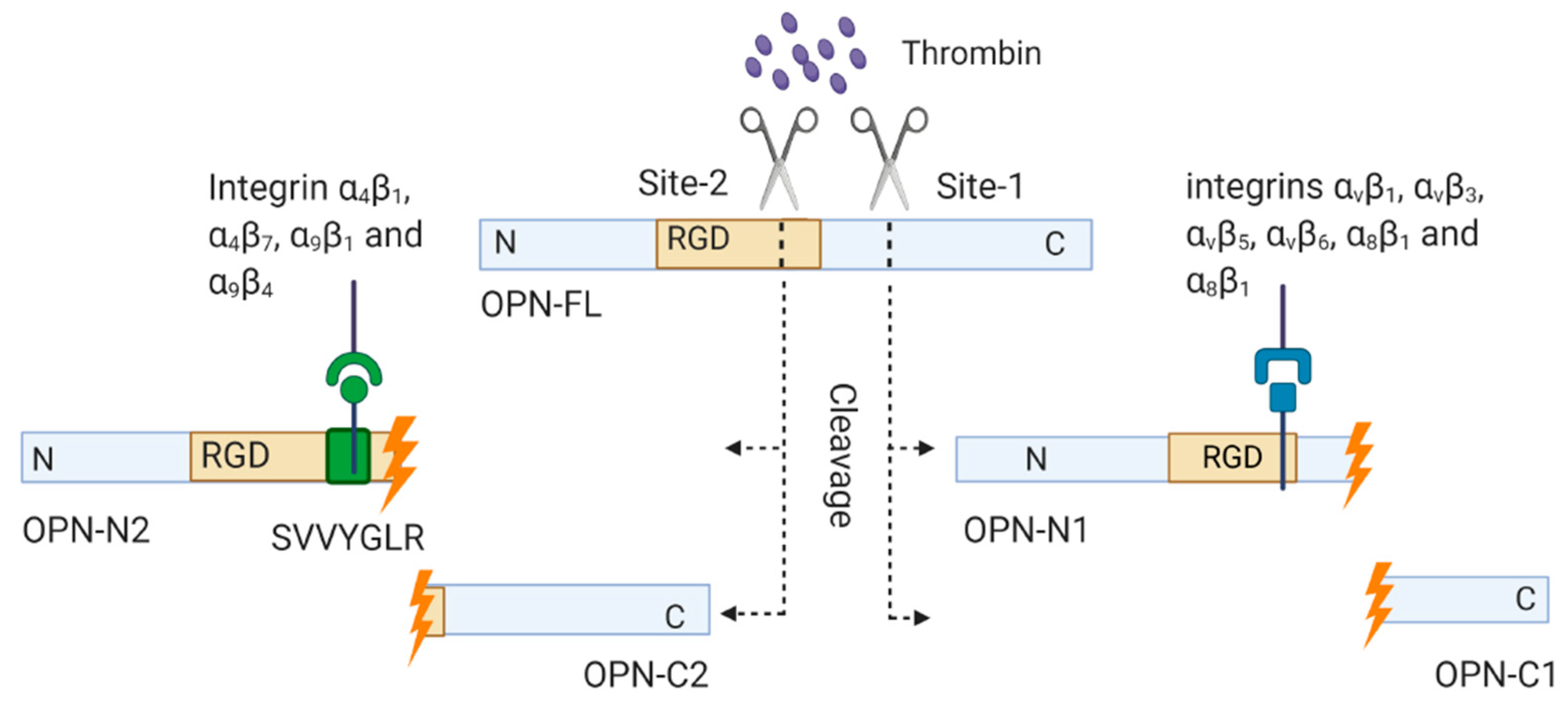
1.3.2. Thrombin-Cleaved OPN
2. Roles of OPN in the Pathogenesis of WNV
2.1. West Nile Virus
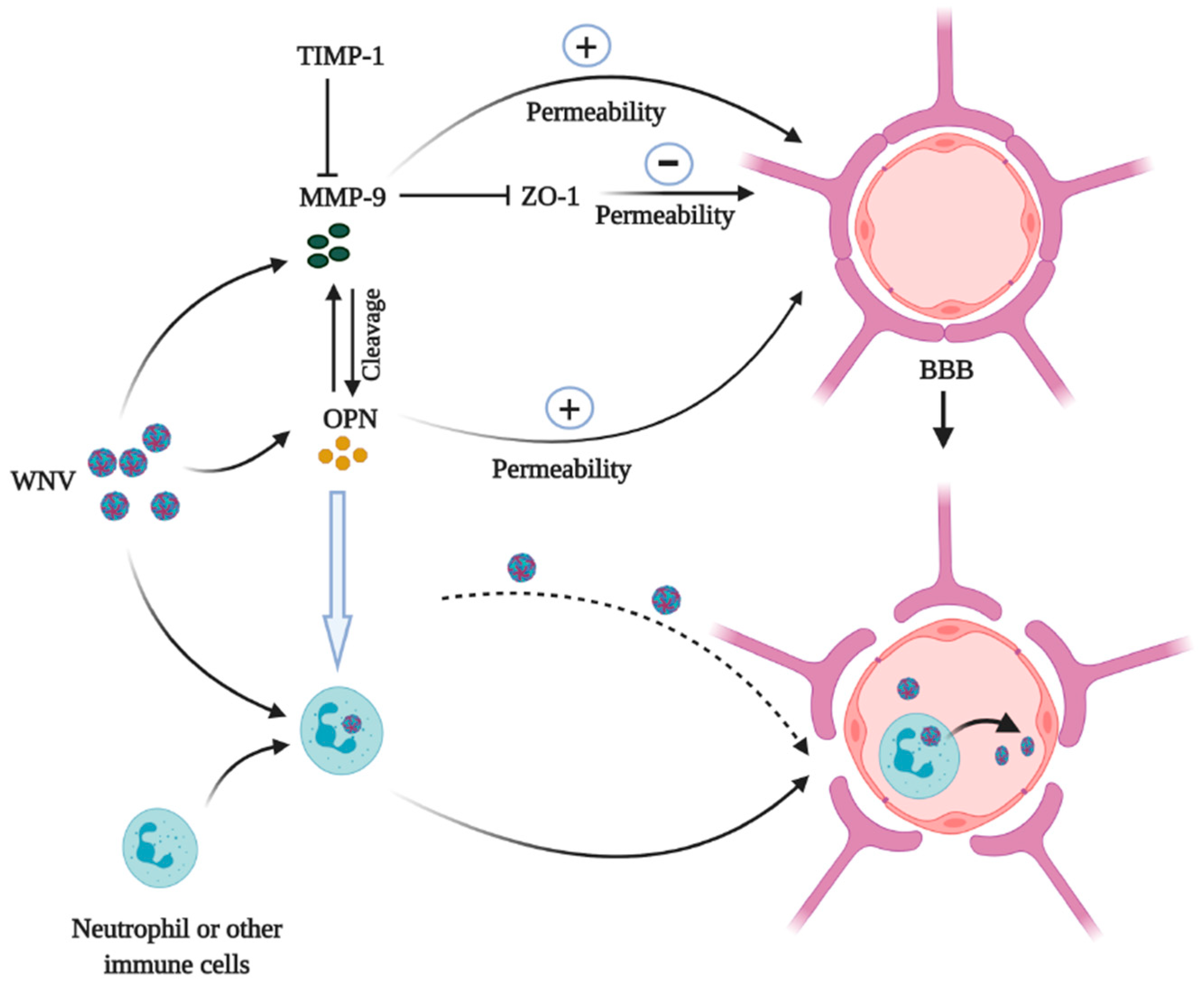
2.2. OPN and WNV
3. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Hao, C.; Cui, Y.; Owen, S.; Li, W.; Cheng, S.; Jiang, W.G. Human osteopontin: Potential clinical applications in cancer (Review). Int. J. Mol. Med. 2017, 39, 1327–1337. [Google Scholar] [CrossRef]
- McKee, M.D.; Cole, W.G. Bone matrix and mineralization. In Pediatric Bone; Academic Press: San Diego, CA, USA, 2012; pp. 9–37. [Google Scholar]
- Qin, C.; Baba, O.; Butler, W.T. Post-translational modifications of sibling proteins and their roles in osteogenesis and dentinogenesis. Crit. Rev. Oral Biol. Med. 2004, 15, 126–136. [Google Scholar] [CrossRef]
- Chaplet, M.; De Leval, L.; Waltregny, D.; Detry, C.; Fornaciari, G.; Bevilacqua, G.; Fisher, L.W.; Castronovo, V.; Bellahcène, A. Dentin matrix protein 1 is expressed in human lung cancer. J. Bone Miner. Res. 2003, 18, 1506–1512. [Google Scholar] [CrossRef]
- Castello, L.M.; Raineri, D.; Salmi, L.; Clemente, N.; Vaschetto, R.; Quaglia, M.; Garzaro, M.; Gentilli, S.; Navalesi, P.; Cantaluppi, V.; et al. Osteopontin at the Crossroads of Inflammation and Tumor Progression. Mediat. Inflamm. 2017, 2017, 4049098. [Google Scholar] [CrossRef]
- Kaleta, B. Role of osteopontin in systemic lupus erythematosus. Arch. Immunol. Exp. 2014, 62, 475–482. [Google Scholar] [CrossRef]
- Denhardt, T.D.; Noda, M. Osteopontin expression and function: Role in bone remodeling. J. Cell. Biochem. 1998, 72, 92–102. [Google Scholar] [CrossRef]
- Ramaiah, K.S.; Rittling, S. Role of osteopontin in regulating hepatic inflammatory responses and toxic liver injury. Expert Opin. Drug Metab. Toxicol. 2007, 3, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Uede, T. Osteopontin, intrinsic tissue regulator of intractable inflammatory diseases. Pathol. Int. 2011, 61, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.; Mistretta, D.; Chambers, A.F.; Krishna, S.; Porter, J.F.; Raghuram, S.; Rittling, S.R. Transcriptional regulation of osteopontin and the metastatic phenotype: evidence for a Ras-activated enhancer in the human OPN promoter. Clin. Exp. Metastasis. 2003, 20, 77–84. [Google Scholar] [CrossRef]
- Rittling, S.R.; Chen, Y.; Feng, F.; Wu, Y. Tumor-derived osteopontin is soluble, not matrix associated. J. Biol. Chem. 2002, 277, 9175–9182. [Google Scholar] [CrossRef]
- Boggio, E.; Dianzani, C.; Gigliotti, C.L. Thrombin Cleavage of Osteopontin Modulates Its Activities in Human Cells In Vitro and Mouse Experimental Autoimmune Encephalomyelitis In Vivo. J. Immunol. Res. 2016, 2016, 9345495. [Google Scholar] [CrossRef] [PubMed]
- Grassinger, J.; Haylock, D.N.; Storan, M.J. Thrombin-cleaved osteopontin regulates hemopoietic stem and progenitor cell functions through interactions with alpha9beta1 and alpha4beta1 integrins. Blood 2009, 114, 49–59. [Google Scholar] [CrossRef]
- Yokasaki, Y.; Sheppard, D. Mapping of the cryptic integrin-binding site in osteopontin suggests a new mechanism by which thrombin can regulate inflammation and tissue repair. Trends Cardiovasc. Med. 2000, 10, 155–159. [Google Scholar] [CrossRef]
- Katagiri, Y.U.; Sleeman, J.; Fujii, H.; Herrlich, P.; Hotta, H.; Tanaka, K.; Chikuma, S.; Yagita, H.; Okumura, K.; Murakami, M.; et al. CD44 variants but not CD44s cooperate with β1-containing integrins to permit cells to bind to osteopontin independently of arginine-glycine-aspartic acid, thereby stimulating cell motility and chemotaxis. Cancer Res. 1999, 59, 219–226. [Google Scholar]
- Weber, G.F. The metastasis gene osteopontin: A candidate target for cancer therapy. Biochim. Biophys. Acta 2001, 1552, 61–85. [Google Scholar] [CrossRef]
- Furger, K.A.; Menon, R.K.; Tuck, A.B.; Bramwell, V.H.C.; Chambers, A.F. The functional and clinical roles of osteopontin in cancer and metastasis. Curr. Mol. Med. 2001, 1, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.D.; Swiderska-Syn, M.; Dollé, L.; Reid, D.; Eksteen, B.; Claridge, L.; Briones-Orta, M.A.; Shetty, S.; Oo, Y.H.; Riva, A.; et al. Osteopontin neutralisation abrogates the liver progenitor cell response and fibrogenesis in mice. Gut. 2015, 64, 1120–1131. [Google Scholar] [CrossRef]
- Shimada, M.; Greer, P.A.; McMahon, A.P.; Bouxsein, M.L.; Schipani, E. In vivo targeted deletion of calpain small subunit, Capn4, in cells of the osteoblast lineage impairs cell proliferation, differentiation, and bone formation. J. Biol. Chem. 2008, 283, 21002–21010. [Google Scholar] [CrossRef]
- Shinohara, M.L.; Kim, J.H.; Garcia, V.A.; Cantor, H. Engagement of the type I interferon receptor on dendritic cells inhibits T helper 17 cell development: Role of intracellular osteopontin. Immunity 2008, 29, 68–78. [Google Scholar] [CrossRef]
- Young, M.F.; Kerr, J.M.; Termine, J.D.; Wewer, U.M.; Wang, M.G.; McBride, O.W.; Fisher, L.W. cDNA cloning, mRNA distribution and heterogeneity, chromosomal location, and RFLP analysis of human osteopontin (OPN). Genomics 1990, 7, 491–502. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a means to cope with environmental insults: Regulation of inflammation, tissue remodeling, and cell survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- Junaid, A.; Moon, M.C.; Harding, G.E.; Zahradka, P. Osteopontin localizes to the nucleus of 293 cells and associates with polo-like kinase-1. Am. J. Physiol. Cell Physiol. 2007, 292, C919–C926. [Google Scholar] [CrossRef] [PubMed]
- O’Regan, A.; Berman, J.S. Osteopontin: A key cytokine in cell-mediated and granulomatous inflammation. Int. J. Exp. Pathol. 2000, 81, 373–390. [Google Scholar] [CrossRef]
- Lin, J.; Myers, A.L.; Wang, Z.; Nancarrow, D.J.; Ferrer-Torres, D.; Handlogten, A.; Leverenz, K.; Bao, J.; Thomas, D.G.; Wang, T.D.; et al. Osteopontin (OPN/SPP1) isoforms collectively enhance tumor cell invasion and dissemination in esophageal adenocarcinoma. Oncotarget 2015, 6, 22239. [Google Scholar] [CrossRef]
- Shinohara, M.L.; Kim, H.J.; Kim, J.H.; Garcia, V.A.; Cantor, H. Alternative translation of osteopontin generates intracellular and secreted isoforms that mediate distinct biological activities in dendritic cells. Proc. Natl. Acad. Sci. USA 2008, 105, 7235–7239. [Google Scholar] [CrossRef]
- Shinohara, M.L.; Lu, L.; Bu, J. Osteopontin expression is essential for interferon-α production by plasmacytoid dendritic cells. Nat. Immunol. 2006, 7, 498–506. [Google Scholar] [CrossRef]
- Suzuki, K.; Zhu, B.; Rittling, S.R. Colocalization of intracellular osteopontin with CD44 is associated with migration, cell fusion, and resorption in osteoclasts. J. Bone Min. Res. 2002, 17, 1486–1497. [Google Scholar] [CrossRef]
- Zohar, R.; Suzuki, N.; Suzuki, K. Intracellular osteopontin is an integral component of the CD44-ERM complex involved in cell migration. J. Cell Physiol. 2000, 184, 118–130. [Google Scholar] [CrossRef]
- Wung, J.K.; Perry, G.; Kowalski, A. Increased expression of the remodeling- and tumorigenic-associated factor osteopontin in pyramidal neurons of the Alzheimer’s disease brain. Curr. Alzheimer Res. 2007, 4, 67–72. [Google Scholar] [CrossRef]
- Christensen, B.; Kazanecki, C.C.; Petersen, T.E.; Rittling, S.R.; Denhardt, D.T.; Sørensen, E.S. Cell type-specific posttranslational modifications of mouse osteopontin are associated with different adhesive properties. J. Biol. Chem. 2007, 282, 19463–19472. [Google Scholar] [CrossRef]
- He, B.; Mirza, M.; Weber, G.F. An osteopontin splice variant induces anchorage independence in human breast cancer cells. Oncogene 2006, 25, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Shanmugam, V.; Chackalaparampil, I.; Kundu, G.C.; Mukherjee, A.B.; Mukherjee, B.B. Altered sialylation of osteopontin prevents its receptor-mediated binding on the surface of oncogenically transformed tsB77 cells. Biochemistry 1997, 36, 5729–5738. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.; Zachariae, E.D.; Scavenius, C.; Thybo, M.; Callesen, M.M.; Kløverpris, S.; Oxvig, C.; Enghild, J.J.; Sorensen, E.S. Identification of transglutaminase reactive residues in human osteopontin and their role in polymerization. PLoS ONE 2014, 9, 113650. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.S.; Greenberg, C.S. TGM2 and implications for human disease: Role of alternative splicing. Front. Biosci. 2013, 18, 504–519. [Google Scholar]
- Lorand, L.; Graham, R.M. Transglutaminases: Crosslinking enzymes with pleiotropic functions. Nat. Rev. Mol. Cell Biol. 2003, 4, 140–156. [Google Scholar] [CrossRef]
- Kaartinen, M.T.; El-Maadawy, S.; Räsänen, N.H.; McKee, M.D. Tissue transglutaminase and its substrates in bone. J. Bone Miner. Res. 2002, 17, 2161–2173. [Google Scholar] [CrossRef]
- Kaartinen, M.T.; Murshed, M.; Karsenty, G.; McKee, M.D. Osteopontin upregulation and polymerization by transglutaminase 2 in calcified arteries of Matrix Gla protein-deficient mice. J. Histochem. Cytochem. 2007, 55, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Patarca, R.; Schwartz, J.; Singh, P.; Cantor, H. Definition of a specific interaction between the early T lymphocyte activation 1 (Eta-1) protein and murine macrophages in vitro and its effect upon macrophages in vivo. J. Exp. Med. 1990, 171, 1931–1942. [Google Scholar] [CrossRef]
- Nishimichi, N.; Hayashita-Kinoh, H.; Chen, C.; Matsuda, H.; Sheppard, D.; Yokosaki, Y. Osteopontin undergoes polymerization in vivo and gains chemotactic activity for neutrophils mediated by integrin α9β1. J. Biol. Chem. 2011, 286, 11170–11178. [Google Scholar] [CrossRef]
- Senger, D.R.; Perruzzi, C.A. Cell migration promoted by a potent GRGDS-containing thrombin-cleavage fragment of osteopontin. Biochim. Biophys. Acta 1996, 1314, 13–24. [Google Scholar] [CrossRef]
- Green, M.P.; Ludbrook, S.B.; Miller, D.D.; Horgan, C.M.T.; Barry, S.T. Structural elements of the osteopontin SVVYGLR motif important for the interaction with alpha(4) integrins. FEBS Lett. 2001, 503, 75–79. [Google Scholar] [CrossRef]
- Smith, L.L.; Cheung, H.K.; Ling, L.E. Osteopontin N-terminal domain contains a cryptic adhesive sequence recognized by alpha9beta1 integrin. J. Biol. Chem. 1996, 271, 28485–28491. [Google Scholar] [CrossRef]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The integrin α9β1 binds to a novel recognition sequence (SVVYGLR) in the thrombin-cleaved amino-terminal fragment of osteopontin. J. Biol. Chem. 1999, 274, 36328–36334. [Google Scholar] [CrossRef]
- Triantafilou, K.; Kar, S.; Vakakis, E.; Kotecha, S.; Triantafilou, M. Human respiratory syncytial virus viroporin SH: A viral recognition pathway used by the host to signal inflammasome activation. Thorax 2013, 68, 66–75. [Google Scholar] [CrossRef]
- Bai, F.; Thompson, E.A.; Vig, P.J.S.; Leis, A.A. Current Understanding of West Nile Virus Clinical Manifestations, Immune Responses, Neuroinvasion, and Immunotherapeutic Implications. Pathogens 2019, 8, 193. [Google Scholar] [CrossRef]
- Paul, A.M.; Acharya, D.; Duty, L. Osteopontin facilitates West Nile virus neuroinvasion via neutrophil “Trojan horse” transport. Sci. Rep. 2017, 7, 4722. [Google Scholar] [CrossRef]
- Bortell, N.; Flynn, C.; Conti, B.; Fox, H.S.; Marcondes, M.C.G. Osteopontin Impacts West Nile virus Pathogenesis and Resistance by Regulating Inflammasome Components and Cell Death in the Central Nervous System at Early Time Points. Mediat. Inflamm. 2017, 2017, 7582437. [Google Scholar] [CrossRef]
- Luissint, A.C.; Artus, C.; Glacial, F.; Ganeshamoorthy, K.; Couraud, P.O. Tight junctions at the blood brain barrier: Physiological architecture and disease-associated dysregulation. Fluids Barriers CNS 2012, 9, 23. [Google Scholar] [CrossRef]
- Zhao, Z.; Nelson, A.R.; Betsholtz, C.; Zlokovic, B.V. Establishment and Dysfunction of the Blood-Brain Barrier. Cell 2015, 163, 1064–1078. [Google Scholar] [CrossRef]
- Diamond, M.S.; Klein, R.S. West Nile virus: Crossing the blood-brain barrier. Nat. Med. 2004, 10, 1294–1295. [Google Scholar] [CrossRef]
- Samuel, M.A.; Diamond, M.S. Pathogenesis of West Nile Virus infection: A balance between virulence, innate and adaptive immunity, and viral evasion. J. Virol. 2006, 80, 9349–9360. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.A.; Wang, H.; Siddharthan, V.; Morrey, J.D.; Diamond, M.S. Axonal transport mediates West Nile virus entry into the central nervous system and induces acute flaccid paralysis. Proc. Natl. Acad. Sci. USA 2007, 104, 17140–17145. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Dai, J.; Bai, F. Matrix metalloproteinase 9 facilitates West Nile virus entry into the brain. J. Virol 2008, 82, 8978–8985. [Google Scholar] [CrossRef]
- Agrawal, S.; Anderson, P.; Durbeej, M. Dystroglycan is selectively cleaved at the parenchymal basement membrane at sites of leukocyte extravasation in experimental autoimmune encephalomyelitis. J. Exp. Med. 2006, 203, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Esparza, J.; Kruse, M.; Lee, J.; Michaud, M.; Madri, J.A. MMP-2 null mice exhibit an early onset and severe experimental autoimmune encephalomyelitis due to an increase in MMP-9 expression and activity. FASEB J. 2004, 18, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Romanic, A.M.; Madri, J.A. Extracellular matrix-degrading proteinases in the nervous system. Brain Pathol. 1994, 4, 145–156. [Google Scholar] [CrossRef]
- Brilha, S.; Ong, C.W.M.; Weksler, B.; Romero, N.; Couraud, P.O.; Friedland, J.S. Matrix metalloproteinase-9 activity and a downregulated Hedgehog pathway impair blood-brain barrier function in an in vitro model of CNS tuberculosis. Sci. Rep. 2017, 7, 16031. [Google Scholar] [CrossRef]
- Tang, J.; Kang, Y.; Huang, L.; Wu, L.; Peng, Y. TIMP1 preserves the blood–brain barrier through interacting with CD63/integrin β1 complex and regulating downstream FAK/RhoA signaling. Acta Pharm. Sin. B 2020, 10, 987–1003. [Google Scholar] [CrossRef]
- Rangaswami, H.; Kundu, G.C. Osteopontin stimulates melanoma growth and lung metastasis through NIK/MEKK1-dependent MMP-9 activation pathways. Oncol. Rep. 2007, 18, 909–915. [Google Scholar] [CrossRef]
- Lindsey, M.L.; Zouein, F.A.; Tian, Y.; Padmanabhan, I.R.; de Castro Brás, L.E. Osteopontin is proteolytically processed by matrix metalloproteinase 9. Can. J. Physiol. Pharmacol. 2015, 93, 879–886. [Google Scholar] [CrossRef]
- Chan, J.L.; Reeves, T.M.; Phillips, L.L. Osteopontin expression in acute immune response mediates hippocampal synaptogenesis and adaptive outcome following cortical brain injury. Exp. Neurol. 2014, 261, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Doyle, K.P.; Yang, T.; Lessov, N.S. Nasal administration of osteopontin peptide mimetics confers neuroprotection in stroke. J. Cereb. Blood Flow Metab. 2008, 28, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Meller, R.; Stevens, S.L.; Minami, M. Neuroprotection by osteopontin in stroke. J. Cereb. Blood Flow Metab. 2005, 25, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Ayer, R.; Sugawara, T. Protective effects of recombinant osteopontin on early brain injury after subarachnoid hemorrhage in rats. Crit. Care Med. 2010, 38, 612–618. [Google Scholar] [CrossRef]
- Shin, T. Osteopontin as a two-sided mediator in acute neuroinflammation in rat models. Acta Histochem. 2012, 114, 749–754. [Google Scholar] [CrossRef]
- Bai, F.; Kong, K.F.; Dai, J. A paradoxical role for neutrophils in the pathogenesis of West Nile virus. J. Infect. Dis. 2010, 202, 1804–1812. [Google Scholar] [CrossRef]
- Beasley, D.W.; Li, L.; Suderman, M.T.; Barrett, A.D. Mouse neuroinvasive phenotype of West Nile virus strains varies depending upon virus genotype. Virology 2002, 296, 17–23. [Google Scholar] [CrossRef]
- Roulston, A.; Marcellus, R.C.; Branton, P.E. Viruses and apoptosis. Annu. Rev. Microbiol. 1999, 53, 577–628. [Google Scholar] [CrossRef]
- Song, G.; Ming, Y.; Mao, Y.; Bao, S.; Ouyang, G. Osteopontin Prevents Curcumin-Induced Apoptosis and Promotes Survival Through Akt Activation via αvβ3 Integrins in Human Gastric Cancer Cells. Exp. Biol. Med. 2008, 233, 1537–1545. [Google Scholar] [CrossRef]
- Iida, T.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? Int. J. Mol. Sci. 2017, 19, 7. [Google Scholar] [CrossRef]
- Rittling, R.S.; Chambers, A.F. Role of osteopontin in tumour progression. Br. J. Cancer 2004, 90, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Saleh, S.; Thompson, D.E.; McConkey, J.; Murray, P.; Moorehead, R.A. Osteopontin regulates proliferation, apoptosis, and migration of murine claudin-low mammary tumor cells. BMC Cancer 2016, 16, 359. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, A.M.; Hocker, S.E.; Leis, A.A. West Nile virus induces a post-infectious pro-inflammatory state that explains transformation of stable ocular myasthenia gravis to myasthenic crises. J. Neurol. Sci. 2018, 395, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Leis, A.A.; Szatmary, G.; Ross, M.A.; Stokic, D.S. West nile virus infection and myasthenia gravis. Muscle Nerve 2014, 49, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Segovia, J.; Sabbah, A.; Mgbemena, V. TLR2/MyD88/NF-κB pathway, reactive oxygen species, potassium efflux activates NLRP3/ASC inflammasome during respiratory syncytial virus infection. PLoS ONE 2012, 7, e29695. [Google Scholar] [CrossRef] [PubMed]
- Han, R.K.; Cheng, Y.F.; Zhou, S.S. Increased circulating Th17 cell populations and elevated CSF osteopontin and IL-17 concentrations in patients with Guillain-Barré syndrome. J. Clin. Immunol. 2014, 34, 94–103. [Google Scholar] [CrossRef]
- Hassin-Baer, S.; Kirson, E.D.; Shulman, L. Stiff-person syndrome following West Nile fever. Arch. Neurol. 2004, 61, 938–941. [Google Scholar] [CrossRef]
- Leis, A.A.; Stokic, D.S. Neuromuscular manifestations of west nile virus infection. Front. Neurol. 2012, 3, 37. [Google Scholar] [CrossRef]
- Almhanna, K.; Palanichamy, N.; Sharma, M.; Hobbs, R.; Sil, A. Unilateral brachial plexopathy associated with West Nile virus meningoencephalitis. Clin. Infect. Dis. 2003, 36, 1629–1630. [Google Scholar] [CrossRef]
- Yu, H.; Liu, X.; Zhong, Y. The Effect of Osteopontin on Microglia. BioMed Res. Int. 2017, 2017, 1879437. [Google Scholar] [CrossRef]
- Clemente, N.; Raineri, D.; Cappellano, G. Osteopontin Bridging Innate and Adaptive Immunity in Autoimmune Diseases. J. Immunol. Res. 2016, 2016, 7675437. [Google Scholar] [CrossRef] [PubMed]
- Frenzel, D.F.; Borkner, L.; Scheurmann, J.; Singh, K.; Scharffetter-Kochanek, K.; Weiss, J.M. Osteopontin deficiency affects imiquimod-induced psoriasis-like murine skin inflammation and lymphocyte distribution in skin, draining lymph nodes and spleen. Exp. Derm. 2015, 24, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Hur, E.M.; Youssef, S.; Haws, M.E.; Zhang, S.Y.; Sobel, R.A.; Steinman, L. Osteopontin-induced relapse and progression of autoimmune brain disease through enhanced survival of activated T cells. Nat. Immunol. 2007, 8, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Kon, S.; Nakayama, Y.; Matsumoto, N.; Ito, K.; Kanayama, M.; Kimura, C.; Kouro, H.; Ashitomi, D.; Matsuda, T.; Uede, T. A novel cryptic binding motif, LRSKSRSFQVSDEQY, in the C-terminal fragment of MMP-3/7-cleaved osteopontin as a novel ligand for α9β1 integrin is involved in the anti-type II collagen antibody-induced arthritis. PLoS ONE 2014, 9, e116210. [Google Scholar] [CrossRef] [PubMed]
- Rittling, R.S.; Singh, R. Osteopontin in Immune-mediated Diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef]
- Phillips, D.W.; Vincent, A. Pathogenesis of myasthenia gravis: Update on disease types, models, and mechanisms. F1000Research 2016, 5, 1513. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazneen, F.; Bai, F. The Roles of Osteopontin in the Pathogenesis of West Nile Encephalitis. Vaccines 2020, 8, 748. https://doi.org/10.3390/vaccines8040748
Nazneen F, Bai F. The Roles of Osteopontin in the Pathogenesis of West Nile Encephalitis. Vaccines. 2020; 8(4):748. https://doi.org/10.3390/vaccines8040748
Chicago/Turabian StyleNazneen, Farzana, and Fengwei Bai. 2020. "The Roles of Osteopontin in the Pathogenesis of West Nile Encephalitis" Vaccines 8, no. 4: 748. https://doi.org/10.3390/vaccines8040748
APA StyleNazneen, F., & Bai, F. (2020). The Roles of Osteopontin in the Pathogenesis of West Nile Encephalitis. Vaccines, 8(4), 748. https://doi.org/10.3390/vaccines8040748