Priming with Recombinant BCG Expressing HTI Enhances the Magnitude and Breadth of the T-Cell Immune Responses Elicited by MVA.HTI in BALB/c Mice

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
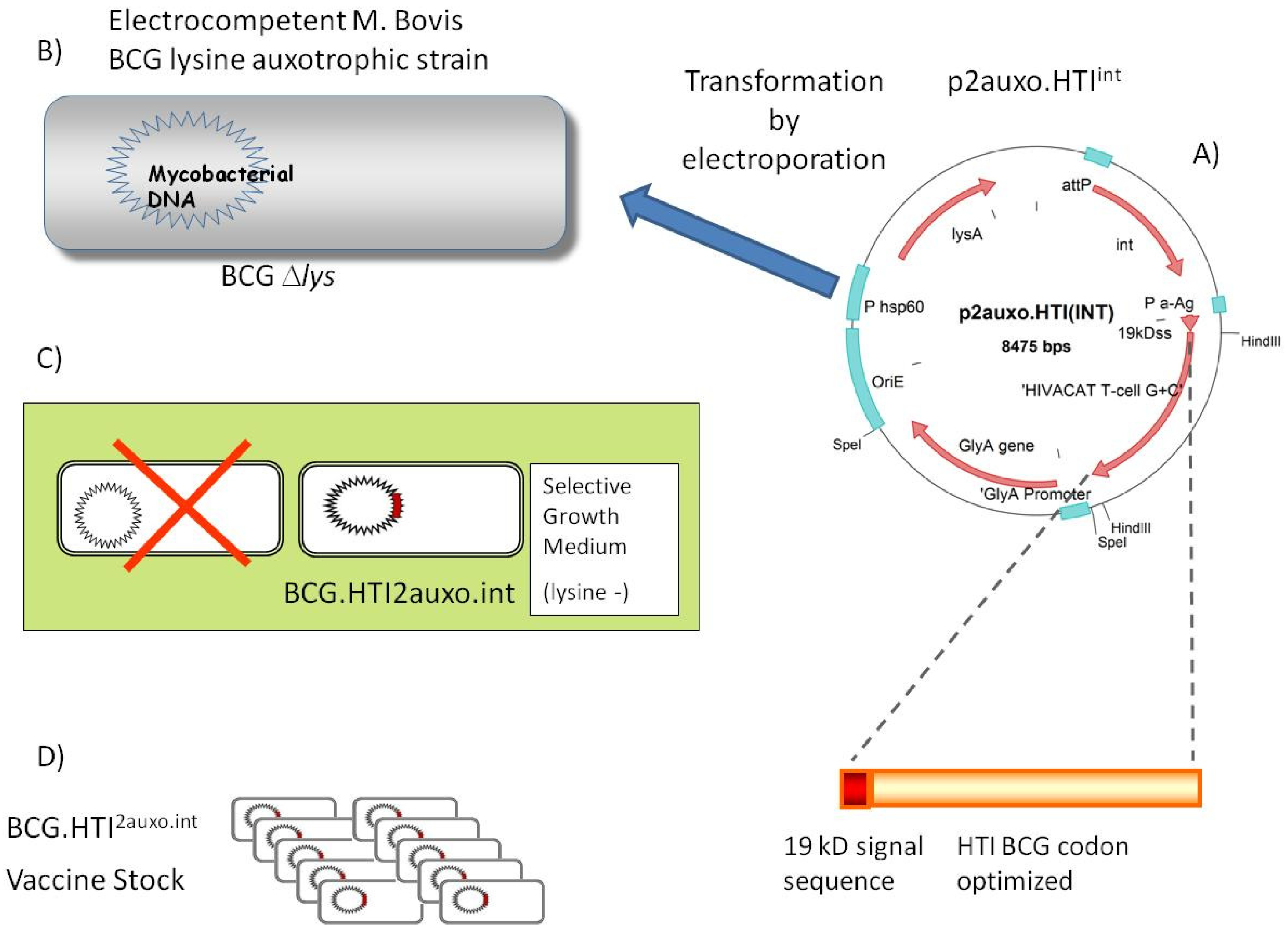
2.1. Construction of the BCG.HTI2auxo.int Strain Using an Antibiotic-Free Plasmid Selection System
2.2. Bacterial Cultures
2.3. Viral Strains
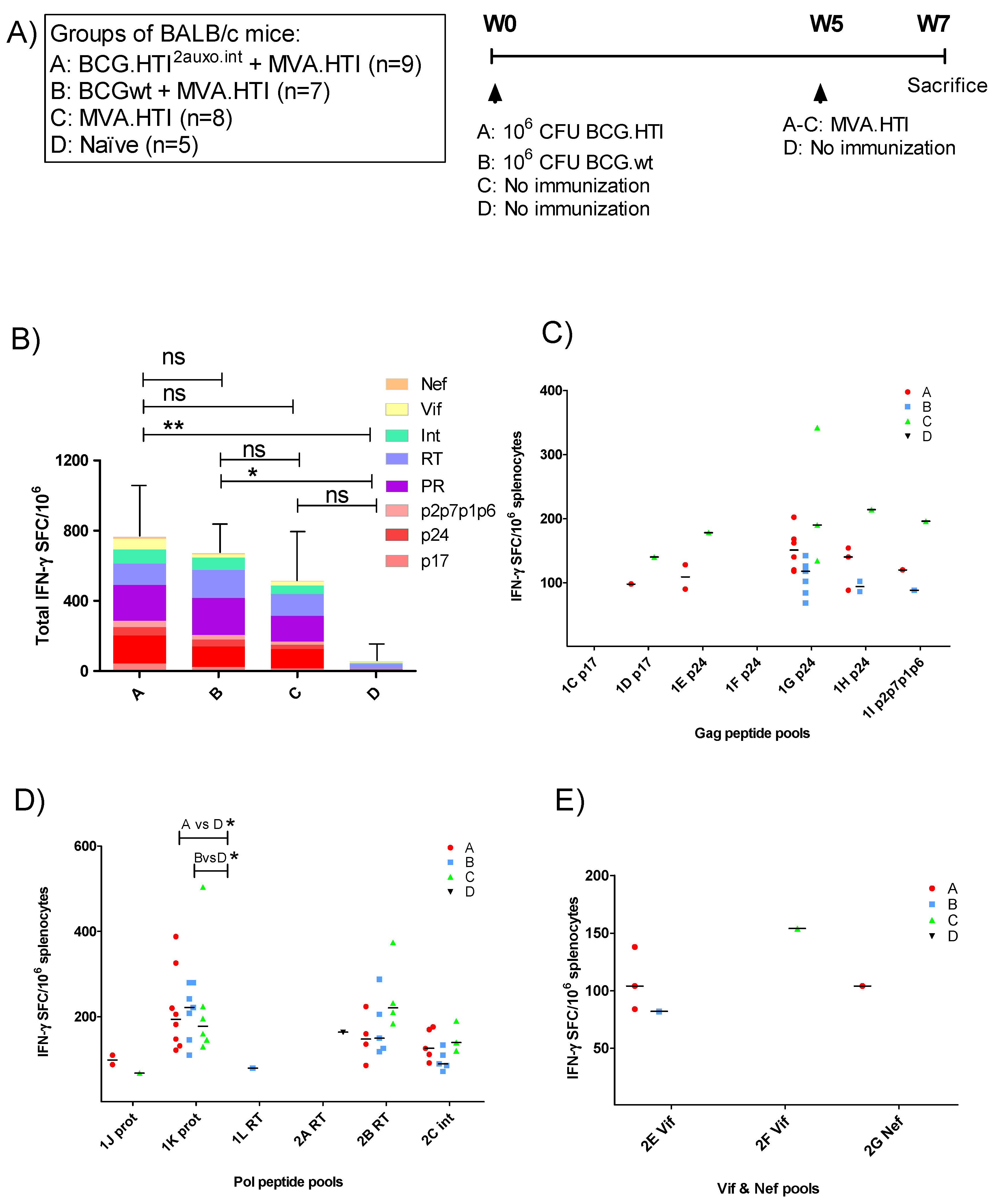
2.4. Immunization of Mice and Isolation of Splenocytes
2.5. IFN-γ ELISpot Analysis
2.6. Statistical Analysis
2.7. Ethics Statement
3. Results
3.1. Construction of the BCG.HTI2auxo.int Vaccine Strain
3.2. The BCG.HTI2auxo.int Prime-MVA.HTI Boost Regimen Elicits HIV-1-Specific T-Cell Responses
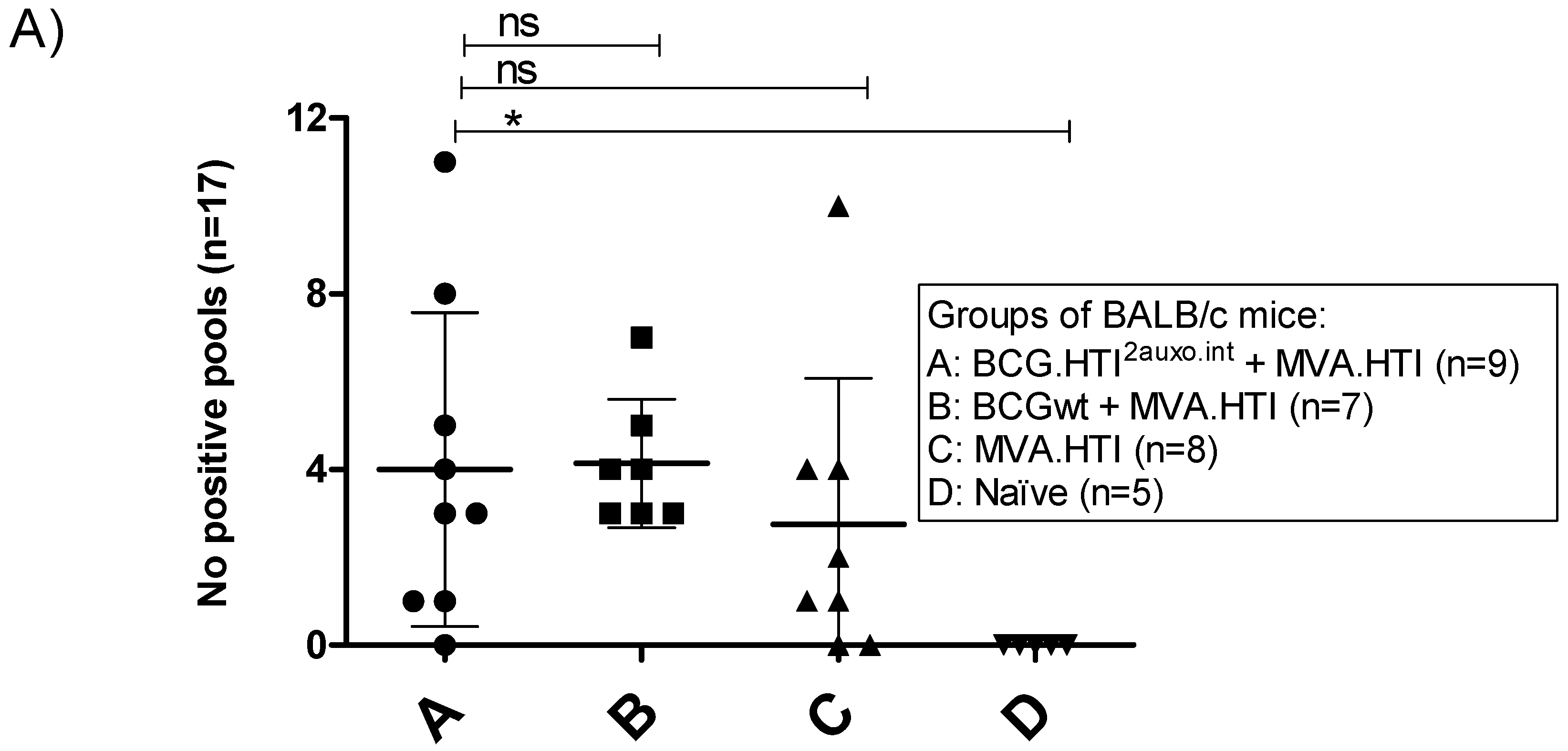
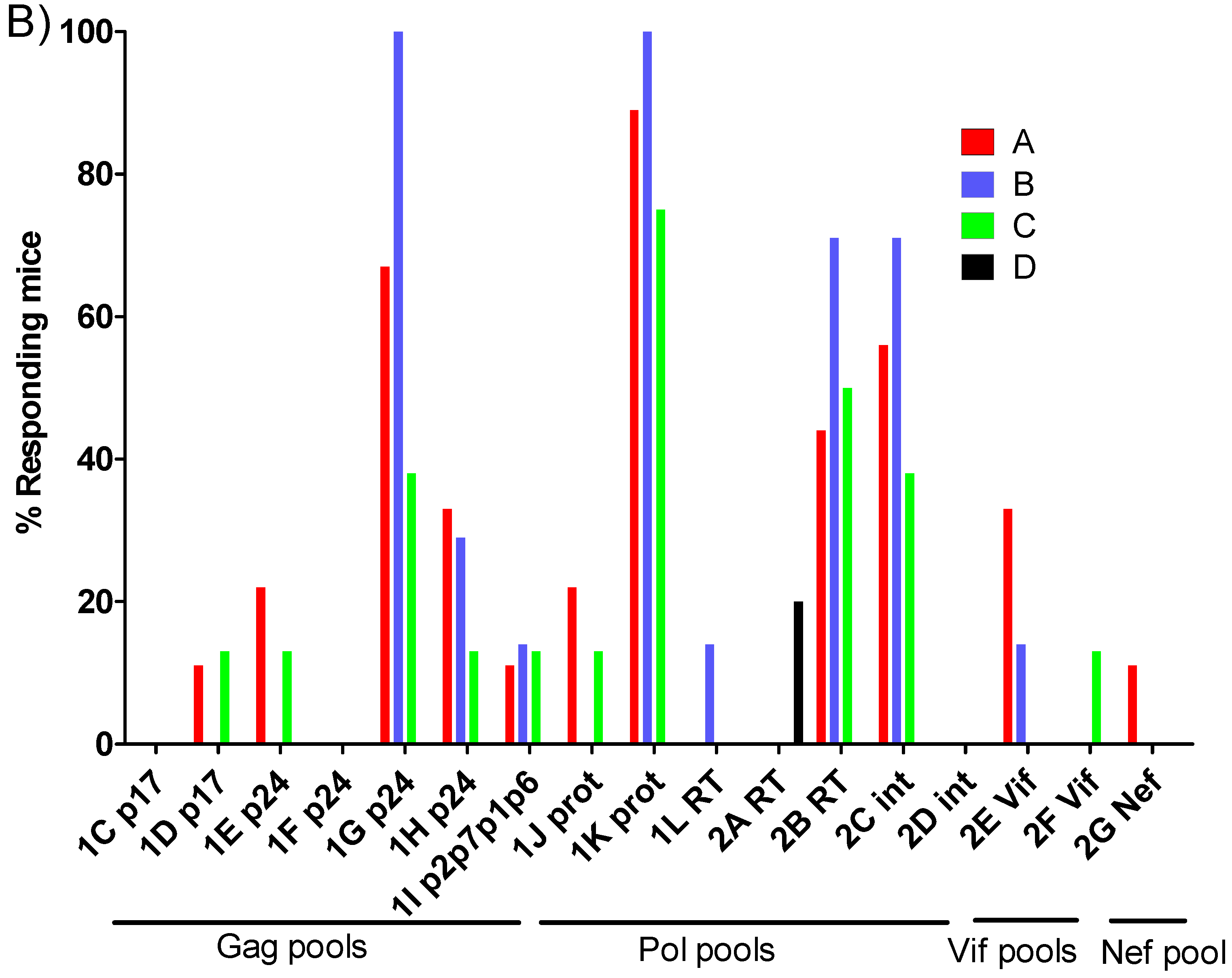
3.3. Breadth of T-Cell Immune Responses in BCG.HT2auxo.int + MVA.HTI Immunized BALB/c Mice
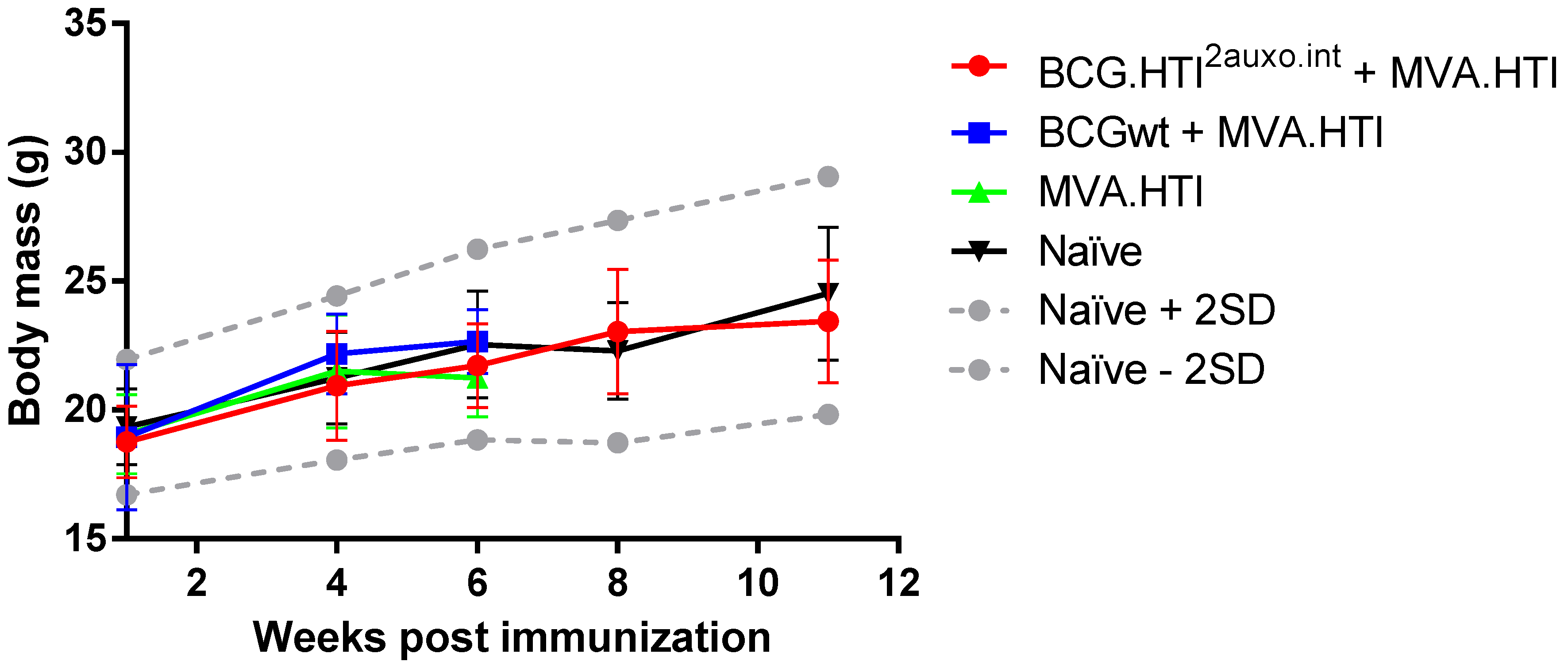
3.4. The BCG.HTI2auxo.int + MVA.HTI Prime-Boost Regimen Is Safe in BALB/c Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Global HIV & AIDS Statistics—2020 Fact Sheet|UNAIDS. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 2 October 2020).
- World Health Organization Global HIV & AIDS Statistics—2018 fact sheet|UNAIDS. Available online: https://www.unaids.org/sites/default/files/media_asset/UNAIDS_FactSheet_en.pdf (accessed on 6 September 2019).
- Rowland-Jones, S.L.; Dong, T.; Fowke, K.R.; Kimani, J.; Krausa, P.; Newell, H.; Blanchard, T.; Ariyoshi, K.; Oyugi, J.; Ngugi, E.; et al. Cytotoxic T cell responses to multiple conserved HIV epitopes in HIV-resistant prostitutes in Nairobi. J. Clin. Invest. 1998, 102, 1758–1765. [Google Scholar] [CrossRef]
- McBrien, J.B.; Kumar, N.A.; Silvestri, G. Mechanisms of CD8+ T cell-mediated suppression of HIV/SIV replication. Eur. J. Immunol. 2018, 48, 898–914. [Google Scholar] [CrossRef]
- Stover, C.K.; Bansal, G.P.; Hanson, M.S.; Burlein, J.E.; Palaszynski, S.R.; Young, J.F.; Koenig, S.; Young, D.B.; Sadziene, A.; Barbour, A.G. Protective immunity elicited by recombinant bacille Calmette-Guerin (BCG) expressing outer surface protein A (OspA) lipoprotein: A candidate Lyme disease vaccine. J. Exp. Med. 1993, 178, 197–209. [Google Scholar] [CrossRef]
- Aldovini, A.; Young, R.A. Humoral and cell-mediated immune responses to live recombinant BCG-HIV vaccines. Nature 1991, 351, 479–482. [Google Scholar] [CrossRef]
- Chapman, R.; Chege, G.; Shephard, E.; Stutz, H.; Williamson, A.-L. Recombinant Mycobacterium bovis BCG as an HIV vaccine vector. Curr. HIV Res. 2010, 8, 282–298. [Google Scholar] [CrossRef]
- Sixsmith, J.D.; Panas, M.W.; Lee, S.; Gillard, G.O.; White, K.; Lifton, M.A.; Balachandran, H.; Mach, L.; Miller, J.P.; Lavine, C.; et al. Recombinant Mycobacterium bovis Bacillus Calmette-Guerin Vectors Prime for Strong Cellular Responses to Simian Immunodeficiency Virus Gag in Rhesus Macaques. Clin. Vaccine Immunol. 2014, 21, 1385–1395. [Google Scholar] [CrossRef]
- Gheorghiu, M.; Lagrange, P.H.; Fillastre, C. The stability and immunogenicity of a dispersed-grown freeze-dried Pasteur BCG vaccine. J. Biol. Stand. 1988, 16, 15–26. [Google Scholar] [CrossRef]
- Matsuo, K.; Yasutomi, Y. Mycobacterium bovis Bacille Calmette-Guérin as a Vaccine Vector for Global Infectious Disease Control. Tuberc. Res. Treat. 2011, 2011, 574591. [Google Scholar] [CrossRef][Green Version]
- Joseph, J.; Saubi, N.; Pezzat, E.; Gatell, J.M. Progress towards an HIV vaccine based on recombinant bacillus Calmette-Guérin: Failures and challenges. Expert Rev. Vaccines 2006, 5, 827–838. [Google Scholar] [CrossRef]
- Melancon-Kaplan, J.; Hunter, S.W.; McNeil, M.; Stewart, C.; Modlin, R.L.; Rea, T.H.; Convit, J.; Salgame, P.; Mehra, V.; Bloom, B.R. Immunological significance of Mycobacterium leprae cell walls. Proc. Natl. Acad. Sci. USA 1988, 85, 1917–1921. [Google Scholar] [CrossRef]
- Oiso, R.; Fujiwara, N.; Yamagami, H.; Maeda, S.; Matsumoto, S.; Nakamura, S.; Oshitani, N.; Matsumoto, T.; Arakawa, T.; Kobayashi, K. Mycobacterial trehalose 6,6’-dimycolate preferentially induces type 1 helper T cell responses through signal transducer and activator of transcription 4 protein. Microb. Pathog. 2005, 39, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Naka, T.; Doi, T.; Yano, I. Direct molecular mass determination of trehalose monomycolate from 11 species of mycobacteria by MALDI-TOF mass spectrometry. Microbiology 2005, 151, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Brightbill, H.D.; Libraty, D.H.; Krutzik, S.R.; Yang, R.B.; Belisle, J.T.; Bleharski, J.R.; Maitland, M.; Norgard, M.V.; Plevy, S.E.; Smale, S.T.; et al. Host defense mechanisms triggered by microbial lipoproteins through toll-like receptors. Science 1999, 285, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.who.int/wer/2004/en/wer7904.pdf?ua=1 (accessed on 2 October 2020).
- Kaufmann, S.H.E. Vaccination Against Tuberculosis: Revamping BCG by Molecular Genetics Guided by Immunology. Front. Immunol. 2020, 11, 316. [Google Scholar] [CrossRef] [PubMed]
- Abarca, K.; Rey-Jurado, E.; Muñoz-Durango, N.; Vázquez, Y.; Soto, J.A.; Gálvez, N.M.S.; Valdés-Ferrada, J.; Iturriaga, C.; Urzúa, M.; Borzutzky, A.; et al. Safety and immunogenicity evaluation of recombinant BCG vaccine against respiratory syncytial virus in a randomized, double-blind, placebo-controlled phase I clinical trial. EClinicalMedicine 2020, 27, 100517. [Google Scholar] [CrossRef]
- World Health Organization. BCG vaccine: WHO position paper, February 2018—Recommendations. Vaccine 2018, 36, 3408–3410. [Google Scholar] [CrossRef]
- Im, E.-J.; Saubi, N.; Virgili, G.; Sander, C.; Teoh, D.; Gatell, J.M.; McShane, H.; Joseph, J.; Hanke, T. Vaccine platform for prevention of tuberculosis and mother-to-child transmission of human immunodeficiency virus type 1 through breastfeeding. J. Virol. 2007, 81, 9408–9418. [Google Scholar] [CrossRef]
- Hanke, T.; McMichael, A.J. Design and construction of an experimental HIV-1 vaccine for a year-2000 clinical trial in Kenya. Nat. Med. 2000, 6, 951–955. [Google Scholar] [CrossRef]
- Saubi, N.; Im, E.-J.; Fernández-Lloris, R.; Gil, O.; Cardona, P.-J.; Gatell, J.M.; Hanke, T.; Joseph, J. Newborn Mice Vaccination with BCG.HIVA222 + MVA.HIVA Enhances HIV-1-Specific Immune Responses: Influence of Age and Immunization Routes. Clin. Dev. Immunol. 2011, 2011, 1–11. [Google Scholar] [CrossRef]
- Saubi, N.; Gea-Mallorquí, E.; Ferrer, P.; Hurtado, C.; Sánchez-Úbeda, S.; Eto, Y.; Gatell, J.M.; Hanke, T.; Joseph, J. Engineering new mycobacterial vaccine design for HIV-TB pediatric vaccine vectored by lysine auxotroph of BCG. Mol. Ther. Methods Clin. Dev. 2014, 1, 14017. [Google Scholar] [CrossRef]
- Mahant, A.; Saubi, N.; Eto, Y.; Guitart, N.; Gatell, J.M.; Hanke, T.; Joseph, J. Preclinical development of BCG.HIVA 2auxo.int, harboring an integrative expression vector, for a HIV-TB Pediatric vaccine. Enhancement of stability and specific HIV-1 T-cell immunity. Hum. Vaccin. Immunother. 2017, 1–13. [Google Scholar] [CrossRef]
- Arbues, A.; Aguilo, J.I.; Gonzalo-Asensio, J.; Marinova, D.; Uranga, S.; Puentes, E.; Fernandez, C.; Parra, A.; Cardona, P.J.; Vilaplana, C.; et al. Construction, characterization and preclinical evaluation of MTBVAC, the first live-attenuated M. tuberculosis-based vaccine to enter clinical trials. Vaccine 2013, 31, 4867–4873. [Google Scholar] [CrossRef]
- Broset, E.; Saubi, N.; Guitart, N.; Aguilo, N.; Uranga, S.; Kilpeläinen, A.; Eto, Y.; Hanke, T.; Gonzalo-Asensio, J.; Martín, C.; et al. MTBVAC-Based TB-HIV Vaccine Is Safe, Elicits HIV-T Cell Responses, and Protects against Mycobacterium tuberculosis in Mice. Mol. Ther. Methods Clin. Dev. 2019, 13, 253–264. [Google Scholar] [CrossRef]
- Ondondo, B.; Murakoshi, H.; Clutton, G.; Abdul-Jawad, S.; Wee, E.G.-T.; Gatanaga, H.; Oka, S.; McMichael, A.J.; Takiguchi, M.; Korber, B.; et al. Novel Conserved-region T-cell Mosaic Vaccine with High Global HIV-1 Coverage Is Recognized by Protective Responses in Untreated Infection. Mol. Ther. 2016, 24, 832–842. [Google Scholar] [CrossRef]
- Kilpeläinen, A.; Saubi, N.; Guitart, N.; Moyo, N.; Wee, E.G.; Ravi, K.; Hanke, T.; Joseph, J. Priming with Recombinant BCG Expressing Novel HIV-1 Conserved Mosaic Immunogens and Boosting with Recombinant ChAdOx1 Is Safe, Stable, and Elicits HIV-1-Specific T-Cell Responses in BALB/c Mice. Front. Immunol. 2019, 10, 923. [Google Scholar] [CrossRef]
- Létourneau, S.; Im, E.-J.; Mashishi, T.; Brereton, C.; Bridgeman, A.; Yang, H.; Dorrell, L.; Dong, T.; Korber, B.; McMichael, A.J.; et al. Design and Pre-Clinical Evaluation of a Universal HIV-1 Vaccine. PLoS ONE 2007, 2, e984. [Google Scholar] [CrossRef]
- Mothe, B.; Hu, X.; Llano, A.; Rosati, M.; Olvera, A.; Kulkarni, V.; Valentin, A.; Alicea, C.; Pilkington, G.R.; Sardesai, N.Y.; et al. A human immune data-informed vaccine concept elicits strong and broad T-cell specificities associated with HIV-1 control in mice and macaques. J. Transl. Med. 2015, 13, 60. [Google Scholar] [CrossRef]
- Kilpeläinen, A.; Saubi, N.; Guitart, N.; Olvera, A.; Hanke, T.; Brander, C.; Joseph, J. Recombinant BCG Expressing HTI Prime and Recombinant ChAdOx1 Boost Is Safe and Elicits HIV-1-Specific T-Cell Responses in BALB/c Mice. Vaccines 2019, 7, 78. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Matsuo, K.; Hamatake, M.; Hamano, T.; Ohsu, T.; Matsumoto, S.; Yamada, T.; Yamazaki, S.; Hasegawa, A.; Yamamoto, N.; et al. Mycobacterial codon optimization enhances antigen expression and virus-specific immune responses in recombinant Mycobacterium bovis bacille Calmette-Guérin expressing human immunodeficiency virus type 1 Gag. J. Virol. 2005, 79, 8716–8723. [Google Scholar] [CrossRef]
- Pavelka, M.S.; Jacobs, W.R. Comparison of the construction of unmarked deletion mutations in Mycobacterium smegmatis, Mycobacterium bovis bacillus Calmette-Guerin, and Mycobacterium tuberculosis H37RV by allelic exchange. J. Bacteriol. 1999, 181, 4780–4789. [Google Scholar] [CrossRef]
- Lee, M.H.; Pascopella, L.; Jacobs, W.R.; Hatfull, G.F. Site-specific integration of mycobacteriophage L5: Integration-proficient vectors for Mycobacterium smegmatis, Mycobacterium tuberculosis, and bacille Calmette-Guérin. Proc. Natl. Acad. Sci. USA 1991, 88, 3111–3115. [Google Scholar] [CrossRef]
- Hansen, S.G.; Wu, H.L.; Burwitz, B.J.; Hughes, C.M.; Hammond, K.B.; Ventura, A.B.; Reed, J.S.; Gilbride, R.M.; Ainslie, E.; Morrow, D.W.; et al. Broadly targeted CD8+ T cell responses restricted by major histocompatibility complex E. Science 2016, 351, 714–720. [Google Scholar] [CrossRef]
- Hannoun, Z.; Lin, Z.; Brackenridge, S.; Kuse, N.; Akahoshi, T.; Borthwick, N.; McMichael, A.; Murakoshi, H.; Takiguchi, M.; Hanke, T. Identification of novel HIV-1-derived HLA-E-binding peptides. Immunol. Lett. 2018, 202, 65–72. [Google Scholar] [CrossRef]
- Joosten, S.A.; Van Meijgaarden, K.E.; Van Weeren, P.C.; Kazi, F.; Geluk, A.; Savage, N.D.L.; Drijfhout, J.W.; Flower, D.R.; Hanekom, W.A.; Klein, M.R.; et al. Mycobacterium tuberculosis peptides presented by HLA-E molecules are targets for human CD8+ T-cells with cytotoxic as well as regulatory activity. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef]
- Gallinaro, A.; Borghi, M.; Pirillo, M.F.; Cecchetti, S.; Bona, R.; Canitano, A.; Michelini, Z.; Di Virgilio, A.; Olvera, A.; Brander, C.; et al. Development and Preclinical Evaluation of an Integrase Defective Lentiviral Vector Vaccine Expressing the HIVACAT T Cell Immunogen in Mice. Mol. Ther. Methods Clin. Dev. 2020, 17, 418–428. [Google Scholar] [CrossRef]
- Rosario, M.; Hopkins, R.; Fulkerson, J.; Borthwick, N.; Quigley, M.F.; Joseph, J.; Douek, D.C.; Greenaway, H.Y.; Venturi, V.; Gostick, E.; et al. Novel Recombinant Mycobacterium bovis BCG, Ovine Atadenovirus, and Modified Vaccinia Virus Ankara Vaccines Combine To Induce Robust Human Immunodeficiency Virus-Specific CD4 and CD8 T-Cell Responses in Rhesus Macaques. J. Virol. 2010, 84, 5898–5908. [Google Scholar] [CrossRef]
- Moyo, N.; Borthwick, N.J.; Wee, E.G.; Capucci, S.; Crook, A.; Dorrell, L.; Hanke, T. Long-term follow up of human T-cell responses to conserved HIV-1 regions elicited by DNA/simian adenovirus/MVA vaccine regimens. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Borthwick, N.; Ahmed, T.; Ondondo, B.; Hayes, P.; Rose, A.; Ebrahimsa, U.; Hayton, E.-J.; Black, A.; Bridgeman, A.; Rosario, M.; et al. Vaccine-elicited human T cells recognizing conserved protein regions inhibit HIV-1. Mol. Ther. 2014, 22, 464–475. [Google Scholar] [CrossRef]
- Ondondo, B.; Abdul-Jawad, S.; Bridgeman, A.; Hanke, T. Characterization of T-cell responses to conserved regions of the HIV-1 proteome in BALB/c mice. Clin. Vaccine Immunol. 2014, 21, 1565–1572. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saubi, N.; Kilpeläinen, A.; Eto, Y.; Chen, C.-W.; Olvera, À.; Hanke, T.; Brander, C.; Joseph-Munné, J. Priming with Recombinant BCG Expressing HTI Enhances the Magnitude and Breadth of the T-Cell Immune Responses Elicited by MVA.HTI in BALB/c Mice. Vaccines 2020, 8, 678. https://doi.org/10.3390/vaccines8040678
Saubi N, Kilpeläinen A, Eto Y, Chen C-W, Olvera À, Hanke T, Brander C, Joseph-Munné J. Priming with Recombinant BCG Expressing HTI Enhances the Magnitude and Breadth of the T-Cell Immune Responses Elicited by MVA.HTI in BALB/c Mice. Vaccines. 2020; 8(4):678. https://doi.org/10.3390/vaccines8040678
Chicago/Turabian StyleSaubi, Narcís, Athina Kilpeläinen, Yoshiki Eto, Chun-Wei Chen, Àlex Olvera, Tomáš Hanke, Christian Brander, and Joan Joseph-Munné. 2020. "Priming with Recombinant BCG Expressing HTI Enhances the Magnitude and Breadth of the T-Cell Immune Responses Elicited by MVA.HTI in BALB/c Mice" Vaccines 8, no. 4: 678. https://doi.org/10.3390/vaccines8040678
APA StyleSaubi, N., Kilpeläinen, A., Eto, Y., Chen, C.-W., Olvera, À., Hanke, T., Brander, C., & Joseph-Munné, J. (2020). Priming with Recombinant BCG Expressing HTI Enhances the Magnitude and Breadth of the T-Cell Immune Responses Elicited by MVA.HTI in BALB/c Mice. Vaccines, 8(4), 678. https://doi.org/10.3390/vaccines8040678