Association of Porcine Swine Leukocyte Antigen (SLA) Haplotypes with B- and T-Cell Immune Response to Foot-and-Mouth Disease Virus (FMDV) Peptides

 , ,
, ,  , , , ,
, , , , 
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Peptide Vaccines
2.2. Pigs and Experimental Design
2.3. SLA Typing by PCR-SSP
2.4. Data Preparation and Statistical Analyses
2.5. Ethics Statement
3. Results
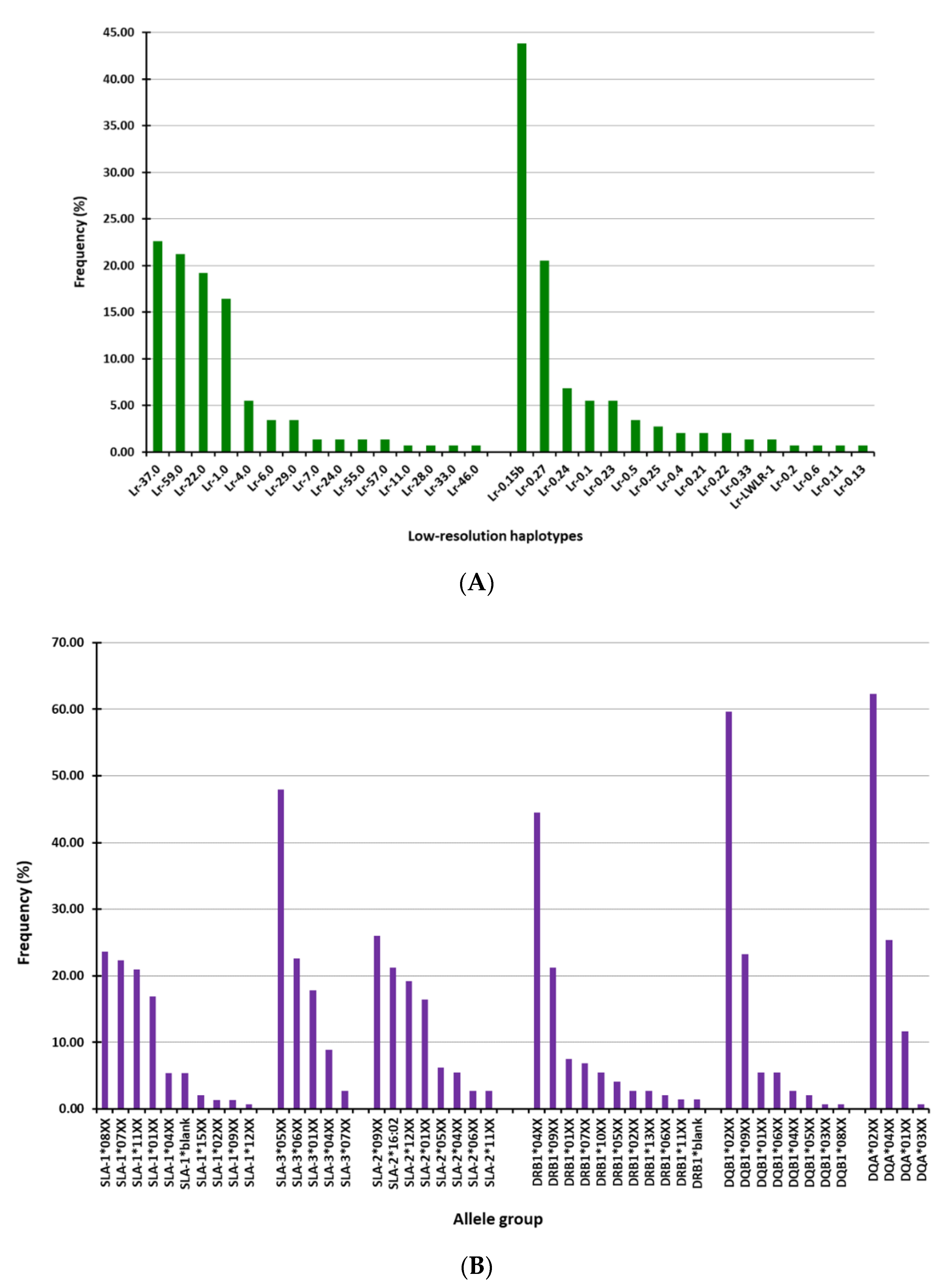
3.1. SLA-I and SLA-II Allele-Groups and Haplotypes Identified in the Studied Pigs
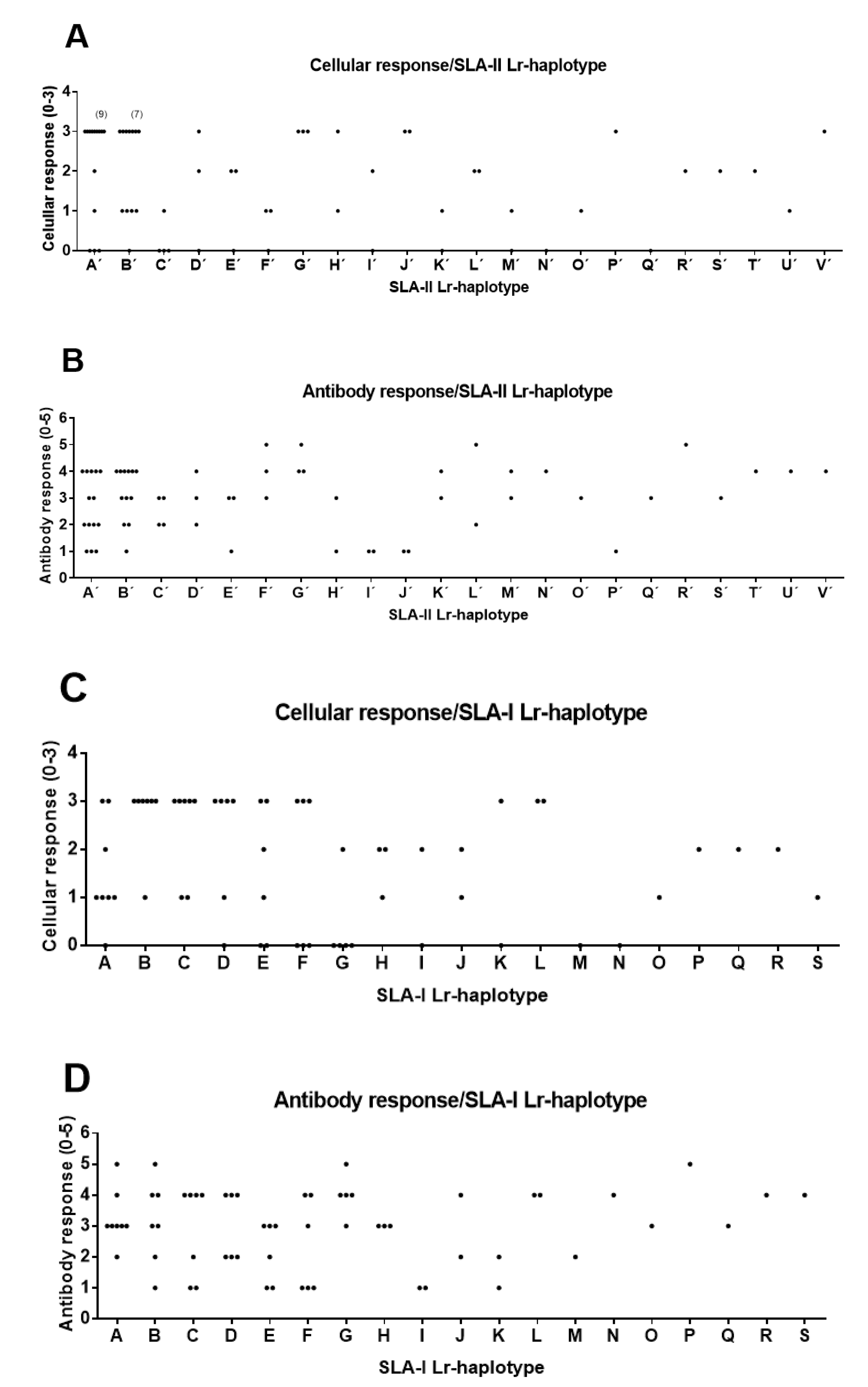
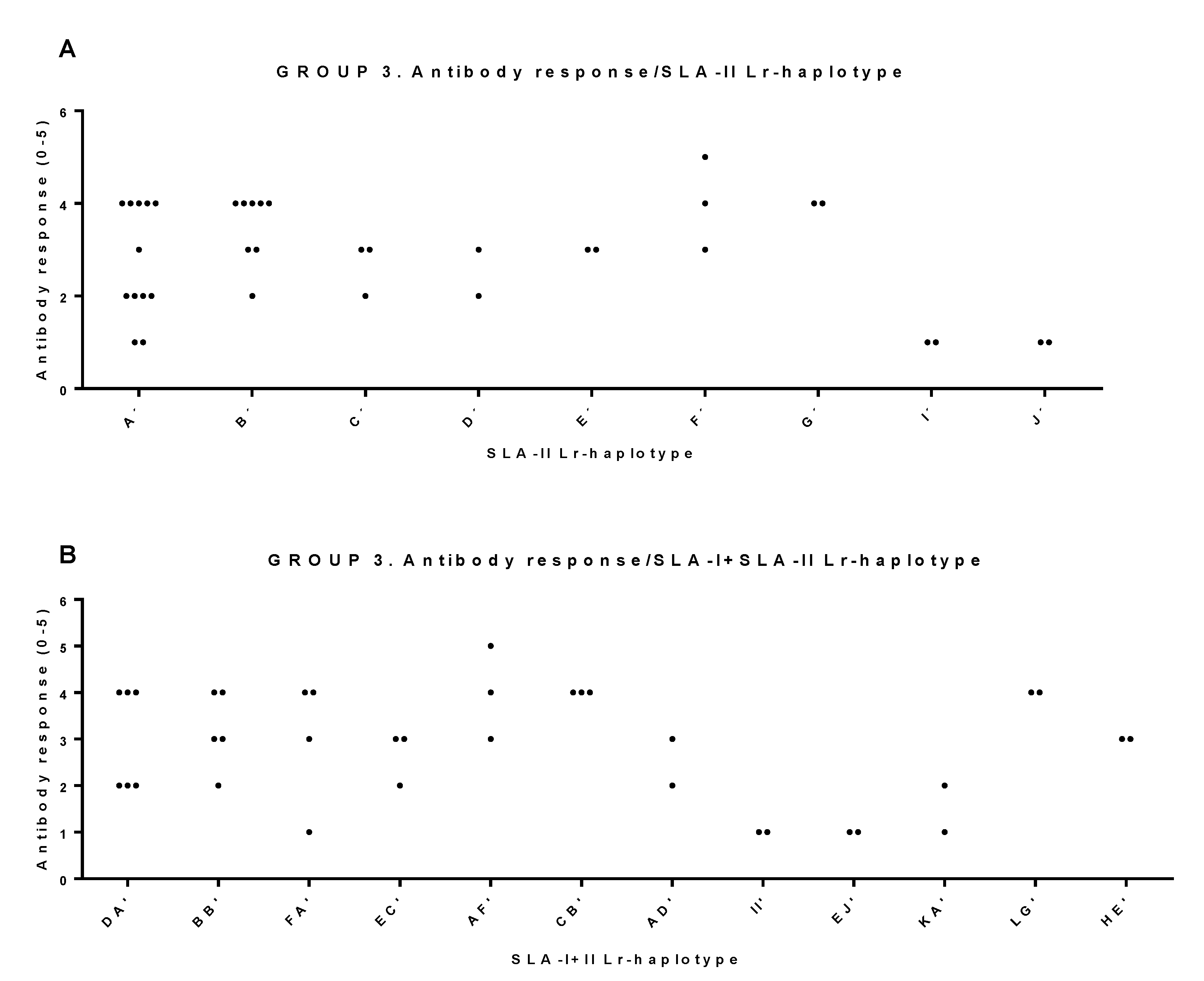
3.2. Correlations between SLA-I and SLA-II Haplotypes and Immune Response to Dendrimer Peptides
3.3. Association of SLA-I and SLA-II Haplotypes with High Immune Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blanco, E.; Andreu, D.; Sobrino, F. Advantages, challenges and future of peptide vacines. In Foot-and-Mouth Disease Virus. Current Research and Emerging Trends; Sobrino, F., Domingo, E., Eds.; Caister Academic Press: Norfolk, UK, 2017; pp. 317–331. [Google Scholar]
- Garcia-Briones, M.M.; Russell, G.C.; Oliver, R.A.; Tami, C.; Taboga, O.; Carrillo, E.; Palma, E.L.; Sobrino, F.; Glass, E.J. Association of bovine DRB3 alleles with immune response to FMDV peptides and protection against viral challenge. Vaccine 2000, 19, 1167–1171. [Google Scholar] [CrossRef]
- Kulski, J.K.; Shiina, T.; Dijkstra, J.M. Genomic Diversity of the Major Histocompatibility Complex in Health and Disease. Cells 2019, 8, 1270. [Google Scholar] [CrossRef]
- Grubman, M.J.; Baxt, B. Foot-and-mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef]
- Robinson, L.; Knight-Jones, T.J.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 7—Pathogenesis and Molecular Biology. Transbound. Emerg. Dis. 2016, 63, S63–S71. [Google Scholar] [CrossRef]
- Knight-Jones, T.J.; Robinson, L.; Charleston, B.; Rodriguez, L.L.; Gay, C.G.; Sumption, K.J.; Vosloo, W. Global Foot-and-Mouth Disease Research Update and Gap Analysis: 1—Overview of Global Status and Research Needs. Transbound. Emerg. Dis. 2016, 63, S3–S13. [Google Scholar] [CrossRef]
- Cao, Y.; Lu, Z.; Liu, Z. Foot-and-mouth disease vaccines: Progress and problems. Expert Rev. Vaccines 2016, 15, 783–789. [Google Scholar] [CrossRef]
- Kleid, D.G.; Yansura, D.; Small, B.; Dowbenko, D.; Moore, D.M.; Grubman, M.J.; McKercher, P.D.; Morgan, D.O.; Robertson, B.H.; Bachrach, H.L. Cloned viral protein vaccine for foot-and-mouth disease: Responses in cattle and swine. Science 1981, 214, 1125–1129. [Google Scholar] [CrossRef]
- Bittle, J.L.; Houghten, R.A.; Alexander, H.; Shinnick, T.M.; Sutcliffe, J.G.; Lerner, R.A.; Rowlands, D.J.; Brown, F. Protection against foot-and-mouth disease by immunization with a chemically synthesized peptide predicted from the viral nucleotide sequence. Nature 1982, 298, 30–33. [Google Scholar] [CrossRef]
- Strohmaier, K.; Franze, R.; Adam, K.H. Location and characterization of the antigenic portion of the FMDV immunizing protein. J. Gen. Virol. 1982, 59, 295–306. [Google Scholar] [CrossRef]
- DiMarchi, R.; Brooke, G.; Gale, C.; Cracknell, V.; Doel, T.; Mowat, N. Protection of cattle against foot-and-mouth disease by a synthetic peptide. Science 1986, 232, 639–641. [Google Scholar] [CrossRef]
- Doel, T.R.; Gale, C.; Brooke, G.; DiMarchi, R. Immunization against foot-and-mouth disease with synthetic peptides representing the C-terminal region of VP1. J. Gen. Virol. 1988, 69, 2403–2406. [Google Scholar] [CrossRef] [PubMed]
- Collen, T.; Dimarchi, R.; Doel, T.R. A T cell epitope in VP1 of foot-and-mouth disease virus is immunodominant for vaccinated cattle. J. Immunol. 1991, 146, 749–755. [Google Scholar] [PubMed]
- Firbas, C.; Jilma, B.; Tauber, E.; Buerger, V.; Jelovcan, S.; Lingnau, K.; Buschle, M.; Frisch, J.; Klade, C.S. Immunogenicity and safety of a novel therapeutic hepatitis C virus (HCV) peptide vaccine: A randomized, placebo controlled trial for dose optimization in 128 healthy subjects. Vaccine 2006, 24, 4343–4353. [Google Scholar] [CrossRef] [PubMed]
- Kunwar, P.; Hawkins, N.; Dinges, W.L.; Liu, Y.; Gabriel, E.E.; Swan, D.A.; Stevens, C.E.; Maenza, J.; Collier, A.C.; Mullins, J.I.; et al. Superior control of HIV-1 replication by CD8+ T cells targeting conserved epitopes: Implications for HIV vaccine design. PLoS ONE 2013, 8, e64405. [Google Scholar] [CrossRef] [PubMed]
- Taboga, O.; Tami, C.; Carrillo, E.; Nunez, J.I.; Rodriguez, A.; Saiz, J.C.; Blanco, E.; Valero, M.L.; Roig, X.; Camarero, J.A.; et al. A large-scale evaluation of peptide vaccines against foot-and-mouth disease: Lack of solid protection in cattle and isolation of escape mutants. J. Virol. 1997, 71, 2606–2614. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.S.; Zhai, X.X.; Jiang, P.; Zhang, Q.; Gao, H.; Li, Z.B.; Han, Y.; Yang, J.; Zhang, Z.H. Identification of two novel foot-and-mouth disease virus cytotoxic T lymphocyte epitopes that can bind six SLA-I proteins. Gene 2018, 653, 91–101. [Google Scholar] [CrossRef]
- Pedersen, L.E.; Patch, J.R.; Kenney, M.; Glabman, R.A.; Nielsen, M.; Jungersen, G.; Buus, S.; Golde, W.T. Expanding specificity of class I restricted CD8(+) T cells for viral epitopes following multiple inoculations of swine with a human adenovirus vectored foot-and-mouth disease virus (FMDV) vaccine. Vet. Immunol. Immunopathol. 2016, 181, 59–67. [Google Scholar] [CrossRef]
- Ning, S.; Wang, Z.B.; Qi, P.; Xiao, J.; Wang, X.J. Crystallization of SLA-2*04:02:02 complexed with a CTL epitope derived from FMDV. Res. Vet. Sci. 2020, 128, 90–98. [Google Scholar] [CrossRef]
- Tam, J.P. Synthetic peptide vaccine design: Synthesis and properties of a high-density multiple antigenic peptide system. Proc. Natl. Acad. Sci. USA 1988, 85, 5409–5413. [Google Scholar] [CrossRef]
- Blanco, E.; Garcia-Briones, M.; Sanz-Parra, A.; Gomes, P.; De Oliveira, E.; Valero, M.L.; Andreu, D.; Ley, V.; Sobrino, F. Identification of T-cell epitopes in nonstructural proteins of foot-and-mouth disease virus. J. Virol. 2001, 75, 3164–3174. [Google Scholar] [CrossRef]
- Cubillos, C.; de la Torre, B.G.; Jakab, A.; Clementi, G.; Borras, E.; Barcena, J.; Andreu, D.; Sobrino, F.; Blanco, E. Enhanced mucosal immunoglobulin A response and solid protection against foot-and-mouth disease virus challenge induced by a novel dendrimeric peptide. J. Virol. 2008, 82, 7223–7230. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Briones, M.M.; Blanco, E.; Chiva, C.; Andreu, D.; Ley, V.; Sobrino, F. Immunogenicity and T cell recognition in swine of foot-and-mouth disease virus polymerase 3D. Virology 2004, 322, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Blanco, E.; Guerra, B.; de la Torre, B.G.; Defaus, S.; Dekker, A.; Andreu, D.; Sobrino, F. Full protection of swine against foot-and-mouth disease by a bivalent B-cell epitope dendrimer peptide. Antivir. Res. 2016, 129, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Cañas-Arranz, R.; Forner, M.; Defaus, S.; de Leon, P.; Bustos, M.J.; Torres, E.; Sobrino, F.; Andreu, D.; Blanco, E. A Single Dose of Dendrimer B2T Peptide Vaccine Partially Protects Pigs against Foot-and-Mouth Disease Virus Infection. Vaccines 2020, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Cañas-Arranz, R.; Forner, M.; Defaus, S.; Rodriguez-Pulido, M.; de Leon, P.; Torres, E.; Bustos, M.J.; Borrego, B.; Saiz, M.; Blanco, E.; et al. A bivalent B-cell epitope dendrimer peptide can confer long-lasting immunity in swine against foot-and-mouth disease. Transbound. Emerg. Dis. 2020, 67, 1614–1622. [Google Scholar] [CrossRef]
- Cañas-Arranz, R.; de León, P.; Forner, M.; Defaus, S.; Bustos, M.J.; Torres, E.; Andreu, D.; Blanco, E.; Sobrino, F. Immunogenicity of a dendrimer B2T peptide harboring a T-cell epitope from FMDV non-structural protein 3D. Front. Vet. Sci. 2020. [Google Scholar] [CrossRef]
- Defaus, S.; Forner, R.; Cañas-Arranz, R.; de León, P.; Bustos, M.J.; Rodríguez-Pulido, M.; Blanco, E.; Sobrino, F.; Andreu, D. Designing functionally versatile, highly immunogenic peptide-based multiepitopic vaccines against foot-and-mouth disease virus. Vaccines 2020, 8, 406. [Google Scholar] [CrossRef]
- Hammer, S.E.; Ho, C.S.; Ando, A.; Rogel-Gaillard, C.; Charles, M.; Tector, M.; Tector, A.J.; Lunney, J.K. Importance of the Major Histocompatibility Complex (Swine Leukocyte Antigen) in Swine Health and Biomedical Research. Annu. Rev. Anim. Biosci. 2020, 8, 171–198. [Google Scholar] [CrossRef]
- Lunney, J.K.; Ho, C.S.; Wysocki, M.; Smith, D.M. Molecular genetics of the swine major histocompatibility complex, the SLA complex. Dev. Comp. Immunol. 2009, 33, 362–374. [Google Scholar] [CrossRef]
- Monso, M.; de la Torre, B.G.; Blanco, E.; Moreno, N.; Andreu, D. Influence of conjugation chemistry and B epitope orientation on the immune response of branched peptide antigens. Bioconjug. Chem. 2013, 24, 578–585. [Google Scholar] [CrossRef]
- Gimsa, U.; Ho, C.S.; Hammer, S.E. Preferred SLA class I/class II haplotype combinations in German Landrace pigs. Immunogenetics 2017, 69, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Lunney, J.K.; Franzo-Romain, M.H.; Martens, G.W.; Lee, Y.J.; Lee, J.H.; Wysocki, M.; Rowland, R.R.; Smith, D.M. Molecular characterization of swine leucocyte antigen class I genes in outbred pig populations. Anim. Genet. 2009, 40, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.S.; Lunney, J.K.; Ando, A.; Rogel-Gaillard, C.; Lee, J.H.; Schook, L.B.; Smith, D.M. Nomenclature for factors of the SLA system, update 2008. Tissue Antigens 2009, 73, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Siegmund, D. Maximally selected chi square statistics. Biometrics 1982, 38, 1011–1016. [Google Scholar] [CrossRef]
- McHugh, M.L. The chi-square test of independence. Biochem. Med. 2013, 23, 143–149. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B2T | B2TT | B2T-TB2 | |
|---|---|---|---|
| General Structure 1 |  |  |  |
| B epitope | acetyl-PVTNVRGDLQVLAQKAARTC-amide | ||
| T-3A | AAIEFFEGMVHDSIK-amide | ||
| T-3D | IFSKHRGDTKMSAED-amide | ||
| Peptides | B2T-3A B2T-3D | B2TT-3A3D B2TT-3D3A | B2T-3A-3A-TB2 |
| MW 2 | 6742.8 Da 6770.8 Da | 8708.1 Da | 14,247.6 Da |
| HPLC 3 | 6.9 min (98%) 5.1 min (98%) | 6.2 min (98%) 6.0 min (97%) | 7.9 min (97%) |
| Experiment 1 | Peptide Vaccine | Amount (mg) | Boost | Animal Number |
|---|---|---|---|---|
| 1 [26,28] | B2T-3A | 2 | Yes | Sw01–04 |
| B2T-TB2 | 2 | Yes | Sw05–08 | |
| B2T-TB2 | 0.5 | Yes | Sw09–12 | |
| PBS | - | - | Sw13, 14 | |
| 2 [24] | B4T | 2 | Yes | Sw15, 16, 21, 23, 24, 30 |
| B2T-3A | 2 | Yes | Sw17–19, 26, 27, 29 | |
| B2T-THIO | 2 | Yes | Sw20, 22, 25, 28, 31 | |
| 3 [25] | B2T-3A | 2 | No | Sw32–36 |
| B2T-3A | 0.5 | No | Sw37–41 | |
| PBS | - | - | Sw42, 43 | |
| 4 [27] | B2T-3A | 2 | Yes | Sw44–47 |
| B2T-3D | 2 | Yes | Sw48–51 | |
| PBS | - | - | Sw52, 53 | |
| 5 [P. De León, manuscript in preparation] | B2T-3A | 2 | Yes | Sw54–57 |
| B2T-3D | 2 | Yes | Sw58–61 | |
| B2TT-3A3D | 2 | Yes | Sw62–65 | |
| B2TT-3D3A | 2 | Yes | Sw66–69 | |
| B2 | 2 | Yes | Sw70, 71 | |
| PBS | - | - | Sw72, 73 |
| Lr-Hp SLA-I 1 | Number 2 | Lr-Hp SLA-II 1 | Number 2 | ||
|---|---|---|---|---|---|
| A | 59.0/1.0 | 8 | A’ | 0.15b/0.15b | 14 |
| B | 22.0/59.0 | 7 | B’ | 0.15b/0.27 | 12 |
| C | 37.0/59.0 | 7 | C’ | 0.1/0.15b | 4 |
| D | 22.0/37.0 | 6 | D’ | 0.27/0.4 | 3 |
| E | 1.0/37.0 | 6 | E’ | 0.15b/0.24 | 3 |
| F | 1.0/22.0 | 6 | F’ | 0.1/0.27 | 3 |
| G | 59.0/4.0 | 5 | G’ | 0.15b/0.21 | 3 |
| H | 29.0/37.0 | 3 | H’ | 0.15b/0.25 | 2 |
| I | 57.0/37.0 | 2 | I’ | 0.24/0.33 | 2 |
| J | 6.0/4.0 | 2 | J’ | 0.24/0.24 | 2 |
| K | 37.0/37.0 | 2 | K’ | 0.23/0.5 | 2 |
| L | 7.0/22.0 | 2 | L’ | 0.15b/0.23 | 2 |
| M | 1.0/6.0 | 1 | M’ | 0.27/0.23 | 2 |
| N | 6.0/46.0 | 1 | N’ | 0.15b/0.5 | 1 |
| O | 33.0/28.0 | 1 | O’ | LWLR-1/0.27 | 1 |
| P | 6.0/24.0 | 1 | P’ | 0.24/0.27 | 1 |
| Q | 1.0/4.0 | 1 | Q’ | 0.27/0.5 | 1 |
| R | 22.0/24.0 | 1 | R’ | 0.23/0.22 | 1 |
| S | 22.0/11.0 | 1 | S’ | 0.1/0.22 | 1 |
| T’ | 0.27/0.22 | 1 | |||
| U’ | 0.23/0.11 | 1 | |||
| V’ | 0.13/0.27 | 1 |
| SLA Lr Haplotype | Cellular 1 | Antibody 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| SLA-I | SLA-II | SLA-I/II | SLA-I | SLA-II | SLA-I/II | |||
| Group 0 | χ2 | 0.053 | 0.015 | 0.099 | 0.065 | 0.102 | 0.042 | |
| LR χ2 | 0.046 | 0.038 | 0.265 | 0.251 | 0.634 | 0.914 | ||
| Cv | 0.59 | 0.66 | 0.81 | 0.58 | 0.61 | 0.81 | ||
| Group 1 | χ2 | 0.021 | 0.022 | 0.061 | 0.029 | 0.082 | 0.055 | |
| LR χ2 | 0.039 | 0.034 | 0.168 | 0.095 | 0.358 | 0.502 | ||
| Fisher’s | exact | 0.011 | 0.003 | 0.81 | 0.012 | 0.125 | 0.806 | |
| Cv | 0.64 | 0.66 | 0.81 | 0.62 | 0.61 | 0.8 | ||
| Group 2 | χ2 | 0.097 | 0.01 | 0.054 | 0.123 | 0.05 | 0.035 | |
| LR χ2 | 0.051 | 0.014 | 0.111 | 0.107 | 0.264 | 0.416 | ||
| Fisher’s | exact | 0.041 | 0.001 | 0.8 | 0.041 | 0.074 | 0.807 | |
| Cv | 0.6 | 0.67 | 0.8 | 0.58 | 0.62 | 0.8 | ||
| Group 3 | χ2 | 0.077 | 0.004 | 0.034 | 0.067 | 0.01 | 0.03 | |
| LR χ2 | 0.128 | 0.008 | 0.068 | 0.024 | 0.019 | 0.022 | ||
| Fisher’s | exact | 0.148 | 0.001 | 0.029 | 0.044 | 0.011 | 0.02 | |
| Cv | 0.64 | 0.67 | 0.71 | 0.72 | 0.65 | 0.72 | ||
| SLA-I, HIGH T-Cell Response 1 | SLA-I, HIGH Antibody Response 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| SLA-1 | SLA-3 | SLA-2 | Lr-Hp 4 | Haplotype 5 | SLA-1 | SLA-3 | SLA-2 | Lr-Hp 4 | Haplotype 5 |
| 08XX | 07XX | 05XX | 7.0 (2) | L | 01XX | 01XX | 01XX | 1.0 (4) | A, F |
| 01XX | 01XX | 01XX | 1.0 (7) | A, E, F | 04XX | 04XX | 04XX | 4.0 (5) | G |
| 07XX | 05XX | 09XX | 37.0 (12) | C, D, E, K | 07XX | 05XX | 09XX | 37.0 (7) | C, D |
| 11XX | 05XX | 16:02 | 59.0 (13) | A, B, C | 08XX | 06XX | 12XX | 22.0 (12) | B, D, F, L, R, S |
| 08XX | 06XX | 12XX | 22.0 (15) | B, D, F, L | 11XX | 05XX | 16:02 | 59.0 (12) | A, B, C, G |
| SLA-I, LOW Antibody Response 3 | SLA-II, HIGH T-Cell Response 1 | ||||||||
| SLA-1 | SLA-3 | SLA-2 | Lr-Hp 4 | Haplotype 5 | DRB1 | DQB1 | DQA | Lr-Hp 4 | Haplotype 5 |
| 02XX | 01XX | 11XX | 57.0 (2) | I | 02XX | 04XX | 02XX | 0.4 (1) | D’ |
| 11XX | 05XX | 16:02 | 59.0 (3) | B, C | 04XX | 03XX | 02XX | 0.13 (1) | V’ |
| 08XX | 06XX | 12XX | 22.0 (4) | B, F | 01XX | 05XX | 04XX | 0.21 (3) | G’ |
| 01XX | 01XX | 01XX | 1.0 (5) | E, F | 07XX | 02XX | 02XX | 0.24 (3) | J’, P’ |
| 07XX | 05XX | 09XX | 37.0 (8) | C, E, I, K | 09XX | 09XX | 04XX | 0.27 (10) | B’, D’, P’, V’ |
| 04XX | 02XX | 02XX | 0.15b (20) | A’, B’, G’, H’ | |||||
| SLA-II, HIGH Antibody Response 2 | SLA-II, LOW Antibody Response 3 | ||||||||
| DRB1 | DQB1 | DQA | Lr-Hp 4 | Haplotype 5 | DRB1 | DQB1 | DQA | Lr-Hp 4 | Haplotype 5 |
| 01XX | 05XX | 04XX | 0.21 (3) | G’ | 07XX | 02XX | 02XX | 0.24 (8) | J’, P’ |
| 10XX | 06XX | 01XX | 0.23 (5) | K’, L’, M’, U’ | 04XX | 02XX | 02XX | O.15b (9) | A’, B’, E’, H’ |
| 09XX | 09XX | 04XX | 0.27 (11) | B’, D’, P’, V’ | |||||
| 04XX | 02XX | 02XX | 0.15b (20) | A’, B’, G’, H’ | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de León, P.; Cañas-Arranz, R.; Saez, Y.; Forner, M.; Defaus, S.; Cuadra, D.; Bustos, M.J.; Torres, E.; Andreu, D.; Blanco, E.; et al. Association of Porcine Swine Leukocyte Antigen (SLA) Haplotypes with B- and T-Cell Immune Response to Foot-and-Mouth Disease Virus (FMDV) Peptides. Vaccines 2020, 8, 513. https://doi.org/10.3390/vaccines8030513
de León P, Cañas-Arranz R, Saez Y, Forner M, Defaus S, Cuadra D, Bustos MJ, Torres E, Andreu D, Blanco E, et al. Association of Porcine Swine Leukocyte Antigen (SLA) Haplotypes with B- and T-Cell Immune Response to Foot-and-Mouth Disease Virus (FMDV) Peptides. Vaccines. 2020; 8(3):513. https://doi.org/10.3390/vaccines8030513
Chicago/Turabian Stylede León, Patricia, Rodrigo Cañas-Arranz, Yago Saez, Mar Forner, Sira Defaus, Dolores Cuadra, María J. Bustos, Elisa Torres, David Andreu, Esther Blanco, and et al. 2020. "Association of Porcine Swine Leukocyte Antigen (SLA) Haplotypes with B- and T-Cell Immune Response to Foot-and-Mouth Disease Virus (FMDV) Peptides" Vaccines 8, no. 3: 513. https://doi.org/10.3390/vaccines8030513
APA Stylede León, P., Cañas-Arranz, R., Saez, Y., Forner, M., Defaus, S., Cuadra, D., Bustos, M. J., Torres, E., Andreu, D., Blanco, E., Sobrino, F., & Hammer, S. E. (2020). Association of Porcine Swine Leukocyte Antigen (SLA) Haplotypes with B- and T-Cell Immune Response to Foot-and-Mouth Disease Virus (FMDV) Peptides. Vaccines, 8(3), 513. https://doi.org/10.3390/vaccines8030513