Anti-Influenza Effect of Nanosilver in a Mouse Model

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
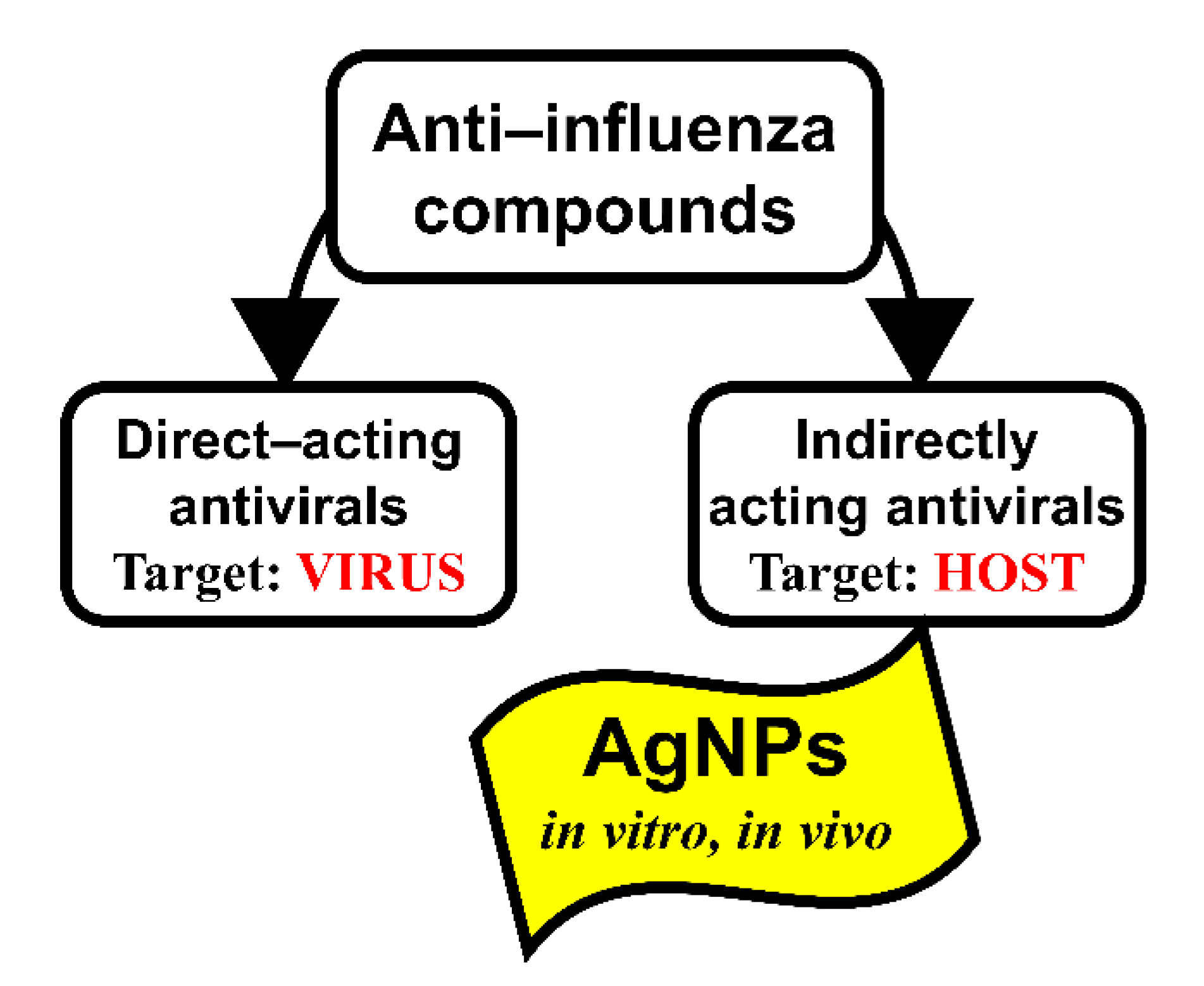
1. Introduction
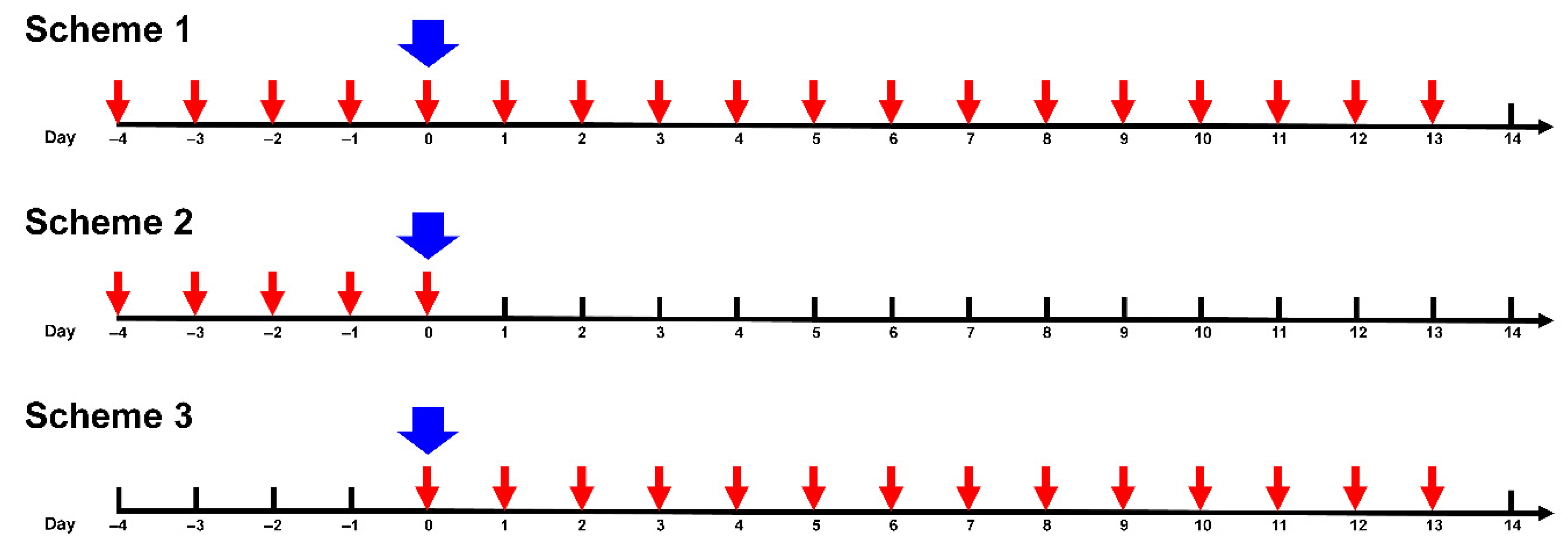
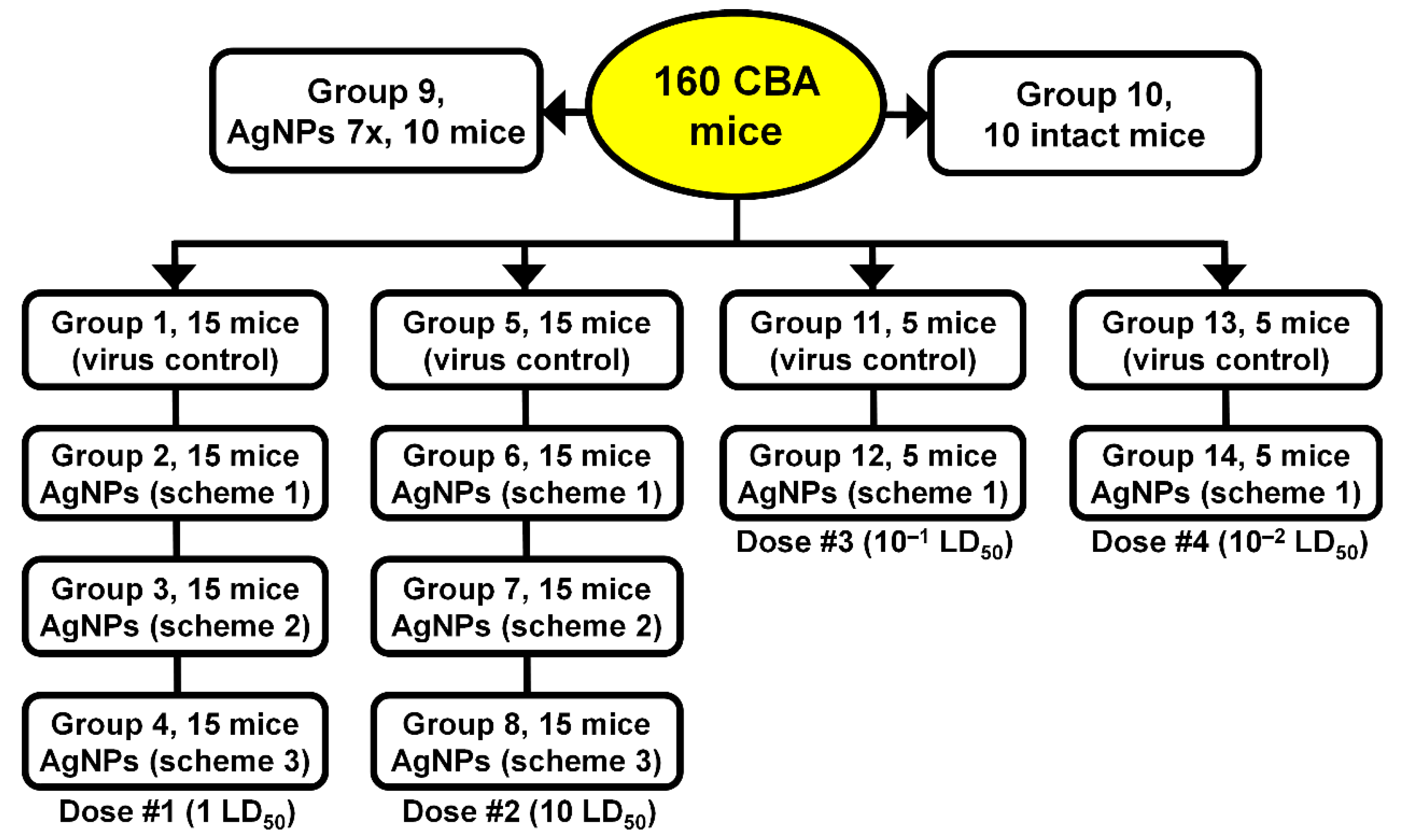
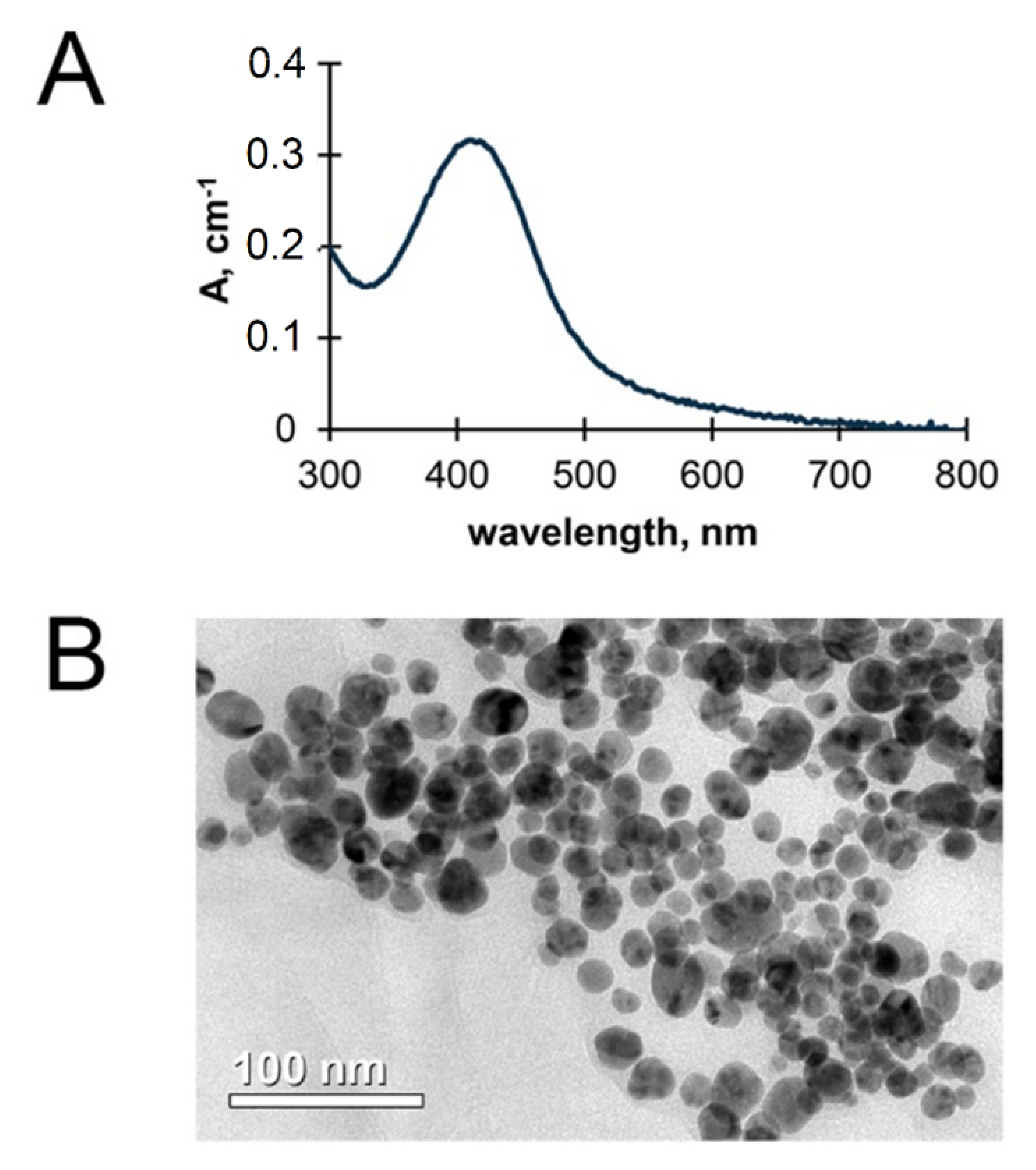
2. Materials and Methods
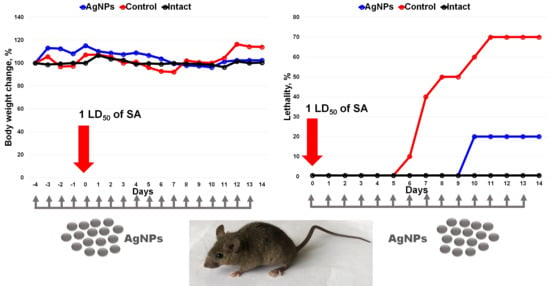
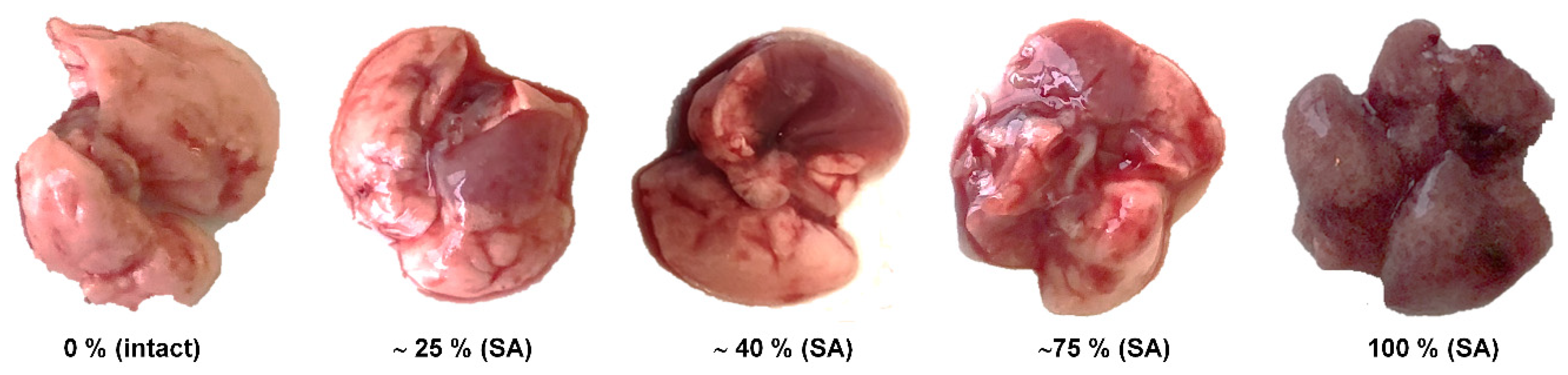
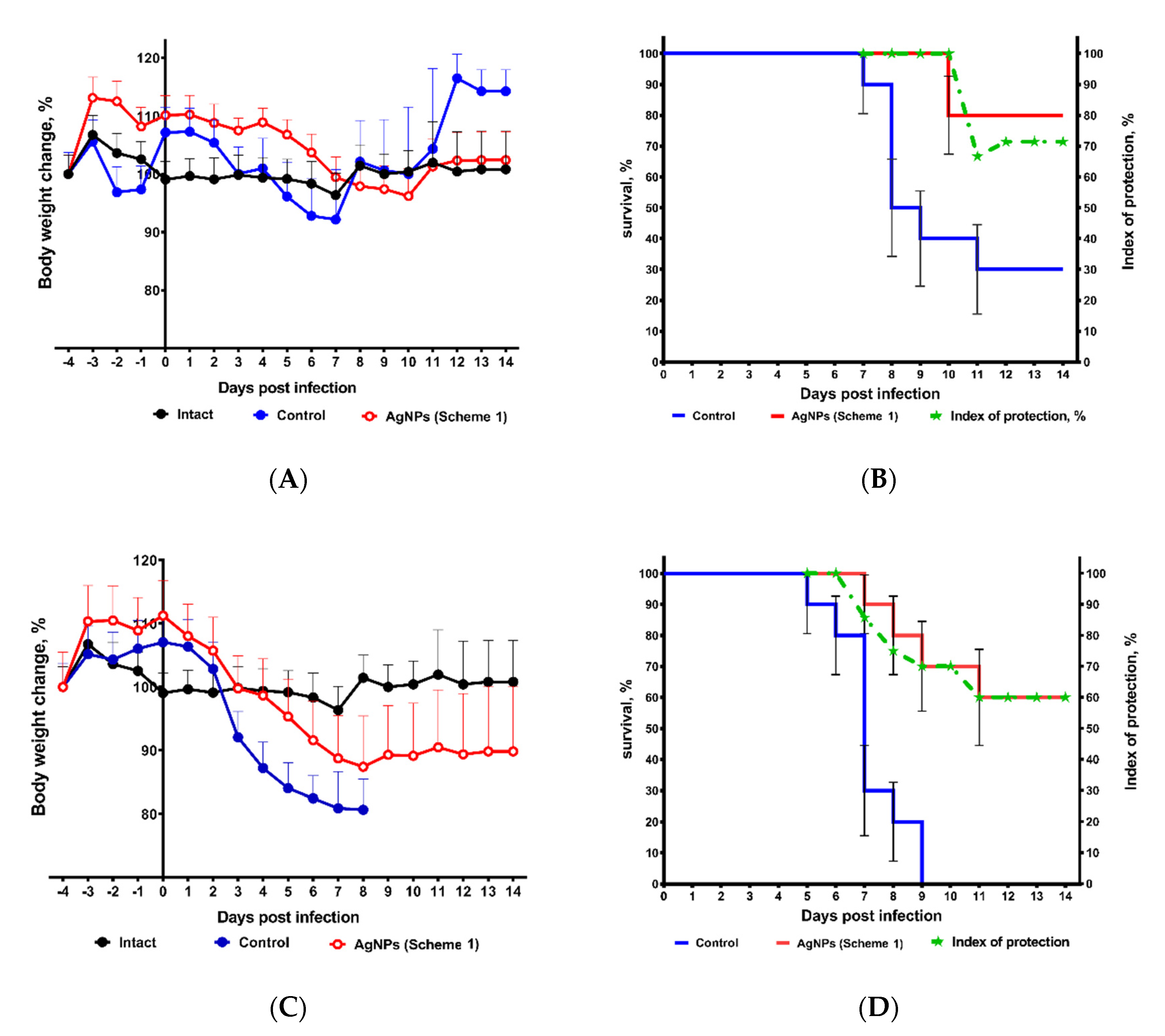
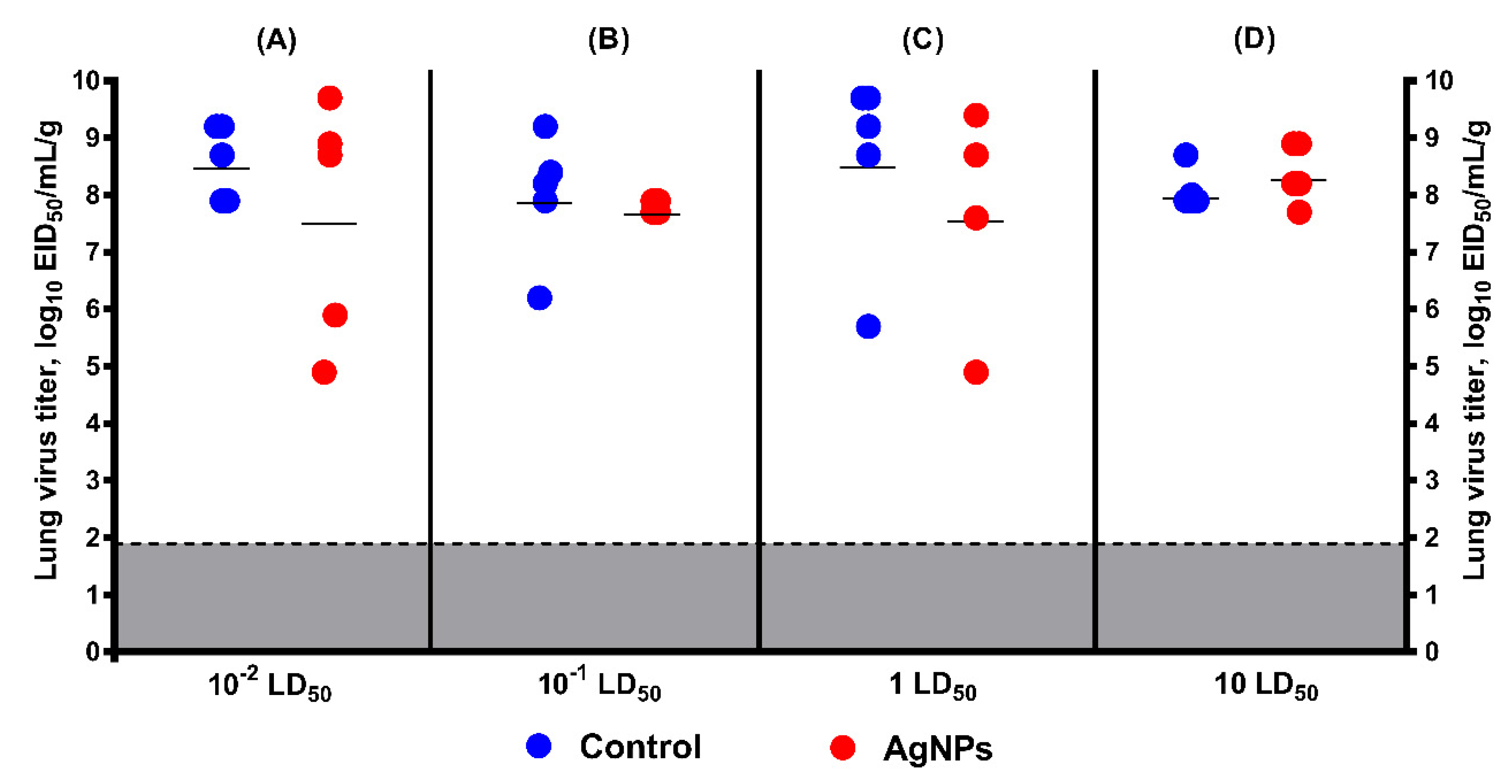
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- CDC. Prevention and Control of Seasonal Influenza with Vaccines: Recommendations of the Advisory Committee on Immunization Practices—United States, 2019–2020 Influenza Season. Available online: https://www.cdc.gov/mmwr/volumes/68/rr/rr6803a1.htm (accessed on 5 October 2020).
- NVAC. Strategies to achieve the healthy people 2020 annual influenza vaccine coverage goal for health-care personnel: Recommendations from the National Vaccine Advisory Committee. Public Health Rep. 2013, 128, 7–25. [Google Scholar] [CrossRef]
- WHO. Influenza Vaccines. Available online: https://www.who.int/influenza/vaccines/en/ (accessed on 23 October 2020).
- Mameli, C.; Cocchi, I.; Fumagalli, M.; Zuccotti, G. Influenza vaccination: Effectiveness, indications, and limits in the pediatric population. Front. Pediatr. 2019, 7, 317. [Google Scholar] [CrossRef]
- Shie, J.J.; Fang, J.M. Development of effective anti-influenza drugs: Congeners and conjugates—A review. J. Biomed Sci. 2019, 26, 84. [Google Scholar] [CrossRef] [PubMed]
- Forterre, P. Manipulation of cellular syntheses and the nature of viruses: The virocell concept. C. R. Chim. 2011, 14, 392–399. [Google Scholar] [CrossRef]
- Forterre, P. The virocell concept and environmental microbiology. ISME J. 2013, 7, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Howard-Varona, C.; Lindback, M.M.; Bastien, G.E.; Solonenko, N.; Zayed, A.A.; Jang, H.; Andreopoulos, B.; Brewer, H.M.; Glavina Del Rio, T.; Adkins, J.N.; et al. Phage-specific metabolic reprogramming of virocells. ISME J. 2020, 14, 881–895. [Google Scholar] [CrossRef]
- Tan, S.L.; Ganji, G.; Paeper, B.; Proll, S.; Katze, M.G. Systems biology and the host response to viral infection. Nat. Biotechnol. 2007, 25, 1383–1389. [Google Scholar] [CrossRef]
- Tripathi, S.; Garcia-Sastre, A. Antiviral innate immunity through the lens of systems biology. Virus Res. 2016, 218, 10–17. [Google Scholar] [CrossRef]
- Troy, N.M.; Bosco, A. Respiratory viral infections and host responses; insights from genomics. Respir. Res. 2016, 17, 156. [Google Scholar] [CrossRef]
- Ackerman, C.M.; Chang, C.J. Copper signaling in the brain and beyond. J. Biol. Chem. 2018, 293, 4628–4635. [Google Scholar] [CrossRef]
- Kardos, J.; Héja, L.; Simon, Á.; Jablonkai, I.; Kovács, R.; Jemnitz, K. Copper signalling: Causes and consequences. Cell Commun. Signal. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, L.; Cotruvo, J.A., Jr.; Chan, J.; Kaluarachchi, H.; Muchenditsi, A.; Pendyala, V.S.; Jia, S.; Aron, A.T.; Ackerman, C.M.; Wal, M.N.; et al. Copper regulates cyclic-AMP-dependent lipolysis. Nat. Chem. Biol. 2016, 12, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Silva, A.M.; Matos, M.; Monteiro, S.M.; Álvaro, A.R. Copper induced apoptosis in Caco-2 and Hep-G2 cells: Expression of caspases 3, 8 and 9, AIF and p53. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185–186, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Bondanese, V.P.; Lamboux, A.; Simon, M.; Lafont, J.E.; Albalat, E.; Pichat, S.; Vanacker, J.M.; Telouk, P.; Balter, V.; Oger, P.; et al. Hypoxia induces copper stable isotope fractionation in hepatocellular carcinoma, in a HIF-independent manner. Metallomics 2016, 8, 1177–1184. [Google Scholar] [CrossRef]
- Feng, W.; Ye, F.; Xue, W.; Zhou, Z.; Kang, Y.J. Copper regulation of hypoxia-inducible factor-1 activity. Mol. Pharmacol. 2009, 75, 174–182. [Google Scholar] [CrossRef]
- Yuan, S.; Chen, S.; Xi, Z.; Liu, Y. Copper-finger protein of Sp1: The molecular basis of copper sensing. Metallomics 2017, 9, 1169–1175. [Google Scholar] [CrossRef]
- Xie, H.; Kang, Y.J. Role of copper in angiogenesis and its medicinal implications. Curr. Med. Chem. 2009, 16, 1304–1314. [Google Scholar] [CrossRef]
- D’Ambrosi, N.; Rossi, L. Copper at synapse: Release, binding and modulation of neurotransmission. Neurochem. Int. 2015, 90, 36–45. [Google Scholar] [CrossRef]
- De Luca, A.; Barile, A.; Arciello, M.; Rossi, L. Copper homeostasis as target of both consolidated and innovative strategies of anti-tumor therapy. J. Trace Elem. Med. Biol. 2019, 55, 204–213. [Google Scholar] [CrossRef]
- Haag, F.; Ahmed, L.; Reiss, K.; Block, E.; Batista, V.S.; Krautwurst, D. Copper-mediated thiol potentiation and mutagenesis-guided modeling suggest a highly conserved copper-binding motif in human OR2M3. Cell Mol. Life Sci. 2020, 77, 2157–2179. [Google Scholar] [CrossRef]
- Pavelková, M.; Vysloužil, J.; Kubová, K.; Vetchý, D. Biological role of copper as an essential trace element in the human organism. Ceska Slov. Farm. 2018, 67, 143–153. [Google Scholar]
- Lin, X.; Wang, R.; Zou, W.; Sun, X.; Liu, X.; Zhao, L.; Wang, S.; Jin, M. The influenza virus H5N1 infection can induce ROS production for viral replication and host cell death in A549 cells modulated by human Cu/Zn superoxide dismutase (SOD1) overexpression. Viruses 2016, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Pyo, C.W.; Shin, N.; Jung, K.I.; Choi, J.H.; Choi, S.Y. Alteration of copper-zinc superoxide dismutase 1 expression by influenza A virus is correlated with virus replication. Biochem. Biophys. Res. Commun. 2014, 450, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.; Pyo, C.W.; Jung, K.I.; Choi, S.Y. Influenza A virus PB1-F2 is involved in regulation of cellular redox state in alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2015, 459, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.I.; Pyo, C.W.; Choi, S.Y. Influenza A virus-induced autophagy contributes to enhancement of virus infectivity by SOD1 downregulation in alveolar epithelial cells. Biochem. Biophys. Res. Commun. 2018, 498, 960–966. [Google Scholar] [CrossRef]
- Dagdag, R.P.; Burkhead, J.L.; Bortz, E. Influenza versus host: The role of novel copper-related host factors in antiviral immunity. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Doty, R.L.; Berman, A.H.; Izhar, M.; Hamilton, H.B.; Villano, D.; Vazquez, B.E.; Warrum, M.N.; Mahbob, M. Influenza vaccinations and chemosensory function. Am. J. Rhinol. Allergy 2014, 28, 50–53. [Google Scholar] [CrossRef]
- Moein, S.T.; Hashemian, S.M.; Mansourafshar, B.; Khorram-Tousi, A.; Tabarsi, P.; Doty, R.L. Smell dysfunction: A biomarker for COVID-19. Int. Forum Allergy Rhinol. 2020, 10, 944–950. [Google Scholar] [CrossRef]
- Rupp, J.C.; Locatelli, M.; Grieser, A.; Ramos, A.; Campbell, P.J.; Yi, H.; Steel, J.; Burkhead, J.L.; Bortz, E. Host cell copper transporters CTR1 and ATP7A are important for influenza A virus replication. Virol. J. 2017, 14, 11. [Google Scholar] [CrossRef]
- Chaturvedi, U.C.; Shrivastava, R. Interaction of viral proteins with metal ions: Role in maintaining the structure and functions of viruses. FEMS Immunol. Med. Microbiol. 2005, 43, 105–114. [Google Scholar] [CrossRef]
- Oxford, J.S.; Perrin, D.D. Inhibition of the particle-associated RNA-dependent RNA polymerase activity of influenza viruses by chelating agents. J. Gen. Virol. 1974, 23, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, C.S.; Shuck, K.; Lear, J.D.; Dieckmann, G.R.; DeGrado, W.F.; Lamb, R.A.; Pinto, L.H. Cu (II) inhibition of the proton translocation machinery of the influenza A virus M2 protein. J. Biol. Chem. 1999, 274, 5474–5482. [Google Scholar] [CrossRef] [PubMed]
- Tomas, E.; Samuel, I.; Popescu, A.; Cajal, N. Comparative study of some characteristics of influenza virus A/PR8/34 (H1N1) cultivated on chorioallantoic membrane fragments in the presence of ceruloplasmin or of parainfluenza type I (Sendai) virus. Virologie 1983, 34, 295–301. [Google Scholar] [PubMed]
- Tomas, E.; Topârceanu, F. Considerations about the possible function of ceruloplasmin in influenza and parainfluenza virus infections. Virologie 1986, 37, 279–287. [Google Scholar] [PubMed]
- Van Lenten, B.J.; Wagner, A.C.; Nayak, D.P.; Hama, S.; Navab, M.; Fogelman, A.M. High-density lipoprotein loses its anti-inflammatory properties during acute influenza A infection. Circulation 2001, 103, 2283–2288. [Google Scholar] [CrossRef]
- Vashchenko, G.; MacGillivray, R.T. Multi-copper oxidases and human iron metabolism. Nutrients 2013, 5, 2289–2313. [Google Scholar] [CrossRef]
- Jeffery, C.J. Protein moonlighting: What is it, and why is it important? Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2018, 373. [Google Scholar] [CrossRef]
- Bielli, P.; Calabrese, L. Structure to function relationships in ceruloplasmin: A “moonlighting” protein. Cell Mol. Life Sci. 2002, 59, 1413–1427. [Google Scholar] [CrossRef]
- Das, S.; Sahoo, P.K. Ceruloplasmin, a moonlighting protein in fish. Fish Shellfish Immunol. 2018, 82, 460–468. [Google Scholar] [CrossRef]
- Golenkina, E.A.; Viryasova, G.M.; Galkina, S.I.; Gaponova, T.V.; Sud’ina, G.F.; Sokolov, A.V. Fine regulation of neutrophil oxidative status and apoptosis by ceruloplasmin and its derivatives. Cells 2018, 7, 8. [Google Scholar] [CrossRef]
- Harvey, L.J.; McArdle, H.J. Biomarkers of copper status: A brief update. Br. J. Nutr. 2008, 99, S10–S13. [Google Scholar] [CrossRef] [PubMed]
- Ilyechova, E.; Skvortsov, A.; Zatulovsky, E.; Tsymbalenko, N.; Shavlovsky, M.; Broggini, M.; Puchkova, L. Experimental switching of copper status in laboratory rodents. J. Trace Elem. Med. Biol. 2011, 25, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Puchkova, L.V.; Broggini, M.; Polishchuk, E.V.; Ilyechova, E.Y.; Polishchuk, R.S. Silver ions as a tool for understanding different aspects of copper metabolism. Nutrients 2019, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Le Ouay, B.; Stellacci, F. Antibacterial activity of silver nanoparticles: A surface science insight. Nano Today 2015, 10, 339–354. [Google Scholar] [CrossRef]
- Orlov, I.A.; Sankova, T.P.; Babich, P.S.; Sosnin, I.M.; Ilyechova, E.Y.; Kirilenko, D.A.; Brunkov, P.N.; Ataev, G.L.; Romanov, A.E.; Puchkova, L.V. New silver nanoparticles induce apoptosis-like process in E. coli and interfere with mammalian copper metabolism. Int. J. Nanomed. 2016, 11, 6561–6574. [Google Scholar] [CrossRef]
- Ilyechova, E.Y.; Saveliev, A.N.; Skvortsov, A.N.; Babich, P.S.; Zatulovskaia, Y.A.; Pliss, M.G.; Korzhevskii, D.E.; Tsymbalenko, N.V.; Puchkova, L.V. The effects of silver ions on copper metabolism in rats. Metallomics 2014, 6, 1970–1987. [Google Scholar] [CrossRef]
- Zatulovskiy, E.A.; Skvortsov, A.N.; Rusconi, P.; Ilyechova, E.Y.; Babich, P.S.; Tsymbalenko, N.V.; Broggini, M.; Puchkova, L.V. Serum depletion of holo-ceruloplasmin induced by silver ions in vivo reduces uptake of cisplatin. J. Inorg. Biochem. 2012, 116, 88–96. [Google Scholar] [CrossRef]
- Rai, M.; Deshmukh, S.D.; Ingle, A.P.; Gupta, I.R.; Galdiero, M.; Galdiero, S. Metal nanoparticles: The protective nanoshield against virus infection. Crit. Rev. Microbiol. 2016, 42, 46–56. [Google Scholar] [CrossRef]
- Lara, H.H.; Garza-Treviño, E.N.; Ixtepan-Turrent, L.; Singh, D.K. Silver nanoparticles are broad-spectrum bactericidal and virucidal compounds. J. Nanobiotechnol. 2011, 9, 30. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tavakoli, A.; Moradi, A.; Tabarraei, A.; Bokharaei-Salim, F.; Zahmatkeshan, M.; Farahmand, M.; Javanmard, D.; Kiani, S.J.; Esghaei, M.; et al. Inhibition of H1N1 influenza virus infection by zinc oxide nanoparticles: Another emerging application of nanomedicine. J. Biomed. Sci 2019, 26, 70. [Google Scholar] [CrossRef]
- Xiang, D.X.; Chen, Q.; Pang, L.; Zheng, C.L. Inhibitory effects of silver nanoparticles on H1N1 influenza A virus in vitro. J. Virol. Methods 2011, 178, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Mehrbod, P.; Motamed, N.; Tabatabaian, M.; Soleimani Estyar, R.; Amini, E.; Shahidi, M.; Kheiri, M.T. In vitro antiviral effect of “nanosilver“ on influenza virus. DARU J. Pharm. Sci. 2008, 17, 88–93. [Google Scholar]
- Xiang, D.; Zheng, Y.; Duan, W.; Li, X.; Yin, J.; Shigdar, S.; O’Connor, M.L.; Marappan, M.; Zhao, X.; Miao, Y.; et al. Inhibition of A/Human/Hubei/3/2005 (H3N2) influenza virus infection by silver nanoparticles in vitro and in vivo. Int. J. Nanomed. 2013, 8, 4103–4113. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, I.; Rekstin, A.; Al Farroukh, M.; Bazhenova, E.; Katelnikova, A.; Puchkova, L.; Rudenko, L. Non-mouse-adapted H1N1pdm09 virus as a model for influenza research. Viruses 2020, 12, 590. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- European_Council. Directive 2010/63/EU of the European Parliament and of the Council of September 22, 2010, on the Protection of Animals Used for Scientific Purposes. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32010L0063 (accessed on 4 October 2020).
- Karkishenko, N.N.; Grachev, C.V. Manual for Laboratory Animals and Alternative Models in Biomedical Research; Profile-2C: Moscow, Russia, 2010; p. 358. Available online: http://www.scbmt.ru/mag/rukovodstvo.pdf (accessed on 4 October 2020).
- Owen, J.; Smith, H. Detection of ceruloplasmin after zone electrophoresis. Clin. Chim. Acta 1961, 6, 441–444. [Google Scholar] [CrossRef]
- Cabrera, A.; Alonzo, E.; Sauble, E.; Chu, Y.L.; Nguyen, D.; Linder, M.C.; Sato, D.S.; Mason, A.Z. Copper binding components of blood plasma and organs, and their responses to influx of large doses of (65) Cu, in the mouse. Biometals 2008, 21, 525–543. [Google Scholar] [CrossRef]
- Bernevic, B.; El-Khatib, A.H.; Jakubowski, N.; Weller, M.G. Online immunocapture ICP-MS for the determination of the metalloprotein ceruloplasmin in human serum. BMC Res. Notes 2018, 11, 213. [Google Scholar] [CrossRef]
- Moradi-Sardareh, H.; Basir, H.R.G.; Hassan, Z.M.; Davoudi, M.; Amidi, F.; Paknejad, M. Toxicity of silver nanoparticles on different tissues of Balb/C mice. Life Sci. 2018, 211, 81–90. [Google Scholar] [CrossRef]
- Sankova, T.P.; Orlov, I.A.; Sosnin, I.; Kirilenko, D.A.; Skomorokhova, E.A.; Puchkova, L. Influence of silver nanoparticles (SNPs) with different size and shape on copper status in mice. FEBS Open Bio. 2018, 8, 480. [Google Scholar] [CrossRef]
- Małaczewska, J. The in vitro effect of commercially available noble metal nanocolloids on the splenocyte proliferative response and cytokine production in mice. Pol. J. Vet. Sci. 2014, 17, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Gevkaliuk, N.O.; Sydliaruk, N.I.; Posolenyk, L.Y.; Vydoinyk, O.Y.; Kuchyrka, L.I. The state of oxidative homeostasis in children with influenza stomatitis. Wiadomosci Lekarskie 2019, 72, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Kurrer, M.; Bachmann, M.F.; Kopf, M. Interleukin-1 is responsible for acute lung immunopathology but increases survival of respiratory influenza virus infection. J. Virol. 2005, 79, 6441–6448. [Google Scholar] [CrossRef] [PubMed]
- Lauder, S.N.; Jones, E.; Smart, K.; Bloom, A.; Williams, A.S.; Hindley, J.P.; Ondondo, B.; Taylor, P.R.; Clement, M.; Fielding, C.; et al. Interleukin-6 limits influenza-induced inflammation and protects against fatal lung pathology. Eur. J. Immunol. 2013, 43, 2613–2625. [Google Scholar] [CrossRef] [PubMed]
- Dutta, M.; Dutta, P.; Medhi, S.; Borkakoty, B.; Biswas, D. Immune response during influenza virus infection among the population of Assam, Northeast India. Indian J. Med. Microbiol. 2019, 37, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Gitlin, J.D.; Schroeder, J.J.; Lee-Ambrose, L.M.; Cousins, R.J. Mechanisms of caeruloplasmin biosynthesis in normal and copper-deficient rats. Biochem. J. 1992, 282, 835–839. [Google Scholar] [CrossRef]
- Di Bella, L.M.; Alampi, R.; Biundo, F.; Toscano, G.; Felice, M.R. Copper chelation and interleukin-6 proinflammatory cytokine effects on expression of different proteins involved in iron metabolism in HepG2 cell line. BMC Biochem. 2017, 18, 1. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Lowen, A.C. Animal models for influenza virus pathogenesis and transmission. Viruses 2010, 2, 1530–1563. [Google Scholar] [CrossRef]
- Margine, I.; Krammer, F. Animal models for influenza viruses: Implications for universal vaccine development. Pathogens 2014, 3, 845–874. [Google Scholar] [CrossRef]
- Zarubaev, V.V.; Garshinina, A.V.; Kalinina, N.A.; Shtro, A.A.; Belyaevskaya, S.V.; Slita, A.V.; Nebolsin, V.E.; Kiselev, O.I. Activity of ingavirin (6-[2-(1H-Imidazol-4-yl)ethylamino]-5-oxo-hexanoic Acid) against human respiratory viruses in in vivo experiments. Pharmaceuticals 2011, 4, 1518–1534. [Google Scholar] [CrossRef]
- Nakajima, N.; Van Tin, N.; Sato, Y.; Thach, H.N.; Katano, H.; Diep, P.H.; Kumasaka, T.; Thuy, N.T.; Hasegawa, H.; San, L.T.; et al. Pathological study of archival lung tissues from five fatal cases of avian H5N1 influenza in Vietnam. Mod. Pathol. 2013, 26, 357–369. [Google Scholar] [CrossRef] [PubMed]
- El Kebir, D.; Filep, J.G. Role of neutrophil apoptosis in the resolution of inflammation. Sci. World J. 2010, 10, 1731–1748. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Maines, T.R.; Creager, H.M.; Katz, J.M.; Tumpey, T.M. Oseltamivir inhibits influenza virus replication and transmission following ocular-only aerosol inoculation of ferrets. Virology 2015, 484, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Stephen, E.L.; Dominik, J.W.; Moe, J.B.; Spertzel, R.O.; Walker, J.S. Treatment of influenza infection of mice by using rimantadine hydrochlorides by the aerosol and intraperitoneal routes. Antimicrob. Agents Chemother. 1975, 8, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Toots, M.; Plemper, R.K. Next-generation direct-acting influenza therapeutics. Transl. Res. 2020. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
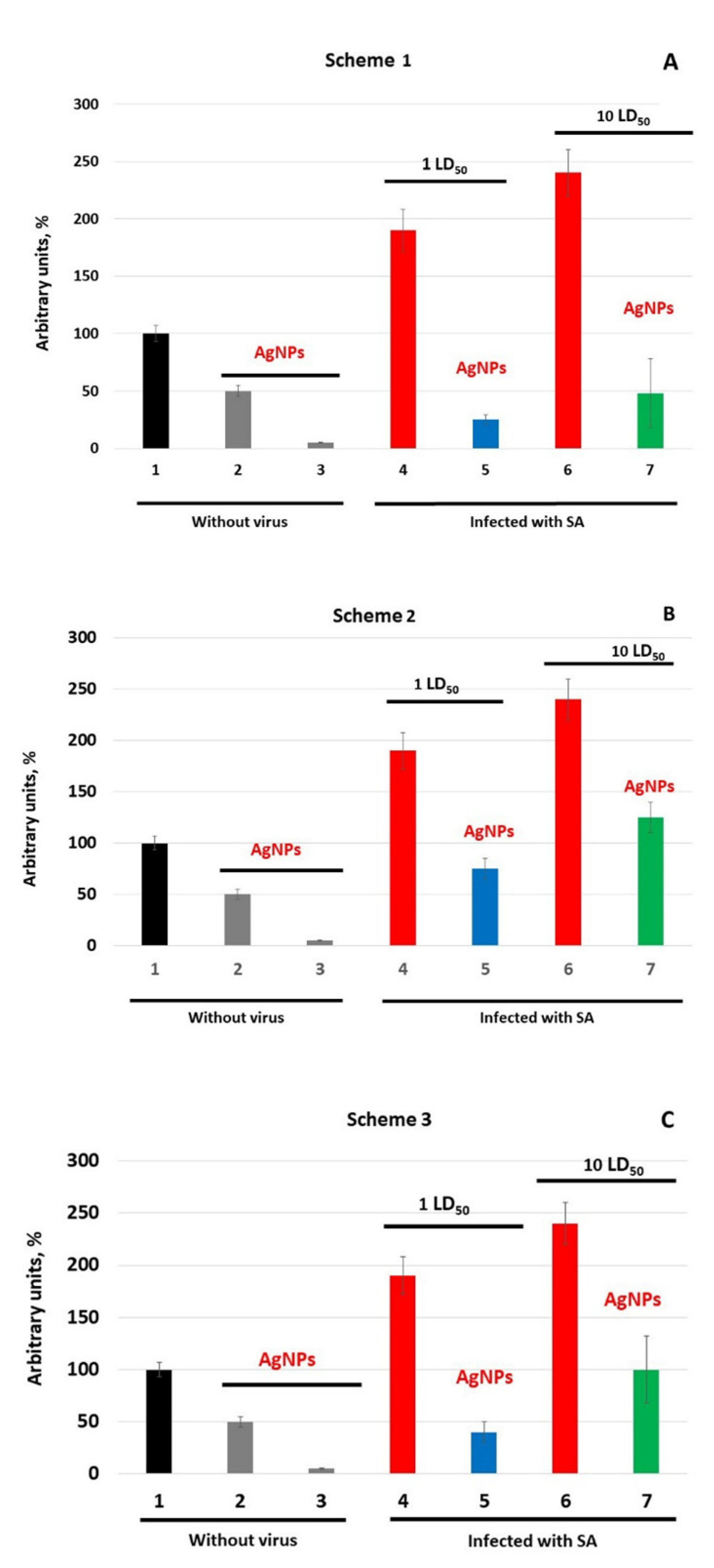{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Group | Metal Concentration, µM | |||
|---|---|---|---|---|
| 3 Days After Infection | At the End of Experiment | |||
| [Cu] | [Ag] | [Cu] | [Ag] | |
| Intact | 9.4 ± 0.47 | 0 | - | - |
| AgNP treatment during | ||||
| 4 days | 7.3 ± 0.24 | 1.9 ± 0.01 | - | - |
| 7 days | 4.7 ± 0.19 | 2.8 ± 0.11 | - | - |
| The viral dose of 1 LD50 per mouse | ||||
| Control | 14.6 ± 0.53 | - | - | - |
| Scheme 1 | 6.5 ± 0.21 | 1.2 ± 0.03 | 9.3 | 3.7 |
| Scheme 2 | 12.4 ± 0.56 | 1.3 ± 0.06 | 12.7 | 0.25 |
| Scheme 3 | 12.8 ± 0.39 | 1.9 ± 0.03 | 8.8 | 3.2 |
| The viral dose of 10 LD50 per mouse | ||||
| control | 15.3 ± 0.33 | - | - | - |
| Scheme 1 | 7.4 ± 0.69 | 0.9 ± 0.02 | 8.3 | 2.3 |
| Scheme 2 | 12.8 ± 0.53 | 1.3 ± 0.05 | - | - |
| Scheme 3 | 13.4 ± 0.17 | 1.8 ± 0.04 | 8.5 | 2.3 |
| Scheme | A Number of Fatal Cases on Day Post-Infection (n = 10) | Lethal Cases 1 | Lung Lesions 2 | ALE 3, Days | IP 4 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |||||
| The viral dose of 1 LD50 per mouse | |||||||||||||||
| Control | 0 | 0 | 1 | 3 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 70% | 40.0% | 9.8 | NA 5 |
| Scheme 1 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 20% | 6.3% | 13.2 | 71% |
| Scheme 2 | 0 | 0 | 1 | 2 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 60% | 60.0% | 10.7 | 14% |
| Scheme 3 | 0 | 0 | 1 | 1 | 3 | 1 | 1 | 0 | 0 | 0 | 0 | 70% | ND 6 | 9.8 | 0% |
| The viral dose of 10 LD50 per mouse | |||||||||||||||
| Control | 1 | 1 | 5 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 100% | 75.0% | 6.2 | NA |
| Scheme 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 40% | 25.0% | 11.5 | 60% |
| Scheme 2 | 0 | 1 | 0 | 3 | 2 | 2 | 1 | 1 | 0 | 0 | 0 | 100% | 65.0% | 8.1 | 0% |
| Scheme 3 | 0 | 0 | 2 | 2 | 2 | 0 | 0 | 2 | 0 | 0 | 0 | 80% | ND | 9.2 | 20% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiseleva, I.V.; Farroukh, M.A.; Skomorokhova, E.A.; Rekstin, A.R.; Bazhenova, E.A.; Magazenkova, D.N.; Orlov, I.A.; Rudenko, L.G.; Broggini, M.; Puchkova, L.V. Anti-Influenza Effect of Nanosilver in a Mouse Model. Vaccines 2020, 8, 679. https://doi.org/10.3390/vaccines8040679
Kiseleva IV, Farroukh MA, Skomorokhova EA, Rekstin AR, Bazhenova EA, Magazenkova DN, Orlov IA, Rudenko LG, Broggini M, Puchkova LV. Anti-Influenza Effect of Nanosilver in a Mouse Model. Vaccines. 2020; 8(4):679. https://doi.org/10.3390/vaccines8040679
Chicago/Turabian StyleKiseleva, Irina V., Mohammad Al Farroukh, Ekaterina A. Skomorokhova, Andrei R. Rekstin, Ekaterina A. Bazhenova, Daria N. Magazenkova, Iurii A. Orlov, Larisa G. Rudenko, Massimo Broggini, and Ludmila V. Puchkova. 2020. "Anti-Influenza Effect of Nanosilver in a Mouse Model" Vaccines 8, no. 4: 679. https://doi.org/10.3390/vaccines8040679
APA StyleKiseleva, I. V., Farroukh, M. A., Skomorokhova, E. A., Rekstin, A. R., Bazhenova, E. A., Magazenkova, D. N., Orlov, I. A., Rudenko, L. G., Broggini, M., & Puchkova, L. V. (2020). Anti-Influenza Effect of Nanosilver in a Mouse Model. Vaccines, 8(4), 679. https://doi.org/10.3390/vaccines8040679